
DOI:
https://doi.org/10.14483/2256201X.23587Publicado:
01-01-2026Número:
Vol. 29 Núm. 1 (2026): Enero-junioSección:
Artículos de investigación científica y tecnológicaPropagación in vitro de Erytrhina edulis Triana a partir de segmentos nodales
In vitro Propagation of Erytrhina edulis Triana from Nodal Segments
Palabras clave:
ácido indolbutírico, embriones cigóticos, Fabaceae, raíces adventicias (es).Palabras clave:
Indolebutyric acid, zygotic embryos, Fabaceae, adventitious roots (en).Descargas
Referencias
Akbari, F., Arminian, A., Kahrizi, D., & Fazeli, A. (2017). Effect of nitrogen sources on some morphological characteristics of in vitro Stevia rebaudiana Bertoni. Cellular and Molecular Biology, 63(2), 107-111. https://doi.org/10.14715/cmb/2017.63.2.17
Arango, O., Bolaños, V., Ricaurte, D., Caicedo, M., & Guerrero, Y. (2012). Obtención de un extracto proteico a partir de harina de chachafruto (Erythrina edulis). Universidad y Salud, 14(2), 161-167. http://www.scielo.org.co/pdf/reus/v14n2/v14n2a06.pdf
Avendaño, N., & Castillo, A. (2014). El género Erythrina L.(leguminosae-faboideae) en Venezuela. Acta Botánica Venezuelica, 37(2), 123-164. https://www.jstor.org/stable/26611054
Costa, G. M. D., Nepomuceno, C. F., & Santana, J. R. F. D. (2010). Propagação in vitro de Erythrina velutina. Ciência Rural, 40(5), 1090-1096. https://doi.org/10.1590/S0103-84782010005000084
Curi, J. C., Pintado, R. M., & Soriano, F. (2020). Micropropagación de Prosopis pallida (Humb &Bonpl. Ex Willd.) Kunth a partir de yemas apicales. Revista Colombiana de Biotecnología, 22(1), 18-26. https://doi.org/10.15446/rev.colomb.biote.v22n1.70949
de Almeida, M. R., Aumond, M., da Costa, C. T., Schwambach, J., Ruedell, C. M., Correa, L. R., & Fett-Neto, A. G. (2017). Environmental control of adventitious rooting in Eucalyptus and Populus cuttings. Trees, 31, 1377-1390. https://doi.org/10.1007/s00468-017-1550-6
Díaz, L. P., Medina, L. F., Latife, J., Digonzelli, P. A., & Sosa, S. B. (2004). Aclimatación de plantas micropropagadas de caña de azúcar utilizando el humus de lombriz. RIA. Revista de Investigaciones Agropecuarias, 33(2), 115-128. https://www.redalyc.org/pdf/864/86433208.pdf
Díaz-Páez, M., Werden, L. K., Zahawi, R. A., Usuga, J., & Polanía, J. (2021). Vegetative propagation of native tree species: an alternative restoration strategy for the tropical Andes. Restoration Ecology, 30(7), e13611. https://doi.org/10.1111/rec.13611
Fonseca, P. T., Nepomuceno, C. F., Alvim, B. F. M., & de Santana, J. R. F. (2014). Resposta morfogênica de embriões zigóticos de Erythrina velutina Willd. (Leguminosae) cultivados in vitro. Revista Ceres, 61(5), 605-611. https://doi.org/10.1590/0034-737X201461050002
Geisler, G. E., Pinto, T. T., Santos, M., & Paulilo, M. T. S. (2016). Seed structures in water uptake, dormancy release, and germination of two tropical forest Fabaceae species with physically dormant seeds. Brazilian Journal of Botany, 40, 67-77. https://doi.org/10.1007/s40415-016-0334-3
Guariz, H. R., Shimizu, G. D., Paula, J. C., Sperandio, H. V., Ribeiror Junior, W. A., Oliveira, H. C., Jussiani, E. I., Andrello, A. C., Marubayashi, R. Y. P., Picoli, M. H. S., Ruediger, J., Couto, A. P. S., & Moraes, K. A. M. (2022). Anatomy and germination of Erythrina velutina seeds under a different imbibition period in gibberellin. Seeds, 1(3), 210-220. https://doi.org/10.3390/seeds1030018
Intiquilla, A., Jiménez-Aliaga, K., Zavaleta, A. I., Arnao, I., Peña, C., Chavez-Hidalgo, E. L., & Hernández-Ledesma, B. (2016). Erythrina edulis (pajuro) seed protein: A new source of antioxidant peptides. Natural Product Communications, 11(6), 1934578X1601100. https://doi.org/10.1177/1934578X1601100620
