
DOI:
https://doi.org/10.14483/2256201X.23871Publicado:
01-01-2026Número:
Vol. 29 Núm. 1 (2026): Enero-junioSección:
Artículos de investigación científica y tecnológicaAlmacenamiento y acumulación de carbono en clones de caucho en la Orinoquía colombiana
Carbon Storage and Accumulation in Rubber Clones in the Colombian Orinoquía
Palabras clave:
Biomass, genotypes, Hevea brasiliensis, mathematical models, plantation (en).Palabras clave:
biomasa, genotipo, Hevea brasiliensis, modelo matemático, plantación (es).Descargas
Referencias
Andrade, H., Orjuela, J., & Joven, C. (2022). Modelos de biomasa aérea y subterránea de Hevea brasiliensis y Theobroma grandiflorum en la Amazonía colombiana. Colombia forestal, 25(2), 57-69. https://doi.org/10.14483/2256201X.18464
Archontoulis, S., & Miguez, F. (2015). Nonlinear Regression Models and Applications in Agricultural Research. Agronomy Journal, 107(2), 786-798. https://doi.org/10.2134/agronj2012.0506
Blagodatsky, S., Xu, J., & Cadisch, G. (2016). Carbon balance of rubber (Hevea brasiliensis) plantations: A review of uncertainties at plot, landscape and production level. Agriculture, Ecosystems & Environment, 221, 8-19. https://doi.org/10.1016/j.agee.2016.01.025
Brahma, B., Sileshi, G., Nath, A., & Das, A. (2017). Development and evaluation of robust tree biomass equations for rubber tree (Hevea brasiliensis) plantations in India. Forest Ecosystems, 4(1), 14. https://doi.org/10.1186/s40663-017-0101-3
Castañeda-Garzón, S., Mora, A., Tarazona, M., & Hernández, D. (2025). Growth models of Hevea brasiliensis genotypes in clonal fields of the Colombian Orinoquia. Revista Facultad Nacional de Agronomía Medellín, 78(2), 11057-11068. https://doi.org/10.15446/rfnam.v78n2.113865
Castañeda-Garzón, S., Rivano, F., & Mora, A. (2024). Crecimiento de clones de Hevea brasiliensis (Willd. Ex A.Juss.) Müll.Arg. En etapa juvenil establecidos en campos clonales, Meta, Colombia. Temas Agrarios, 29(1), 53-65. https://doi.org/10.21897/5099hh92
Diniz, A., Pereira, M., Balieiro, F., Silva, E, Santos, F, Lisboa, F, Oliveira, A., & Cruz, R. (2015). Contrasts in areas of rubber tree clones in regard to soil and biomass carbon stocks. Revista Brasileira de Ciência Do Solo, 39(5), 1378-1385. https://doi.org/10.1590/01000683rbcs20140274
Durán, E., Duque, L., & Suárez, J. (2011). Estimación de carbono en sistemas agroforestales de Hevea brasiliensis en la Amazonia Colombiana. Ingenierías & Amazonia, 4(1), 19-28.
Fonseca, W., Ávila, C., Murillo, R., & Rojas, M. (2021). Predicción de biomasa y carbono en plantaciones clonales de Tectona grandis L.f. Colombia forestal, 24(1), 31-44. https://doi.org/10.14483/2256201X.15961
Fu, Y., Tan, H., Kou, W., Xu, W., Wang, H., & Lu, N. (2024). Estimation of rubber plantation biomass based on variable optimization from Sentinel-2 remote sensing imagery. Forests, 15(6), 900. https://doi.org/10.3390/f15060900
Hytönen, J., Nurmi, J., Kaakkurivaara, N., & Kaakkurivaara, T. (2019). Rubber tree (Hevea brasiliensis) biomass, nutrient content, and heating values in Southern Thailand. Forests, 10(8), 638. https://doi.org/10.3390/f10080638
Instituto Amazónico de Investigaciones Científicas -SINCHI-. (2019). Valoración y análisis de la biodiversidad y servicios ecosistémicos asociados a campos clonales de caucho en Caquetá, Amazonia colombiana (A. Sterling Cuellar & C. H. Rodríguez León, Eds.). Instituto Amazónico de Investigaciones Científicas - SINCHI. https://sinchi.org.co/files/publicaciones/novedades%20editoriales/pdf/Caucho_web.pdf
Instituto de Hidrología, Meteorología y Estudios Ambientales - IDEAM, Fundación Natura, Programa de las Naciones Unidas para el Desarrollo, Ministerio de Ambiente y Desarrollo Sostenible, Departamento Nacional de Planeación, & Cancillería de Colombia. (2022). Inventario departamental de gases efecto invernadero y carbono negro-2018. Tercer Informe Bienal de Actualización de Cambio Climático de Colombia. Instituto de Hidrología, Meteorología y Estudios Ambientales - IDEAM. http://documentacion.ideam.gov.co/openbiblio/bvirtual/023936/InventarioDGEICarbonoNegro.pdf
IPCC. (2006). 2006 IPCC Guidelines for National Greenhouse Gas Inventories. Volume 4 Agriculture, forestry and other land use (National Greenhouse Gas Inventories Programme, S. Eggleston, L. Buendia, K. Miwa, T. Ngara, & K. Tanabe, Eds.). https://www.ipcc-nggip.iges.or.jp/public/2006gl/pdf/4_Volume4/V4_00_Cover.pdf
Kabinesh, V., Suwethaasri, D., Baranidharan, K., Ravi, R., Tilak, M., Kalpana, M., Ragunath, K. P., Vennila, S., Hemalatha, P., Vijayabhama, M., Bargavi, S., & Eniya, A. (2025). A critical review of exploring the recent trends and technological advancements in forest biomass estimation. Plant Science Today, 12(sp1), 1-12. https://doi.org/10.14719/pst.6695
Lenth, R. (2016). Least-Squares Means: The R Package lsmeans. Journal of Statistical Software, 69, 1-33. https://doi.org/10.18637/jss.v069.i01
López-Reyes, L., Domínguez-Domínguez, M., Martínez-Zurimendi, P., Zavala-Cruz, J., Gómez-Guerrero, A., & Posada-Cruz, S. (2016). Carbono almacenado en la biomasa aérea de plantaciones de hule (Hevea brasiliensis Müell. Arg.) de diferentes edades. Madera y Bosques, 22(3), 49-60. https://doi.org/10.21829/myb.2016.2231456
Maggiotto, S., Oliveira, D., Marur, C., Stivari, S., Leclerc, M., & Wagner-Riddle, C. (2014). Potential carbon sequestration in rubber tree plantations in the northwestern region of the Paraná State, Brazil. Acta Scientiarum. Agronomy, 36, 239-245. https://doi.org/10.4025/actasciagron.v36i2.17404
Mesike, C., & Idoko, S. (2024). An overview of carbon sequestration potential of rubber tree plantations. Climate Change, 10(1), 1-5. https://doi.org/10.54905/disssi.v10i28.e10cc1043
Monroy, C., & Návar, J. (2004). Ecuaciones de aditividad para estimar componentes de biomasa de Hevea brasiliensis Muell. Arg., en Veracruz, México. Madera y Bosques, 10(2), 29-43. https://doi.org/10.21829/myb.2004.1021273
Moreno, J., Burgos, J., Nieves, H., & Buitrago, C. (2005). Modelo alométrico general para la estimación del secuestro de carbono por plantaciones de caucho Hevea brasilensis mull arg. En Colombia. Colombia forestal, 9(18), 5-21. https://doi.org/10.14483/udistrital.jour.colomb.for.2005.1.a01
NASA. (2025). NASA POWER | Prediction Of Worldwide Energy Resources. https://power.larc.nasa.gov/
Orjuela-Chaves, J. A., & Vargas-Valenzuela, Y. (2014). Potential of carbon storage of rubber (Hevea brasiliensis Müll. Arg.) plantations in monoculture and agroforestry systems in the Colombian Amazon. Tropical and Subtropical Agroecosystems, 17, 231-240.
