DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2010.1.a04Published:
2010-01-01Issue:
Vol. 13 No. 1 (2010): January-JuneSection:
Scientific articleDiversidad y estructura de un robledal en la reserva biológica Cachalú, Encino (SantanderColombia)
Diversity and structure of an oak community in the Cachalú Biological Reserve, Encino (SantanderColombia)
Keywords:
bosques de roble, diversidad, estructura de la vegetación, mortalidad y reclutamiento, parcelas permanentes, SantanderColombia. (es).Keywords:
oak forest, diversity, vegetation structure, mortality and recruitment, long-term plots, SantanderColombia. (en).Downloads
References
Álvarez, E., A. Cogollo, O. Melo, E. Rojas, D. Sánchez, O. Velásquez, E. Jiménez, D. Benítez, C. Velásquez, M. Serna, J. Pérez, F. Cardona & W. Devia. 2007. Monitoreo de los Andes colombianos (2000-3000 msnm) a través del establecimiento de parcelas permanentes, pp.: 75-89. En: Armenteras, D. & N. Rodríguez. (eds.). Monitoreo de los ecosistemas andinos 1995-2005: Síntesis y perspectivas. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá.
Ashton, P. S. & CTFS Working Group. 2004. Floristics and Vegetation of the Forest Dynamics Plots, pp.: 94-97. En: Losos, E. & E. Leigh. (eds.). Tropical Forest Diversity and Dynamism. Findings from a Large-Scale Plot Network. University of Chicago.
Barrera-L., J. & M. Diazgranados-C. 2006. Aspectos estructurales de tres bosques de roble ubicados en el área de amortiguación del Santuario de Fauna y Flora de Iguaque (Boyacá). Pontiicia Universidad Javeriana, pp.: 73-82. En: Solano C. & Vargas N. (eds.). Memorias del I Simposio Internacional de Robles y Ecosistemas Asociados. Fundación Natura - Pontiicia Universidad Javeriana. Bogotá.
Becerra, J. 1989. Estructura y crecimiento de un bosque secundario de Roble. Colombia Forestal 3: 60-64.
Cabrera, E. & D. P. Ramírez. 2007. Estado actual y cambio en los ecosistemas de los Andes colombianos: 1985-2005, pp.: 39-63. En: Armenteras, D. & N. Rodríguez, (eds.). Monitoreo de los ecosistemas andinos 1995-2005: Síntesis y perspectivas. Instituto de Investigación de recursos BiológicosAlexander von Humboldt. Bogotá.
Cantillo, E. E. 2007. La estructura de la vegetación en Colombia: una síntesis preliminar. Tesis de doctorado en Ecología. Instituto de Ciencias Naturales. Universidad Nacional de Colombia. Facultad de Ciencias. Bogotá. 539 p.
Cárdenas L., D. & N. R. Salinas. 2007. Roble: Quercus humboldtii Bonpl., pp.: 202-208. En: Cárdenas L., D. & N. R. Salinas (eds.). Libro rojo de plantas de Colombia. Volumen 4. Especies maderables amenazadas: primera parte. Serie libros rojos de especies amenazadas de Colombia. Instituto Amazónico de Investigaciones Cientíicas SINCHI- Ministerio de Ambiente, Vivienda y Desarrollo Territorial. Bogotá.
Cavelier, J., D. Lizcano, & M. T. Pulido, 2001. Colombia, pp.: 443-496. En: Kappelle M. & A. D. Brown (eds.). Bosques Nublados del Neotrópico. Instituto Nacional de Biodiversidad (INBio). Santo Domingo de Heredia. Costa Rica.
Chaparro, O. 2009. Caracterización estructural de los bosques andinos de la vertiente oriental de la Cuchilla El Fara, Santander. Trabajo de grado en Ingeniería Forestal. Universidad Distrital Francisco José de Caldas. Bogotá. 90 p.
Condit, R. 1998. Tropical forest census plots: Methods and results from BCI, Panamá and a comparison with other plots. Springer. Berlin. 211 p.