Intiquilla, A., Jiménez‐Aliaga, K., Guzmán, F., Álvarez, C. A., Zavaleta, A. I., Izaguirre, V., & Hernández‐Ledesma, B. (2019). Novel antioxidant peptides obtained by alcalase hydrolysis of Erythrina edulis (pajuro) protein. Journal of the Science of Food and Agriculture, 99(5), 2420-2427. https://doi.org/10.1002/jsfa.9449
Javed, S. B., & Anis, M. (2014). Cobalt induced augmentation of in vitro morphogenic potential in Erythrina variegata L.: a multipurpose tree legume. Plant Cell, Tissue and Organ Culture (PCTOC), 120, 463-474. https://doi.org/10.1007/s11240-014-0613-2
Javed, S. B., Alatar, A. A., Anis, M., & Faisal, M. (2017). Synthetic seeds production and germination studies, for short term storage and long distance transport of Erythrina variegata L.: A multipurpose tree legume. Industrial Crops and Products, 105, 41-46. https://doi.org/10.1016/j.indcrop.2017.04.053
Javed, S. B., Alatar, A. A., Anis, M., & El-Sheikh, M. A. (2019). In vitro regeneration of coral tree from three different explants using thidiazuron. HortTechnology, 29(6), 946-951. https://doi.org/10.21273/HORTTECH04398-19
Kirika, M. W., Kahia, J. W., Diby, L. N., Njagi, E. M., Dadjo, C., & Kouame, C. (2015). Micropropagation of an endangered medicinal and indigenous multipurpose tree species: Erythrina abyssinica. HortScience, 50(5), 738-743. https://doi.org/10.21273/HORTSCI.50.5.738
Lozano, E. C., & Zapater, M. A. (2010). El género Erythrina (leguminosae) en Argentina. Darwiniana, Nueva Serie, 48(2), 179-200.
Luangsriumporn, P., Bodhipadma, K., Noichinda, S., Punnakanta, L., & Leung, D. W. (2021). Requirements for efficient plantlet regeneration using cotyledonary nodal explants of purple coral tree (Erythrina fusca Lour.). Vegetos, 34, 37-41. https://doi.org/10.1007/s42535-020-00181-y
Millones, C. E., & Vásquez, E. R. (2008). Micropropagación de plantas de sacha inchi (Plukenetia volubilis L.) provenientes de la provincia de Rodríguez de Mendoza, Región Amazonas. Investigaciones Amazonenses, 2(1), 7-11.
Murashige, T., & Skoog, F. (1962). A revised medium for rapid grown and bioassay with tobacco tissue culture. Physiologia Plantarum, 15(3), 473-497. https://doi.org/10.1111/j.1399-3054.1962.tb08052.x
Palma-Albino, C., Intiquilla, A., Jiménez-Aliaga, K., Rodríguez-Arana, N., Solano, E., Flores, E., Zavaleta, A. I., Izaguirre, V., & Hernández-Ledesma, B. (2021). Albumin from Erythrina edulis (Pajuro) as a promising source of multifunctional peptides. Antioxidantes, 10, 1722. https://doi.org/10.3390/antiox10111722
R Core Team (2022). R: A language and environment for statistical computing. R Foundation for Statistical Computing. https://www.R-project.org/
Russo, R. O. (1991). Erythrina (Leguminosae: Papilionoideae) a versatile genus for agroforestry systems in the tropics. Journal of Sustainable Agriculture, 1(2), 89-109. https://doi.org/10.1300/J064v01n02_06
Rodríguez-Aliaga, N., Jiménez-Aliaga, K., Intiquilla, A., León, J. A., Flores, E., Zavaleta, A. I., Izaquirre, V., Solis-Calero, C. & Hernández-Ledesma, B. (2022). Protection against oxidative stress and metabolic alterations by synthetic peptides derived from Erythrina edulis seed protein. Antioxidantes, 11, 2101. https://doi.org/10.3390/antiox11112101
Schwambach, J., Fadanelli, C., & Fett-Neto, A. G. (2005). Mineral nutrition and adventitious rooting in microcuttings of Eucalyptus globulus. Tree Physiology, 25(4), 487-494. https://doi.org/10.1093/treephys/25.4.487
Sharry, S., Adema, M., Cordal, M. A. B., Villarreal, B., Nikoloff, N., Briones, V., & Abedini, W. (2011). Propagation and conservation of native forest genetic resources of medicinal use by means of in vitro and ex vitro Techniques. Natural Product Communications, 6(7), 985-988. https://doi.org/10.1177/1934578X1100600715
Shasthree, T., Imran, M. A., & Mallaiah, B. (2009). In vitro rooting from callus cultures derived from seedling explants of Erythrina variegata L. Current Trends in Biotechnology and Pharmacy, 3(4), 447-452.