Pardo-Rozo, Y., Andrade-Castañeda, H., Muñoz-Ramos, J., & Velásquez-Restrepo, J. (2021). Carbon capture in three land use systems in the Colombian Amazonia. Revista de Ciencias Agrícolas, 38(2), 111-123. https://doi.org/10.22267/rcia.213802.160
Pinheiro, J., & Bates, D. (2000). MixedEffects Models in S and S-PLUS. SpringerVerlag. https://link.springer.com/book/10.1007/b98882
R Core Team. (2025). R: A Language and Environment for Statistical Computing. R: The R Project for Statistical Computing. https://www.r-project.org/
Rolim, S, & Piotto, D. (2024). Diameter growth models and performance of 100 tropical tree species in silvicultural trials in Brazil. Forest Ecology and Management, 569, 122202. https://doi.org/10.1016/j.foreco.2024.122202
Sileshi, G. (2014). A critical review of forest biomass estimation models, common mistakes and corrective measures. Forest Ecology and Management, 329, 237-254. https://doi.org/10.1016/j.foreco.2014.06.026
Thomas, S., & Martin, A. (2012). Carbon content of tree tissues: A synthesis. Forests, 3(2), 332-352. https://doi.org/10.3390/f3020332
Wauters, J., Coudert, S., Grallien, E., Jonard, M., & Ponette, Q. (2008). Carbon stock in rubber tree plantations in Western Ghana and Mato Grosso (Brazil). Forest Ecology and Management, 255(7), 2347-2361. https://doi.org/10.1016/j.foreco.2007.12.038
Zianis, D., & Mencuccini, M. (2004). On simplifying allometric analyses of forest biomass. Forest Ecology and Management, 187(2), 311-332. https://doi.org/10.1016/j.foreco.2003.07.007
Zuur, A., Hilbe, J., & Ieno, E. (2013). A beginner’s guide to GLM and GLMM with R: A frequentist and bayesian perspective for ecologists. Highland Statistics Limited. https://www.highstat.com/index.php/books2?view=article&id=21&catid=18
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Recibido: 5 de julio de 2025; Aceptado: 3 de octubre de 2025
Resumen
La estimación de biomasa es fundamental para conocer las reservas de carbono. En el paisaje de piedemonte y altillanura del departamento del Meta, Colombia, se calculó la biomasa aérea de nueve clones de Hevea brasiliensis por método directo en árboles de 11 años. Posteriormente, se ajustaron ecuaciones alométricas y se generó un modelo de biomasa a partir de la circunferencia del tronco. Se evaluaron cuatro modelos de crecimiento no lineal y, con el mejor de ellos, se calcularon los parámetros y las métricas de crecimiento de genotipos en los campos clonales La Libertad y Taluma. Los clones FDR 5788 y PMB 1 reportaron una mayor captura de carbono en La Libertad y Taluma. En la región de la Orinoquía, estos resultados son útiles para estimar con mayor precisión la captura de carbono y aportan información para la formulación de estrategias de mitigación del cambio climático y posibles esquemas de pago por servicios ambientales.
Palabras clave:
biomasa, genotipo, Hevea brasiliensis, modelo matemático, plantación.Abstract
Biomass estimations are essential for determining carbon stocks. In the piedmont and high plains of the department of Meta, Colombia, the aboveground biomass of nine Hevea brasiliensis clones was calculated using the direct method in 11-year-old trees. Subsequently, allometric equations were fitted, and a biomass model was generated based on trunk circumference. Four nonlinear growth models were evaluated, and, with the best-performing one, the growth parameters and metrics of the genotypes were calculated in the clonal fields La Libertad and Taluma. The clones FDR 5788 and PMB 1 showed higher carbon capture in both La Libertad and Taluma. In the Orinoquía region, these results are useful for improving the accuracy of carbon capture estimates and providing valuable information to aid in the formulation of climate change mitigation strategies and potential payment schemes for environmental services.
Keywords:
biomass, genotypes, Hevea brasiliensis, mathematical models, plantation.INTRODUCCIÓN
Según el inventario de gases efecto invernadero (GEI) en Colombia, el departamento del Meta ocupó el pri- mer lugar en emisiones (32 693.5 kt CO2eq, 11 % del total nacional) y el tercero en absorción de GEI (-2772.8 kt CO2eq, 12 % del total nacional) (IDEAM et al., 2022). Hasta 2018, este departamento presentaba las ma- yores absorciones por cultivos permanentes del país (16 % del total), asociadas principalmente al cultivo de palma de aceite y caucho, y correspondían a -1073.6 kt CO2eq (39 % del departamento) (IDEAM et al., 2022). Entre las medidas de mitigación de GEI está la gestión de bosques y el incremento de las reservas de carbono, para lo cual es indispensable determinar la captura de carbono.
En los ecosistemas terrestres, se han identificado cinco depósitos claves de carbono: biomasa aérea y subte- rránea, hojarasca, necromasa y carbono orgánico del suelo (IPCC, 2006). Entre ellos, la biomasa aérea es el depósito más visible, dominante, dinámico y significativo, representando aproximadamente el 30 % del total de carbono almacenado (Kabinesh et al., 2025). En el ámbito forestal, la estimación precisa de la biomasa ha ganado relevancia en las últimas décadas, impulsada por la conciencia sobre el cambio climático y el papel crucial que esta desempeña tanto en el secuestro de carbono como en la liberación de GEI como resultado de la deforestación (Kabinesh et al., 2025). Estas estimaciones son útiles para inventarios de carbono y las nego- ciaciones internacionales sobre su comercio.
Hevea brasiliensis (Willd. ex A.Juss.) Müll.Arg. es la mayor fuente de caucho natural, desempeña funciones vitales en diferentes industrias y contribuye a la captura de carbono y la reducción de GEI (Fu et al., 2024; Mesike & Idoko, 2024). Las principales reservas de carbono en caucho se encuentran en la biomasa aérea y subterránea, en el carbono orgánico del suelo y en otros componentes menores como la hojarasca, la madera muerta y el látex (Blagodatsky et al., 2016; Mesike & Idoko, 2024).
En el caso de H. brasiliensis, se dispone de ecuaciones alométricas de biomasa para India, China, Tailandia y México (Brahma et al., 2017; Fu et al., 2024; Hytönen et al., 2019; López-Reyes et al., 2016; Monroy & Návar, 2004), así como de reportes de stock de carbono en Brasil, México y Ghana (Diniz et al., 2015; Maggiotto et al., 2014; Wauters et al., 2008). Los avances hechos en Colombia incluyen (i) la generación de ecuaciones de biomasa aérea y subterránea para Caquetá, Guaviare, Meta, Santander y Tolima a partir del muestreo de árboles de entre 1 y 36 años, provenientes de los clones FX 3864, FX 25, FX 2261, IAN 713, IAN 873 y PB 86 (Moreno et al., 2005); (ii) la generación de modelos para la Amazonía a partir de árboles de entre 1 y 50 años muestreados en Caquetá sin distinción de clon (Andrade et al., 2022); y (iii) la estimación de la captura de carbono en sistemas agroforestales (SAF) y plantaciones en Caquetá (Durán et al., 2011; Orjuela-Chaves & Vargas-Valenzuela, 2014; SINCHI, 2019; Pardo-Rozo et al., 2021). Sin embargo, en la Orinoquía predominan plantaciones monoclonales (RRIM 600 y FX 3864) bajo distintas condiciones edafoclimáticas, se desconoce su desempeño a nivel genotípico y no hay modelos locales para la estimación de estas variables y su proyección a nivel de área. Por lo tanto, la cuantificación del servicio de captura de carbono podría convertirse para los productores en una posibilidad de acceso a otros mercados (Moreno et al., 2005).
La Corporación Colombiana de Investigación Agropecuaria (Agrosavia) dispone de campos experimentales para evaluar clones de H. brasiliensis de la colección CMS (CIRAD-Michelin Selection), los cuales fueron en- tregados por medio de acuerdos de transferencia. Es por ello que, ante el desconocimiento del desempeño a nivel de genotipo, y en línea con las demandas del sector forestal, el objetivo de este estudio fue determinar el almacenamiento y la acumulación de carbono, en aras de contribuir a la evaluación de clones de caucho en la Orinoquia (Castañeda-Garzón et al., 2024, 2025).
MATERIALES Y MÉTODOS
Área de estudio
En el departamento del Meta (Colombia), en el paisaje de piedemonte llanero, se encuentra el campo clonal a pequeña escala (CCPE) La Libertad, en el Centro de Investigación La Libertad de Agrosavia (Villavicencio, 4°3'37.43" latitud norte, 73°27'52.47" longitud oeste, 335 m de altitud). En esta plantación, en octubre de 2013, se establecieron nueve clones de H. brasiliensis (CDC 312, CDC 56, FDR 4575, FDR 5597, FDR 5788, FX 3864, MDF 180, PMB 1 y RRIM 600), sembrados con un espaciamiento de 6 x 3 m. Adicionalmente, en el pai- saje de altillanura se encuentra el campo clonal a gran escala (CCGE) Taluma, en la Finca Experimental Taluma de Agrosavia (Puerto López, 4°22'39.2" latitud norte, 72°13'49.9" longitud oeste, 176 m de altitud), el cual fue establecido en agosto de 2013, con un espaciamiento de 6 x 3 m y conformado por cinco clones (CDC 312, FDR 5788, FX 3864, PMB 1 y RRIM 600) (Castañeda-Garzón et al., 2024). Estos campos clonales se encuentran en relieve plano y suelos oxisoles de baja fertilidad, moderadamente profundos y fuertemente ácidos, con alto contenido de aluminio, buen drenaje y baja saturación, y se diferencian por sus texturas franco arenosa (La Libertad) y franco-arcillo arenosa (Taluma) (Castañeda-Garzón et al., 2024).
En los dos campos clonales, las prácticas de manejo realizadas fueron la siembra, el control químico y mecáni- co de arvenses sin poda y el control fitosanitario. Allí, el plan de fertilización implementado hasta el tercer año incluyó urea (54 %), KCL (27 %) y fosfato diamónico (19 %) en dosis de 370 g.planta-1. En el CCPE La Libertad, en el cuatro y séptimo año, se aplicó urea (22 %), KCL (44 %), Amidas (22 %), MAP (8 %) y Borozinco (3 %) en dosis de 800g/planta. Al quinto año, en el CCGE Taluma se aplicó urea (40 %), KCL (20 %), fosfato diamónico (11 %), Sulcamag (11 %) y Borozinco (2 %) en dosis de 380 g.planta-1 (Castañeda-Garzón et al., 2025). En el periodo 2013 a 2024, las condiciones climáticas promedio de La Libertad, registradas por la estación meteo- rológica, incluyeron una precipitación media anual de 2703 mm, una temperatura de 26 °C y una humedad relativa de 79 %. En el CCGE Taluma, las condiciones fueron la siguientes: precipitación media anual de 2975 mm, temperatura de 31 °C y humedad relativa de 81 % (NASA, 2025). En ambas localidades, la temporada seca ocurrió entre diciembre y febrero-marzo (Castañeda-Garzón et al., 2025).