Smýkal, P., Vernoud, V., Blair, M. W., Soukup, A., & Thompson, R. D. (2014). The role of the testa during development and in establishment of dormancy of the legume seed. Frontiers in Plant Science, 5, 351. https://doi.org/10.3389/fpls.2014.00351
Solis, R., Cachique, D., Guerrero-Abad, J. C., Sánchez, M. E. R., & Tapia, L. (2018). In vitro propagation of sacha inchi through organogenesis. Pesquisa Agropecuária Brasileira, 53, 1285-1288. https://doi.org/10.1590/S0100-204X2018001100012
Woodward, A. J., Bennett, I. J., & Pusswonge, S. (2006). The effect of nitrogen source and concentration, medium pH and buffering on in vitro shoot growth and rooting in Eucalyptus marginata. Scientia Horticulturae, 110(2), 208-213. https://doi.org/10.1016/j.scienta.2006.07.005
Yildiz, M., & Er, C. (2002). The effect of sodium hypochlorite solutions on in vitro seedling growth and shoot regeneration of flax (Linum usitatissimum). Naturwissenschaften, 89, 259-261. https://doi.org/10.1007/s00114-002-0310-6
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Recibido: 1 de mayo de 2025; Aceptado: 9 de agosto de 2025
Resumen
Debido a su adaptabilidad y versatilidad, Erythrina edulis Triana ex Micheli es una especie importante para el desarrollo de sistemas agroforestales, para la cual la propagación in vitro proporciona una alternativa para su rápida multiplicación. El objetivo de este estudio fue establecer un protocolo para la propagación in vitro de E. edulis por medio segmentos nodales. A partir de semillas esterilizadas y germinadas in vitro, se obtuvieron segmentos nodales que fueron colocados en medios de enraizamiento, empleando un medio basal MS modificado con la fuente de nitrógeno a la mitad y suplementado con la combinación de ácido naftalenacético (ANA) 1 mg.L-1 más 0.5 mg.L-1 de ácido indolbutírico (AIB) y 1, 3 y 5 mg.L-1 de AIB. El uso de 5 mg.L-1 de AIB registró los mayores promedios en cuanto a número y longitud de raíces, y la aclimatación de las plántulas presentó una tasa de supervivencia de más del 90 %. Este estudio constituye un primer reporte de propagación in vitro de E. edulis.
Palabras clave:
embriones cigóticos, raíces adventicias, ácido indolbutírico, Fabaceae.Abstract
Given its adaptability and versatility, Erythrina edulis Triana ex Micheli is an important species in the development of agroforestry systems, for which in vitro propagation provides an alternative for rapid multiplication. The objective of this study was to establish a protocol for the in vitro propagation of E. edulis using nodal segments. Nodal segments were obtained from sterilized and in vitro germinated seeds, and they were placed in rooting media, using a modified MS basal medium at half nitrogen source, supplemented with a combination of 1 mg.L-1 naphthaleneacetic acid (NAA) plus 0.5 mg.L-1 indolebutyric acid (IBA) and 1, 3, and 5 mg.L-1 IBA. The use of 5 mg.L-1 IBA recorded the highest averages regarding root number and length, and seedling acclimatization exhibited a survival rate of over 90%. This study constitutes the first report for the in vitro propagation of E. edulis.
Keywords:
zygotic embryos, adventitious roots, indolebutyric acid, Fabaceae.INTRODUCCIÓN
El género Erytrhina, de la familia Fabaceae, incluye aproximadamente 120 especies distribuidas en regiones tropicales, destacando a E. edulis Triana ex Micheli, E. velutina Willd., E. fusca Lour., E. variegata Linn., E. abyssinica Lam ex DC. y E. falcata Benth. (Avendaño & Castillo, 2014; Lozano & Zapater, 2010). E. edulis, conocida como chachafruto, pajuro o basul, es una importante fuente de alimento; su semilla es empleada en la alimentación humana y sus ramas herbáceas en la alimentación animal. Además, al ser un especie arbórea, es adecuada para la agroforestería gracias a la capacidad de sus nódulos radiculares para fijar el nitrógeno atmosférico, lo que permite la recuperación de este elemento en el suelo. Por ello, esta especie es ideal para la asociación con pastos o cultivos comerciales (Arango et al., 2012). Su semilla se caracteriza por un alto nivel de proteína, que fluctúa entre 18 y 25 %, destacando por su alto valor nutricional y como potencial fuente de péptidos antioxidantes y multifuncionales, ingredientes importantes en los alimentos funcionales para la prevención y el manejo del estrés oxidativo, la hipertensión y enfermedades crónicas relacionadas con alteraciones metabólicas (Intiquilla et al., 2016; Palma-Albino et al., 2021; Rodríguez-Arana et al., 2022). Adicionalmente, estas semillas son ricas en micronutrientes como calcio, magnesio, hierro, sodio, potasio, fósforo y sulfatos, así como en vitaminas como ácido ascórbico, tiamina, niacina y riboflavina (Intiquilla et al., 2019).