Muestreo y estimación de biomasa aérea
Se midió la circunferencia del tronco (CIRC) del 100 % de los árboles (edad: 11 años) del CCPE La Libertad a un metro sobre el nivel del suelo, utilizando una cinta métrica (Rivano et al., 2013). Posteriormente, se calculó el área basal para determinar el diámetro cuadrático (Dg), según se muestra en la Ecuación (1), a partir del cual se seleccionaron tres árboles que estuvieran lo más cerca del Dg por clon (total: 27 individuos).
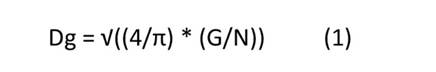
donde: Dg: diámetro medio cuadrático (m), G: área basal (m2), N: número de árboles por parcela, π: constan- te, 3.1416.
Seguidamente, se realizó un muestreo directo de biomasa aérea (BA) de los 27 árboles seleccionados. Tam- bién se registró la altura de la copa y se determinó la totalidad de su peso húmedo, considerando cuatro componentes: tronco (con corteza), ramas (diámetro>5 cm), ramitas (0 cm<diámetro<5 cm) y hojas. De cada componente, se tomó una muestra de aproximadamente 500 g para determinar el peso húmedo y secarla en horno a una temperatura constante de 70 °C hasta obtener un peso constante (peso seco). El contenido de carbono de cada componente por árbol (108 muestras) se determinó por medio del método de combustión de vía seca, utilizando el analizador elemental Flash 2000 CHNS (Thermo Fisher Scientific) en el laboratorio de química del Centro de Investigación Tibaitatá de Agrosavia. A partir del porcentaje de carbono obtenido en laboratorio por componente y por árbol, se determinó el contenido de carbono (kg), multiplicando este valor por la biomasa para obtener la fracción de carbono (FC). Se generó el valor promedio de FC para los 27 individuos derribados.
Para lograr mayor precisión e identificar mejor el patrón típico de las relaciones alométricas entre la circunferen- cia del tronco y la BA (ley alométrica de escalamiento morfológico) (Sileshi, 2014), se cuantificó la BA de 47 árbo- les adicionales en pie (CIRC<30 y >90 cm), representados por los nueve clones estudiados, debido a que no fue posible derribar más individuos, pues se habría modificado el diseño del lote. Se midió la altura del fuste limpio
(m) y el diámetro a la altura del pecho (DAP, cm) de cada individuo en pie seleccionado. Así, la biomasa del fuste se determinó como el producto del volumen del individuo por la densidad de la madera (Fonseca et al., 2021).
Posteriormente, se calculó la BA de 74 árboles multiplicando la biomasa del fuste por el factor de expansión pro- medio para los árboles de caucho. El factor de expansión de la biomasa y la densidad de la madera usada para la especie se obtuvieron a partir de los 27 árboles evaluados mediante el método directo.
Ajuste de la ecuación alométrica para estimar la biomasa aérea
De acuerdo con la naturaleza de las relaciones alométricas, es común que la dispersión de los puntos que re- lacionan la biomasa con la circunferencia del tronco (CIRC) varíe más en ambas variables a medida que sus va- lores aumentan. Por ello, es recomendable seleccionar árboles de tamaños amplios para lograr el patrón no lineal típico de la teoría de escalamiento metabólico. La biomasa de un árbol está vinculada al diámetro de su tronco por una relación de potencia con un exponente igual a 8/3 ≈ 2.67, según se expresa en la Ecuación (2).

donde: B: biomasa del árbol (biomasa aérea) (kg), α: constante de proporcionalidad o normalización, ρ: den- sidad específica de la madera (g.cm-3), r: constante que no depende del entorno donde crecen los árboles, D: diámetro del tronco (cm).
El ajuste de la ecuación para determinar la BA a partir de la CIRC en árboles de H. brasiliensis se basó en la ley alométrica de escalamiento morfológico (Sileshi, 2014), i.e., utilizó la función de potencia que se muestra en la Ecuación (3), la cual describe con gran precisión las relaciones alométricas de las plantas.
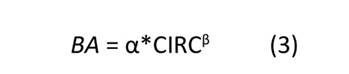
donde: BA: biomasa aérea (kg. árbol-1), α: constante de proporcionalidad o normalización, CIRC: circunferen- cia del tronco a 1 m sobre el suelo (cm), β: parámetro de escalamiento.
El escalamiento alométrico de las especies asume una función de potencia porque caracteriza el crecimiento como un proceso multiplicativo mediante una escala invariante y universal (Zianis & Mencuccini, 2004). Por lo tanto, se utilizaron métodos de ajustes de regresión no lineal, velando por el cumplimiento de los supuestos estadísticos y modelando la heterogeneidad de las varianzas de los residuales. Además, se incluyó el ajuste de la linealización de la función de potencia, i.e., se aplicó una transformación logarítmica con base en el valor de Euler a las variables BA y CIRC, como se muestra en la Ecuación (4).
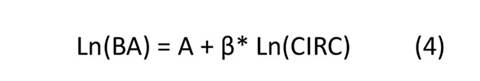
Las ecuaciones alométricas basadas en la función de potencia se ajustaron de la siguiente manera: a) regre- sión no lineal bajo el supuesto de residuales homogéneos (NL), b) regresión no lineal con residuales siguien- do una función de potencia (NL var potencia), c) regresión no lineal con residuales siguiendo una función exponencial (NL var exponencial) y d) regresión lineal con corrección por sesgo como modelo lineal genera- lizado (MLG gamma/log)—para el modelo de potencia con transformación logarítmica que se muestra en la Ecuación (4).
Los ajustes de los modelos a, b y c se realizaron con las funciones nls y gnls del paquete de R nlme (Pinheiro & Bates, 2000), mientras que el del modelo d utilizó la función glm (R Core Team, 2025), lo que permitió em- plear el modelo lineal generalizado y la función de enlace logaritmo con distribución de probabilidad gamma en los residuales (Zuur et al., 2013). La mejor ecuación alométrica se seleccionó considerando criterios de in- formación y visualización de residuales, tal como se ha hecho en otros estudios (Andrade et al., 2022).
Ajuste de funciones de acumulación de carbono
Las curvas de crecimiento son funciones matemáticas que permiten describir el crecimiento de las plantas en lo que respecta a varios de sus atributos (Archontoulis & Miguez, 2015). En este trabajo, las curvas de acumu- lación de carbono se estimaron a partir de los registros semestrales para CIRC en el 100 % de los árboles vivos del CCPE La Libertad y el CCGE Taluma, desde los 0.5 hasta los 11 años de edad. Así, con la ecuación alométri- ca ajustada para BA y la ecuación alométrica para biomasa subterránea (Bs) generada para H. brasiliensis en Colombia (Bs=0.022*circunferencia del tronco2.357) (Moreno et al., 2005), se calculó la biomasa total de cada árbol y cada clon. El contenido de carbono total por árbol se determinó multiplicando la biomasa total por la FC promedio determinada en este estudio.
Se ajustaron cuatro modelos de crecimiento no lineal—presentados en las Ecuaciones (5) a (8)—para estimar el valor máximo potencial de carbono alcanzado por cada clon (C , kg.árbol-1), el tiempo o la edad necesaria para alcanzar la tasa máxima de fijación de carbono absoluto (TM, años) y el valor estimado de la tasa máxima de fijación de carbono absoluto (FC , kg.año-1).
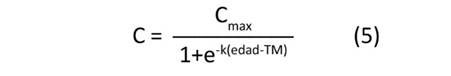
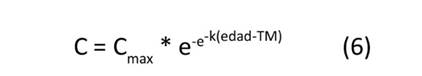
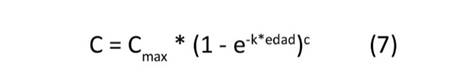
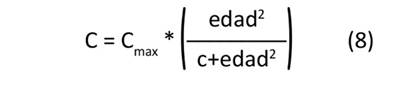
donde: Cmax: valor de carbono potencial máximo alcanzado por árbol, k: parámetro que controla la inclinación o forma de la curva, TM: tiempo o edad necesaria para alcanzar la tasa máxima de fijación de carbono (años), c: parámetro que influencia la velocidad a la que la función se aproxima al valor Cmax.
Los modelos de acumulación de carbono se ajustaron mediante regresión no lineal de efectos mixtos, utilizando el paquete nlme en el software R (R Core Team, 2025). Este enfoque se adoptó para mitigar la autocorrelación tempo- ral y la heterocedasticidad identificadas en las mediciones repetidas (CIRC vs. edad). Los parámetros del modelo se definieron como efectos fijos para cada clon, mientras que la variabilidad de dichos parámetros entre árboles indi- viduales se modeló como efecto aleatorio. La heterogeneidad residual persistente se corrigió incorporando una es- tructura de varianza de potencia (varPower). La selección del modelo óptimo para cada clon se realizó con base en los siguientes criterios de bondad de ajuste: (i) mayor coeficiente de determinación condicionado (R2), (ii) menores valores para los criterios de información de Akaike (AIC) y bayesiano (BIC), (iii) menor error estándar de los residuos.
A partir del mejor modelo de acumulación de carbono ajustado para cada clon se derivaron los parámetros de crecimiento (Cmax, TM, FCmax y el incremento medio anual a 11 años, i.e., IMA11). Posteriormente, para compa- rar el carbono acumulado (t C.ha-1) entre clones de H. brasiliensis en el CCPE La Libertad y el CCGE Taluma, se estimó la BA individual con la ecuación desarrollada en este estudio, a la cual se le adicionó la Bs (de raíces), calculada a partir de la siguiente ecuación: Bs = 0.022*circunferencia del tronco2.357 (Moreno et al., 2005). Para la conversión a carbono, se utilizó la FC promedio determinada en esta investigación.
Finalmente se empleó un análisis de varianza (ANOVA) de efectos mixtos para detectar diferencias significa- tivas en la captura de carbono entre clones, considerando el efecto de los bloques como aleatorio. Las com- paraciones múltiples entre las medias ajustadas se realizaron con el paquete emmeans, y todos los análisis estadísticos se ejecutaron en el software R.
RESULTADOS
Biomasa aérea
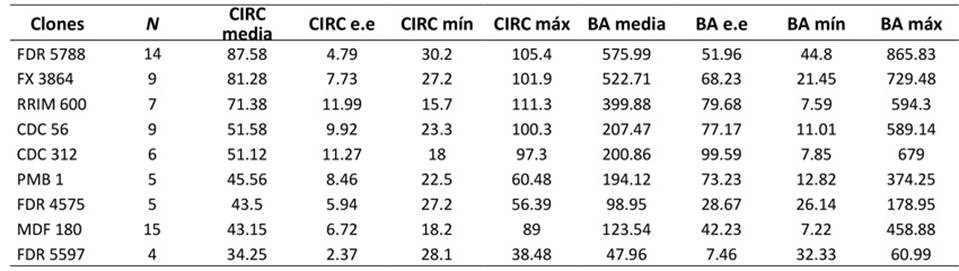
A la edad de 11 años, los clones de H. brasiliensis del CCPE La Libertad que presentaron mayor CIRC y BA promedio fueron FDR 5788 y FX 3864 (Tabla 1). En este estudio se determinó (i) una densidad básica de 0.54 g.cm-3 ± 0.14, (ii) un factor de expansión de biomasa (FEB) de 1.40 ± 0.09 y (iii) una FC de 0.485 ± 0.009. Se observó que la relación alométrica entre BA y CIRC fue creciente y no lineal, lo cual es característico de rela- ciones alométricas con un exponente mayor que 1. Los clones FDR 5788 y FX 3864 se destacaron por acumu- lar mayor BA para una misma CIRC.