La plantación de árboles de propagación vegetativa en los Andes tropicales es ideal para facilitar la recuperación forestal, pues ayuda a establecer especies leñosas que compiten con los pastos, además de mejorar las condiciones para la reorganización de la comunidad vegetal (Díaz-Páez et al., 2021). Al respecto, la importancia del género Erythrina radica en su aporte al desarrollo de sistemas agroforestales, dados su adaptabilidad y variados usos (como postes vivos de cercas, árboles de sombra para cultivos como el café y el cacao, y fuentes de proteínas, fibra y nutrientes para el ganado, entre otros). A esto se suma el rápido crecimiento de sus plantas, su alta producción de biomasa, su fácil propagación a partir de estacas, su capacidad de soportar podas continuas y su rápida brotación, lo que constituye su atractivo para la agroforestería y los sistemas silvopastoriles (Russo, 1991). Por otro lado, este género tiene la capacidad de fijar nitrógeno atmosférico, una característica de gran importancia al establecer plantaciones en suelos de baja fertilidad y/o en restauración (Russo, 1991).
En los últimos años, para Erythrina se han empleado métodos no convencionales como la propagación in vitro, pues las especies de este género son heterocigotas, y al propagarlas por semilla sexual se obtiene una alta variabilidad genética en las plántulas. Estas limitaciones, inherentes a la propagación convencional, limitan el uso potencial de las especies (Kirika et al., 2015; Javed et al., 2014). Existen reportes de propagación in vitro del género Erythrina que emplean explantes como embriones cigóticos, hipocótilos, segmentos nodales y nudos cotiledonales en E. velutina (Costa et al., 2010; Fonseca et al., 2014); segmentos nodales en E. abyssinica (Kirika et al., 2015); segmentos nodales en E. variegata (Javed et al., 2017); y nudos cotiledonales en E. fusca (Luangsriumporn et al., 2021). El uso de embriones cigóticos en especies de Erytrhina representa una alternativa para asegurar la variabilidad genética (Fonseca et al., 2014). En este contexto, la propagación in vitro es una alternativa para la producción de plántulas y la conservación ex situ de las especies de este género, destacando que las plántulas obtenidas a través de esa técnica poseen características genéticas idénticas a las de las plantas patrón. Además, se encuentran libres de virus, son uniformes y brindan plántulas en menos tiempo y con menos espacio (Costa et al., 2010).
No se tiene conocimiento de estudios sobre la propagación in vitro de E. edulis. Por consiguiente, es necesario desarrollar protocolos que permitan la propagación masiva de esta especie, en aras de reducir la presión sobre los bosques naturales donde se desarrolla. Así las cosas, este trabajo buscó establecer un protocolo para la propagación in vitro de plantas de E. edulis mediante segmentos nodales de plántulas obtenidas de la germinación de embriones cigóticos.
MATERIALES Y MÉTODOS
El material biológico utilizado comprendía semillas de pajuro (ecotipo rojo) de E. edulis, provenientes del distrito de Luya, Amazonas, Perú (altitud: 2342 m de altitud, coordenadas: 06º11’54” S, 83º58’32.27” W). A partir de las semillas se cultivaron los embriones, y las plántulas desarrolladas sirvieron como donadoras de segmentos nodales para la fase de enraizamiento.
El medio de cultivo para la germinación de los embriones cigóticos fue preparado empleando sales basales y vitaminas MS (Murashige & Skoog, 1962), suplementando con 100 mg.L-1 de mio-inositol, 25 g.L-1 de sacarosa, 0.1 mg.L-1 de ácido giberélico (AG3), 0.04 mg.L-1 de 6-furfurilaminopurina (KIN) y 1.5 g.L-1 de phytagel. El pH fue ajustado a 5.8. El medio de crecimiento y desarrollo de las plántulas se preparó mediante el medio basal Woody Plant Medium (WPM), con vitaminas MS, 100 mg.L-1 de mio-inositol, 30 g.L-1 de sacarosa, 1.5 g.L-1 de phytagel y 3 g.L-1 de carbón activado. El pH fue ajustado a 5.8. Los segmentos nodales se enraizaron en medio MS más vitaminas, modificado en la fuente de nitrógeno al 50 % (KNO3 y NH4NO3), suplementando con mio-inositol 100 mg.L-1, sacarosa 30 g.L-1 y diferentes concentraciones de reguladores de crecimiento, i.e., ANA: 1 mg.L-1 + AIB: 0.5 mg.L-1, AIB: 1, 3 y 5 mg.L-1. El medio fue gelificado con phytagel 1.5 g.L-1 y carbón activado 3 g.L-1, y el pH fue ajustado a 5.8.