Nota: CIRC: circunferencia del tronco a 1 m (cm), BA: biomasa aérea (kg.árbol-1), n: número de árboles, e.e: error estándar de la media, mín: valor mínimo registrado, máx: valor máximo registrado
Tabla 1: Estadísticas descriptivas para BA y CIRC en 74 árboles de H. brasiliensis del CCPE La Libertad (Villavicencio, Meta, Colombia)
Ajuste de funciones de acumulación de carbono
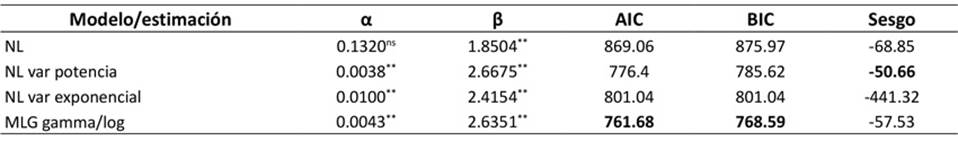
Con base en el AIC y el BIC, el modelo lineal generalizado gamma/log (MLG gamma/log) se identificó como el de mejor ajuste para estimar la BA a partir de la CIRC de los clones de H. brasiliensis estudiados. Aunque el modelo NL con varianza de potencia también mostró un buen desempeño, especialmente en la reducción del sesgo, el MLG gamma/log proporcionó un ajuste más parsimonioso y predictivo según los criterios de información (Tabla 2). También presentó la distribución de residuales más favorable y, aunque la heteroce- dasticidad es un desafío común en la modelación alométrica, mantuvo una cierta dispersión creciente de los residuales con el aumento de la CIRC.
Nota: NL: ajuste no lineal asumiendo homogeneidad de residuales, NL var potencia: ajuste no lineal incorporando un modelo de he terogeneidad de residuales de tipo potencia, NL var exponencial: ajuste no lineal incorporando un modelo de heterogeneidad de residuales de tipo exponencial, MLG gamma/log: regresión lineal con corrección por sesgo como modelo lineal generalizado, α y β: es timaciones de los parámetros de la función de potencia, AIC: criterio de información de Akaike, BIC: criterio de información bayesiano
Tabla 2: Modelos alométricos para estimar la BA a partir de la CIRC de árboles de H. brasiliensis en el CCPE La Libertad (Villavicencio, Meta, Colombia)
Acumulación de carbono
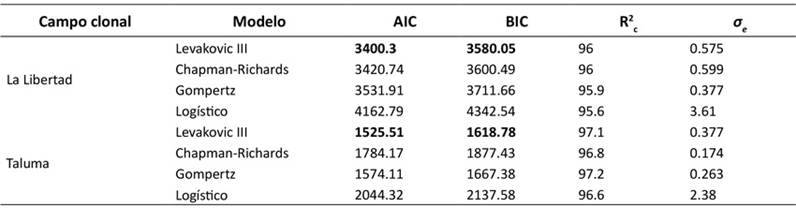
Una vez establecida la ecuación alométrica para la estimación de la BA, se ajustaron diferentes modelos de crecimiento (denominados modelos de acumulación de carbono en este estudio) para describir la dinámica de H. brasiliensis. Para el CCPE La Libertad y el CCGE Taluma, el modelo Levakovic III demostró ser el de mejor ajuste, pues presentó los valores más bajos de AIC y BIC, así como un R2 c alto (Tabla 3), describiendo mejor el patrón de acumulación de carbono en ambos sitios. Aunque el modelo Chapman-Richards también proporcionó un ajuste muy bueno, con valores de R2 c similares—y, en el caso de CCGE Taluma, un σϵ incluso menor—Levakovic III se destacó por su parsimonia y poder explicativo general. La consistencia en el rendimiento de estos modelos entre diferentes localidades sugiere su robustez para la modelación del crecimiento de carbono en H. brasiliensis.
Nota: AIC: criterio de información de Akaike, BIC: criterio de información bayesiano; R2 c: coeficiente de determinación condicional; σe: error estándar de los residuales
Tabla 3: Estadísticos de ajuste para modelos de acumulación de carbono de H. brasiliensis en el CCPE La Libertad y el CCGE Taluma (Villavicencio y Puerto López, Meta, Colombia)
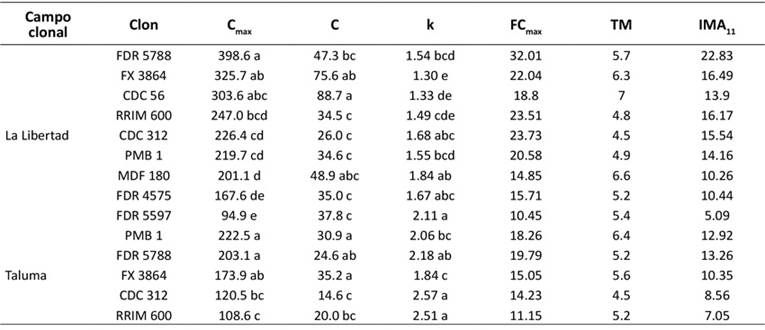
De acuerdo con los parámetros y métricas de crecimiento derivados del modelo Levakovic III, FDR 5788 y PMB 1 presentaron los valores más altos de Cmax en el CCPE La Libertad y el CCGE Taluma respectivamente. Sin embargo, FDR 5788 presentó la mayor FCmax y el IMA11 del carbono más en las dos localidades. A los 4.5 años, el clon CDC 312 alcanzó la mayor TM en las dos localidades, aunque su desempeño fue inferior en los demás parámetros de la curva de acumulación de carbono (Tabla 4).
Nota: Cmax: valor máximo de carbono estimado o asíntota de la curva de crecimiento (kg.árbol-1), k y c: parámetros de Levakovic III, FCmax: tasa máxima de fijación de carbono (kg.año-1), TM: edad del árbol en el cual alcanza la máxima tasa máxima de fijación de carbono (años) o punto de inflexión de la curva sigmoidal ajustada de Levakovic III, IMA11 : incremento medio anual del carbono a 11 años. Los valores de los parámetros de la ecuación de Levakovic III de cada clon que tienen la misma letra son estadísticamente iguales (5 % de significancia).