El establecimiento in vitro de los embriones cigóticos de pajuro inició con el remojo de las semillas por 30 minutos, utilizando detergente y una solución de lejía comercial al 5 % (Clorox, 3.5 v/v de hipoclorito de sodio). Después, las semillas fueron cepilladas para retirar los contaminantes adheridos al tegumento, y luego fueron enjuagadas con agua corriente. Posteriormente, a las semillas se les retiró la cubierta próxima a la radícula del embrión, para luego llevarlas a una cámara de flujo laminar e iniciar el proceso de desinfección. Se agregó etanol al 70 % por un minuto, se enjuagó con agua destilada estéril y se adicionó solución de lejía al 5, 10 y 15 % por 15 y 30 minutos. Luego se realizaron tres enjuagues con agua destilada estéril. Desinfectadas las semillas, empleando pinza y escalpelo, se realizaron cortes de aproximadamente 1 cm por lado en el endospermo próximo al embrión cigótico. Las estructuras cúbicas resultantes fueron colocadas en tubos de ensayo que contenían el medio de germinación. Los embriones cigóticos germinaron a los 15 días de cultivo. La Figura 1 ilustra los procedimientos mencionados. Posteriormente, a los 28 días, los embriones fueron traspasados a medios de crecimiento y desarrollo. Las condiciones para la germinación de los embriones y el desarrollo de las plántulas de pajuro incluyeron una temperatura de 24-26 ºC, una irradiancia de 60 µmol.m-2s-1 y un fotoperiodo de 16/8 horas (luz/oscuridad), empleando lámparas fluorescentes de 36 W con luz fría.

Figura 1: Establecimiento in vitro de los embriones cigóticos de E. edulis (pajuro): a) semilla lavada y desprovista del tegumento en la zona de la radícula del embrión cigótico, b) remojo y desinfección de las semillas, c) embrión cigótico con parte de cotiledones en medio de germinación, d) inicio de la germinación del embrión cigótico a los 15 días de cultivo in vitro, e) crecimiento y desarrollo del embrión cigótico in vitro, f) plántula a las 12 semanas de cultivo en medio de crecimiento y desarrollo. Barra blanca = 1 cm.
Se obtuvieron entre cuatro y cinco segmentos nodales de cada plántula germinada, de aproximadamente 12 semanas, los cuales fueron colocados en medios de enraizamiento. Luego de cinco semanas de cultivo in vitro, se evaluaron el porcentaje de enraizamiento y el número y la longitud de las raíces (Figura 2).

Figura 2: Enraizamiento de segmentos nodales de E. edulis (pajuro): a) plántula donadora de segmentos nodales, b) segmentos nodales con una a dos yemas axilares, c) colocación de los segmentos nodales en medios de enraizamiento, d) segmentos nodales enraizados a la quinta semana de cultivo in vitro. Barra blanca = 1 cm.
Las plántulas enraizadas de pajuro fueron aclimatadas en macetas cuadradas de 5 x 5 x 7.5 cm, empleando el sustrato comercial Premix #3. Posteriormente, fueron transferidas a bolsas negras que contenían un sustrato de suelo franco-arenoso y humus de lombriz en una proporción de 2:1, y se mantuvieron en casa malla (Figura 4).
El diseño experimental empleado fue completamente al azar, con cuatro tratamientos y tres repeticiones por tratamiento. Cada repetición consistió en un vaso de magenta con tres segmentos nodales. La longitud de las raíces se midió mediante observación directa y estimación visual, utilizando una regla graduada. Se realizó un análisis de varianza para la longitud y el número de raíces. Las medias se compararon mediante la prueba de Tukey, con un nivel de significancia de p ≤ 0.05. Los datos de longitud y número de raíces fueron transformados mediante (x + 0.5)1/2 antes de su procesamiento. Se utilizó el programa R versión 4.3.2 para el procesamiento de los datos (R Core Team, 2022).
RESULTADOS
La tasa de contaminación durante el establecimiento in vitro de los embriones cigóticos fue de 0 %, demostrando que este tejido no es afectado por contaminantes cuando se emplea lejía al 10 % por 30 minutos; el protocolo de desinfección de tejidos de la semilla fue eficaz. Asimismo, la tasa de supervivencia de los embriones superó el 90 %, y su germinación fue del 86 % (Figuras 1d y 1e). Con lejía al 5 % por 30 minutos y al 10 % por 15 minutos, las semillas presentaron contaminación fúngica, mientras que su aplicación al 15 % por 15 y 30 minutos contrarrestó la contaminación, pero los embriones perdieron viabilidad.
El crecimiento y desarrollo de las plántulas in vitro en el medio de cultivo fue adecuado, logrando especímenes vigorosos y con buen desarrollo del tallo (Figura 1f). Esto permitió que, a las 12 semanas de cultivo in vitro, se obtuvieran segmentos nodales para su uso en la fase de enraizamiento (Figura 2b).
El enraizamiento de los segmentos nodales ocurrió entre la cuarta y la quinta semana de cultivo (Figura 2d). El mayor número de raíces (1.8) de 3.2 cm se observó en el 91.7 % de los segmentos nodales al emplear 5 mg.L-1 de AIB, mientras que el uso de 1 y 3 mg.L-1 de AIB y la combinación de 2 mg.L-1 de ANA más 0.5 mg.L-1 de AIB mostraron una respuesta inferior (Figura 3a y 3b). Las raíces adventicias inducidas se caracterizaron por ser gruesas y se originaron en la parte basal del segmento nodal (Figuras 2c y 4a ).
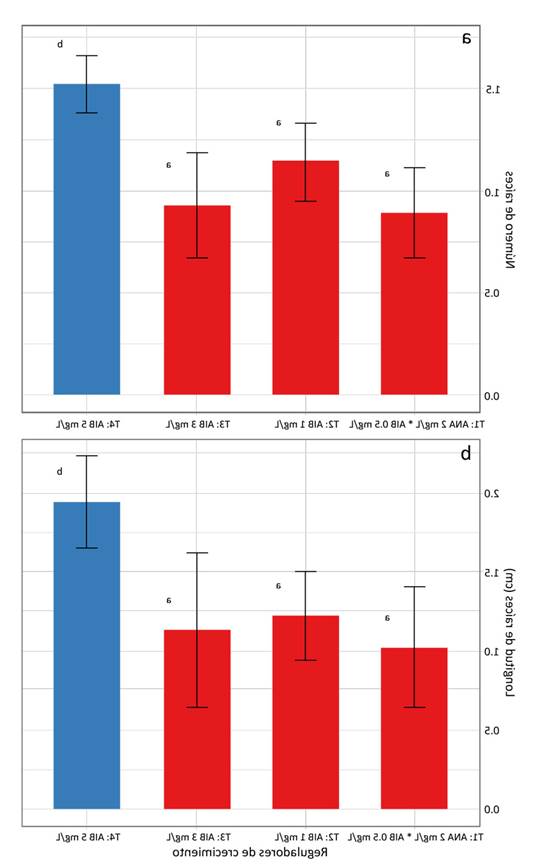
Figura 3: Efecto de los reguladores de crecimiento ANA y AIB en el enraizamiento de segmentos nodales de E. edulis (pajuro): a) número de raíces, b) longitud de raíces. Los datos se presentan con la media ± la desviación estándar. Diferentes letras indican diferencias significativas en los parámetros evaluados para p ≤ 0.05 en la prueba de Tukey.
En la etapa de aclimatación, la tasa de supervivencia superó el 90 %, indicando claramente que la composición del sustrato Premix brindó condiciones óptimas para que las plántulas tuvieran éxito durante el proceso, obteniendo plantas vigorosas y de buen aspecto fitosanitario (Figura 3c).
DISCUSIÓN
El establecimiento de explantes in vitro es un paso importante en el cultivo de tejidos, siendo importante evaluar la eficacia de los métodos de esterilización empleados. El uso de lejía comercial (hipoclorito de sodio) ayuda en la esterilización de los explantes (Yildiz & Er, 2002). Para el género Erythrina se ha empleado cloruro de mercurio o hipoclorito de sodio como solución esterilizante. El uso de cloruro de mercurio al 0.1 %, por periodos de 2 a 3 minutos, se ha reportado en la esterilización de semillas, segmentos nodales, hojas, tallos, cotiledones y raíces de E. variegata (Shasthree et al., 2009; Javed & Anis, 2014; Javed et al., 2019). Por su parte, el hipoclorito de sodio al 10 %, aplicado por 25 minutos, se ha empleado en la esterilización de semillas de E. abyssinica (Kirika et al., 2015), y por 15 minutos en la esterilización de semillas de E. velutina (Costa et al., 2010). En esta investigación no se registró contaminación a los siete días de cultivo tras la esterilización superficial de las semillas mediante la solución de lejía comercial al 10 % (Clorox, 3.5 v/v de hipoclorito de sodio), que se aplicó por 30 minutos. A los 15 días de cultivo in vitro, se observó la emisión de la radícula. Es preciso añadir que, si se utilizaba una menor concentración de lejía comercial (5 %) o un menor tiempo de exposición (15 minutos), las semillas se contaminaban, y, si la concentración excedía el 10 %, el embrión cigótico perdía viabilidad.