Tabla 4: Estimaciones de los parámetros del modelo Levakovic III para los clones de H. brasiliensis en el CCPE La Libertad y el CCGE Taluma (Villavicencio y Puerto López, Meta, Colombia)
El ANOVA y la posterior prueba de comparación de medias revelaron diferencias estadísticas significativas (p<0.05) entre los clones dentro de cada localidad (Figura 1). FDR 5788 presentó la mayor captura de carbono en el CCPE La Libertad (135.1 t C.ha-1), conformando un único grupo estadístico (a) y siendo significativamente superior a los demás clones evaluados. FDR 5597 registró el valor más bajo (grupo d). FX 3864, RRIM 600 y CDC 312 se ubicaron en el segundo grupo de alto rendimiento (b), seguidos por CDC 56 y PMB 1 en el grupo de transición (bc) y por MDF 180 y FDR 4575 en el grupo de captura moderada (c). Por el contrario, en el CCGE Taluma, la captura de carbono fue inferior. PMB 1 y FDR 5788 arrojaron los mejores resultados, sin diferencias significativas entre ellos (grupo a). RRIM 600 presentó la menor capacidad de captura (grupo c), siendo estadísticamente inferior, y los genotipos FX 3864 y CDC 312 reportaron valores intermedios (grupos ab y bc).
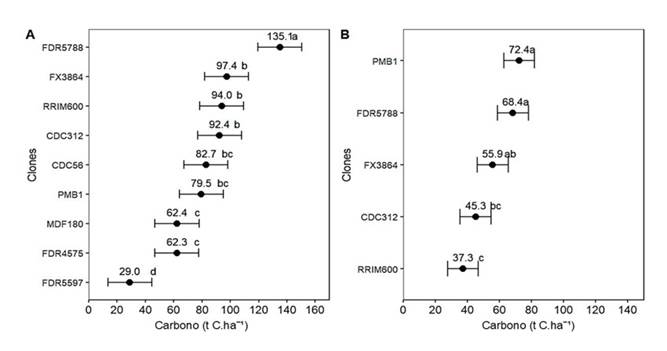
Figura 1: Comparación de captura de carbono (t C.ha-1) para diferentes clones de H. brasiliensis del CCPE La Libertad (A) y el CCGE Taluma (B) (Villavicencio y Puerto López, Meta, Colombia)
DISCUSIÓN
Biomasa aérea
Los valores promedio de BA en el CCPE La Libertad oscilaron entre 34.25 y 87.58 kg.árbol-1 en árboles de 11 años de FDR 5597 y FDR 5788 respectivamente (Tabla 1). FDR 5788 también sobresalió por su BA en un CCGE de nueve años en Caquetá (36.60 t.ha-1), superando a FDR 5597 (16.66), FDR 4575 (14.86), CDC 312 (14.21), CDC 56 (13.32) y MDF 180 (11.37) (SINCHI, 2019). En H. brasiliensis la BA varía según la edad, la ubicación geográfica y el sistema de cultivo (Moreno et al., 2005). Lo anterior se corrobora en reportes de otras plan- taciones, con valores de BA (kg.árbol-1) de 338.23 (nueve años, El Retorno, Caquetá), 91.57 (11 años, Milán, Caquetá), 395.28 (12 años, Cimitarra, Santander), 195.72 (12 años, Puerto López, Meta), 80.80 (13 años, Florencia, Caquetá), 710.94 y 187.36 (19 y 36 años, Doncello, Caquetá), así como en SAF en Doncello, Caquetá (0.1 y 68.4 kg.árbol-1, 0.5 y siete años), y en plantaciones de Caquetá (0.1 y 1.8 t.árbol-1), Brasil (18.69, 56.01 y 324.79 kg.árbol-1; cuatro, seis y 15 años) y México (73.5 t.ha-1) (Durán et al., 2011; Andrade et al., 2022; Maggiotto et al., 2014; López-Reyes et al., 2016).
Acumulación de carbono
La biomasa de las plantaciones de H. brasiliensis depende de varios factores, i.e., densidad de siembra, ma- nejo y edad, y el stock de carbono depende de la biomasa y su contenido de carbono (Hytönen et al., 2019). Generalmente se sugiere un valor de 0.5 para estimar el contenido de carbono en la BA (IPCC, 2006), pero se ha encontrado que asumir este valor genera errores cercanos al 5 % en las estimaciones de carbono forestal (Thomas & Martin, 2012), por lo que se considera necesario realizar estudios por especie. En efecto, la eva- luación de clones de H. brasiliensis en esta investigación arrojó que, a los 11 años de edad, la FC es de 0.485. Esto concuerda con lo reportado para plantaciones de RRIM 600 (20.25 y 30 años) en Tailandia (FC: 0.49); GT1 y PB235 (14 años) en Ghana Occidental, África, y Mato Grosso, Brasil (FC: 0.48); y varios clones (cinco edades) en Caquetá, Guaviare, Meta, Santander y Tolima, Colombia (FC: 0.44) (Hytönen et al., 2019; Wauters et al., 2008; Moreno et al., 2005). En un CCGE de Caquetá se reportaron valores de carbono almacenado (t C.ha-1) para CDC 312 (7.10), FDR 5597 (8.33), FDR 4575 (7.43), FDR 5788 (18.30), CDC 56 (6.66) y MDF 180 (5.68) (SINCHI, 2019). En Tabasco (México) y Río de Janeiro y Paraná (Brasil) los valores fueron los siguientes: 35.79, 50.90, 3.87, 11.79 y 52.68 t C.ha-1 (López-Reyes et al., 2016; Diniz et al., 2015; Maggiotto et al., 2014).
Otros estudios han estimado la biomasa de plantaciones de H. brasiliensis por medio de modelos alométricos generados a partir del diámetro o la circunferencia del tronco, la altura total o el diámetro de copa (Andrade et al., 2022; Blagodatsky et al., 2016; Brahma et al., 2017; Monroy & Návar, 2004; Moreno et al., 2005; Wauters et al., 2008). Sin embargo, se ha identificado una amplia variación en los parámetros de ecuaciones publicadas para la especie de estudio (Blagodatsky et al., 2016). En clones CMS y testigos comerciales, se han reportado valores entre 5.68 y 18.3 t C.ha-1 en Caquetá, 138.9 en Belén de los Andaquíes (Caquetá) y entre 26.10 y 65.30 t C.ha-1 en Brasil (SINCHI, 2019; Pardo-Rozo et al., 2021; Diniz et al., 2015).
Por ejemplo, en el CCPE La Libertad y el CCGE Taluma, el modelo Levakovic III (Tabla 3) arrojó los mayores re- sultados de acumulación de carbono por parte de H. brasiliensis (hasta 11 años) para FDR 5788 (135.1 t C.ha⁻¹) y PMB 1 (72.4 t C.ha⁻¹) (Figura 1). En las dos localidades, FX 3864 fue más eficiente que RIMM 600 en la captu- ra de carbono, CDC 312 presentó valores intermedios, y MDF 180, FDR 4575 y FDR 5597 obtuvieron menores valores, en línea con su menor vigor y tasa de crecimiento (Castañeda-Garzón et al., 2024, 2025). Por otra par- te, el CCPE La Libertad no solo presentó un rango más amplio de captura de carbono (29 a 135.1 t C.ha-¹) con respecto al CCGE Taluma (37.3 a 72.4 t C.ha-¹), sino que también albergó clones con un potencial de almace- namiento significativamente mayor (especialmente FDR 5788) (Figura 1). Estos resultados resaltan la impor- tancia de la selección clonal de H. brasiliensis para maximizar la acumulación de carbono por unidad de área.
Desde otra perspectiva, los parámetros del modelo Levakovic III (Tabla 4) revelaron que, en el CCPE La Liber- tad, el clon FDR 5788 presentó un mayor valor de carbono (398.6 kg.árbol-1) y una tasa máxima de fijación de carbono más alta (32.01 kg.año-1). En contraste, en el CCGE Taluma, PMB 1 presentó mayor carbono (222.5 kg.árbol-1) y una FC de 18.26 kg.año-1. En las dos localidades, FDR 5788 se destacó por alcanzar la FC a los cinco años, así como mayores IMA (22.83 y 13.26 kg.año-1).
Ahora bien, para H. brasiliensis, se indica que el genotipo (clon), la edad, el tipo de suelo, la topografía, la den- sidad de siembra, las prácticas de manejo y las condiciones edafoclimáticas son factores determinantes en la acumulación de carbono en plantaciones (Blagodatsky et al., 2016; Diniz et al., 2015; Mesike & Idoko, 2024). Incluso, se recomienda el uso de modelos locales, reduciendo la incertidumbre y facilitando la estimación de los créditos de carbono (Andrade et al., 2022). Los modelos aquí generados no solamente permiten a produc- tores de la Orinoquía estimar indirectamente la BA y la captura de carbono en los clones CMS, RRIM 600 y FX 3864 bajo condiciones agroecológicas similares a las presentadas en los campos clonales estudiados (CCPE La Libertad y CCGE Taluma), sino que también resaltan el potencial de la especie y robustecen la evaluación y selección de clones de H. brasiliensis en la Orinoquía.
CONCLUSIONES
El modelo alométrico de mejor ajuste para calcular la biomasa aérea de Hevea brasiliensis, diseñado a partir de valores de circunferencia del tronco, permitió identificar que, en el campo clonal La Libertad, los genotipos FDR 5788 y FX 3864 presentan el mejor desempeño hasta la edad de 11 años. Al integrar a este modelo los registros del campo clonal Taluma, fue posible estimar indirectamente la biomasa aérea por genotipo. Este modelo permite estimar con una mayor precisión el desempeño de clones CMS (series CDC, FDR, MDF y PMB), RRIM 600 y FX 3864 bajo condiciones edafoclimáticas similares en el piedemonte y la altillanura colombianos.
El modelo de acumulación de carbono de mejor ajuste fue Levakovic III, pues presentó los valores más bajos en los estadígrafos de Akaike y Bayes, así como el mayor valor de R2 c. Además, la estimación de los parámetros de la curva de acumulación de carbono complementó la descripción de genotipos de Hevea brasiliensis de los campos clonales La Libertad y Taluma (Villavicencio y Puerto López, Meta), y representa el primer hallazgo para clones CMS (CIRAD-Michelin Selection) en la región de la Orinoquía colombiana, resaltando el mejor desempeño de los clones FDR 5788 y PMB 1.
Las diferencias estadísticamente significativas entre clones de Hevea brasiliensis con respecto a la captura de carbono corroboran que la elección del genotipo es una estrategia para la mitigación del cambio climático. La identificación de clones de alto rendimiento es crucial para programas de mejoramiento y establecimiento de plantaciones con fines de captura de carbono en la Orinoquía.
Acknowledgements
Agradecimientos
Esta publicación se deriva de los resultados obtenidos en el proyecto Evaluación del comportamiento de clones de caucho en etapa productiva temprana en regiones caucheras de Colombia, ejecutado por Agrosavia en la Agenda Dinámica Corporativa, financiado con recursos públicos por medio del Ministerio de Agricultura y Desarrollo Rural de Colombia. Agradecemos al consorcio Coopération Internationale en Recherche Agronomique pour Le Développement (CIRAD, Michelin) por facilitar los clones CMS para su evaluación en Colombia. También agradecemos a Víctor Rodríguez, Misael Castañeda y Oscar Rendón (muestreo de biomasa), Hernán Camacho (evaluación en Taluma) y Jhon Zuluaga (metodología forestal).
Referencias
ANEXOS
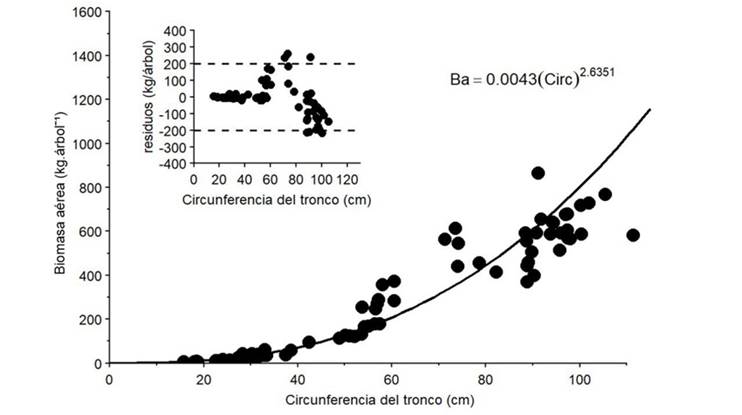
Anexo 1: Modelo alométrico de mejor ajuste y su correspondiente gráfico de residuales para estimar la BA a partir de valores de CIRC en árboles individuales de clones de H. brasiliensis del CCPE La Libertad (Villavicencio, Meta, Colombia)




Licencia
Derechos de autor 2025 Colombia forestal

Esta obra está bajo una licencia internacional Creative Commons Atribución-CompartirIgual 4.0.
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