Las especies Peltophorum dubium (Spreng.) y Mimosa bimucronata (DC) O. Kunze, pertenecientes a la familia Fabaceae de bosques tropicales, se caracterizan por semillas con dormancia física. Este rasgo está asociado al tegumento, que posee macroesclereidas impregnadas con sustancias químicas hidrofóbicas, lo que les confiere impermeabilidad al agua (Geisler et al., 2016). El género Erythrina también contiene especies cuyas semillas poseen testa, con una anatomía similar a la de otras semillas de Fabaceae, compuesta por macroesclereidas compactas, osteoesclereídas y varias capas de parénquima esclerificado, impregnado por compuestos fenólicos y suberina, como se ha reportado para E. velutina (Smýkal et al., 2014; Guariz et al., 2022). En esta investigación, durante el establecimiento de los embriones cigóticos de E. edulis, se retiró el tegumento próximo a la radícula del embrión, permitiendo romper la dormancia física y facilitando el contacto directo entre esta área y el desinfectante, lo que resultó en porcentajes de germinación por encima del 90 %. Este es un hallazgo importante, pues la concentración de lejía comercial (10 %) y el tiempo de exposición (30 minutos) utilizados tuvieron un efecto positivo al restringir la contaminación por debajo de la cubierta seminal sin afectar los tejidos del embrión cigótico, lo que redundó en altas tasas de germinación.
La germinación de los embriones cigóticos de E. edulis en el medio de cultivo MS suplementado con bajas concentraciones de reguladores crecimiento (0.1 mg.L-1 de AG3 y 0.04 mg.L-1 KIN) fue adecuada. Al respecto, al evaluar 200 mg.L-1 de AG3 con diferentes tiempos de exposición, Guariz et al. (2022) concluyeron que las semillas de E. velutina no requieren del uso de giberelinas exógenas, solo de un proceso de escarificación. En este estudio, el retiro de la cubierta seminal para el proceso de desinfección de las semillas y el uso de segmentos cuadrados resultaron adecuados para el establecimiento in vitro los embriones cigóticos de E. edulis.
El crecimiento y desarrollo de las plántulas de E. edulis a partir de los embriones cigóticos fue favorable cuando se empleó el medio basal WPM. Por el contrario, cuando se empleó el medio MS, el crecimiento y desarrollo de las plántulas fue afectado por la presencia de clorosis en las hojas superiores y el necrosamiento de las hojas basales, con su posterior caída. Esto podría deberse a que el MS contiene una mayor cantidad de nutrientes, particularmente de nitrógeno, lo que puede haber producido desórdenes fisiológicos en las plántulas (Curi et al., 2020). El medio basal WPM ha sido empleado en especies del género Erythrina porque muestra una respuesta positiva en lo que respecta al crecimiento y desarrollo de las plántulas, como es el caso de E. velutina (Costa et al., 2010; Fonseca et al., 2015) y E. crista-galli (Sharry et al., 2011). Este medio posee una menor concentración de macroelementos (N, P, K, S), por lo que ofrece buenos resultados en especies leñosas. Por el contrario, en E. variegata, E. velutina, E. abyssinica y E. fusca, las plántulas se desarrollaron adecuadamente en medios MS, probablemente porque estos contienen cuatro veces más nitrógeno que el medio basal WPM, ya que algunas especies de la familia Fabaceae requieren altos niveles de este elemento para desarrollarse adecuadamente (Javed et al., 2014; Kirika et al., 2015; Javed et al., 2019; Luangsriumporn et al., 2021).
En estudios sobre la propagación in vitro del género Erythrina se han empleado segmentos nodales y reguladores de crecimiento, e.g., 1 mg.L-1 de BAP y 0.1 mg.L-1 de ANA en E. variegata, obteniendo una media de 5.9 brotes por explante después de ocho semanas de cultivo in vitro (Javed & Anis, 2014), así como un valor de 7.7 tras cuatro semanas con 0.3 mg.L-1 de TDZ (Javed et al., 2019); 4 y 4.5 mg.L-1 de BAP en E. velutina, obteniendo 2.4-2.7 brotes por explante a los 30 días de cultivo (Costa et al., 2010; Fonseca et al., 2015); y 8.6 mg.L-1 de BAP en E. abyssinica, con 6.8 brotes por explante (Kirika et al., 2015). En este trabajo, las plántulas de E. edulis se propagaron únicamente mediante segmentos nodales extraídos de plántulas a las 12 semanas de cultivo in vitro en medios de cultivo, crecimiento y desarrollo. Todo ello, sin utilizar reguladores de crecimiento, lo cual es importante porque estos segmentos no formaron callos friables.
En cuanto a la inducción de raíces adventicias en los segmentos nodales, se reporta el empleo del medio basal MS con fuente de nitrógeno al 50 % (KNO3 y NH4NO3). Esta modificación ha sido empleada en especies como Plukenetia volubilis (Millones & Vásquez, 2008; Solis et al., 2018), Eucalyptus globulus (Schwambach et al., 2005) y Stevia rebaudiana (Akbari et al., 2017). La disminución de la fuente de nitrógeno tiene un efecto significativo en el número y la longitud de las raíces, y su incremento causa un descenso en la longitud radicular (Schwambach et al., 2005). Al respecto, Woodward et al. (2006) y de Almeida et al. (2017) han afirmado que el amonio ocasiona una disminución del pH en el medio de cultivo, y que los nitratos aumentan este valor, cuyos desbalances pueden afectar el crecimiento y desarrollo de las raíces. En esta investigación, la disminución de la fuente de nitrógeno en el medio basal MS tuvo un efecto positivo, probablemente debido a la mayor estabilidad del pH del medio de enraizamiento, lo que favorece el crecimiento y desarrollo de las raíces.
En el caso de Erythrina, se han empleado diferentes concentraciones de AIB para la inducción, crecimiento y desarrollo de las raíces adventicias. En E. variegata, el uso de 0.5 mg.L-1 de AIB registró el mayor número (3.3 por segmento nodal) y la mayor longitud (3.2 cm) de las raíces a las cuatro semanas de cultivo (Javed & Anis, 2014). En E. abyssinica, 0.5 mg.L-1 de AIB produjeron la mayor longitud radicular (2.4 por segmento nodal) a las cuatro semanas de cultivo (Kirika et al., 2015). En este trabajo, por su parte, se registraron resultados similares para el enraizamiento de los segmentos nodales de E. edulis a partir de la quinta semana de cultivo in vitro. El uso de 5 mg.L-1 de AIB tuvo un efecto positivo en el número de las raíces (1.8 por segmento nodal) y su longitud (3.2 cm), siendo la prueba significativa (Figuras 4a y 4b). Por otro lado, cuando se emplearon concentraciones de AIB por encima de los 5 mg.L-1, se inhibió la inducción de raíces adventicias. Asimismo, estos parámetros disminuyeron con concentraciones menores.

Figura 4.: Aclimatación de plántulas de E. edulis (pajuro) propagadas in vitro: a) plántulas enraizadas in vitro, b) aclimatación de plántulas, c) crecimiento y desarrollo de las plántulas en casa malla
En la quinta semana de aclimatación se registró una tasa de supervivencia del 90 %, por lo que las plantas fueron colocadas en bolsas negras para su crecimiento y desarrollo en casa malla. En el sustrato, conformado por suelo franco-arenoso y humus de lombriz en una proporción de 2:1, las plantas de E. edulis alcanzaron un buen crecimiento y desarrollo, presentando plantones con buena vigorosidad y aspecto fitosanitario. Ambos componentes del sustrato permitieron mejorar la retención de humedad y la aireación, condiciones indispensables para el establecimiento del sistema radicular de las plántulas y para el adecuado desarrollo de las plantas (Díaz et al., 2004).
CONCLUSIONES
Este estudio demostró que se puede realizar la propagación in vitro de E. edulis a partir de segmentos nodales, obteniendo plántulas uniformes sin callos. Este es el primer reporte de este tipo para la especie. Los hallazgos de este trabajo aportan una base para los programas de propagación masiva y conservación ex situ de esta especie y otras del género Erythrina.
Acknowledgements
AGRADECIMIENTOS
Los autores agradecen al Vicerrectorado de Investigación de la Universidad Nacional Toribio Rodríguez de Mendoza del Amazonas por asignar recursos para publicación de esta investigación.
REFERENCIAS




Licencia
Derechos de autor 2025 Colombia forestal

Esta obra está bajo una licencia internacional Creative Commons Atribución-CompartirIgual 4.0.
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