Condit, R., S. P. Hubbell, J. V. Lafrankie, R. Sukumar, N. Manokaran, R. B. Foster & P. S. Ashton. 1996. Species-area and speciesindividual relationship for tropical trees: a comparison of three 50-ha plots. Journal of Tropical cology 84: 549-562.
Condit, R., E. Leigh Jr, S. Loo de Lao & CTFS Working Group. 2004. Species-area relationship and diversity mesaures in the forest dynamics plots, pp.: 79-81. En: Losos, E. & E. Leigh, (eds.). Tropical Forest Diversity and Dynamism. Findings from a Large-Scale Plot Network. University of Chicago. Chicago.
Cortés-B., L. 2009. Caracterización de la vegetación de cuatro bosques de Roble ubicados en las veredas de Patios Altos y Canadá, Municipio de Encino, Santander. Trabajo de grado en Biología. Pontiicia Universidad Javeriana. Facultad de Estudios Ambientales y Rurales. 128 p.
Duque, A. J., W. López, F. H. Moreno & J. Toro. 2008. Variabilidad estructural y ambiental en remanentes de bosques andinos, pp.: 17-33. En: León., J. D. (ed.). Ecología de bosques andinos. Experiencias de investigación. Universidad Nacional de Colombia. Medellín.
Etter, A. & W. Wyngaarden. 2000. Patterns of landscape transformation in Colombia, with emphasis in the Andean Region. Royal Sweedish Academy of Sciences. Ambio 29: 432-439.
Galindo-T., R., J. Betancur & J. Cadena-M. 2003. Estructura y composición lorística de cuatro bosques andinos del Santuario de Flora y Fauna Guanentá-Alto Río Fonce, Cordillera Oriental colombiana. Caldasia 25: 313-325.
Galindo-T., R., J. Betancur & H. Mendoza. 2006. Estructura y composición lorística de los bosques de roble en dos parques nacionales naturales en la Cordillera Oriental Colombiana, pp: 95-100. En: Solano C. & N. Vargas (eds.). Memorias del I Simposio Internacional de Robles y Ecosistemas Asociados. Fundación Natura - Pontiicia Universidad Javeriana. Bogotá.
Galindo-T., C. & D. F. Correa G. 2006. Caracterización isonómica de un relicto de bosque de roble ubicado en la vereda El Ocaso, municipio de Bojacá, pp.: 95-100. En: Solano C. & N. Vargas (eds.). Memorias del I Simposio Internacional de Robles y Ecosistemas Asociados. Fundación Natura - Pontiicia Universidad Javeriana. Bogotá.
Gentry, A. H. 1982. Neotropical Floristic Diversity: phytogeographical connections between Central and South America, Pleistocene climatic luctuations, or an accident of the Andean Orogeny? Annals of the Missouri Botanical Garden 69: 557-593.
Gentry, A. H. 1995. Patterns of diversity and loristic composition in Neotropical Montane Forest, pp.: 103-126. En: Churchill, S. P., H. Balslev, E. Forero, J. L. Luteyn (eds.). Biodiversity and Conservation of Neotropical Montane Forests. Proceedings of the Neotropical Montane Forest Biodiversity and Conservation Symposium. The New York Botanical Garden. Bronx. New York.
González, A. 2009. Producción de frutos del roble Quercus humboldtii Bonpl., en dos bosques andinos de la cordillera oriental. Trabajo de grado en Ingeniería Forestal. Universidad Distrital Francisco José de Caldas. Bogotá. 37p.
González, C., A. Jarvis & J. D. Palacio. 2006. Biogeography of the Colombian oak, Quercus humboldtii Bonpl.: geographical distribution and their climatic adaptation. International Centre for Tropical Agriculture (CIAT)/Museo de Historia Natural, Universidad del Cauca. Popayán. 10 p.
Hooghiemstra, H. & A. M. Cleef. 1993. Pleistocene climatic change and environmental and generic dynamics in the North Andean Montane Forest and Paramo, pp.: 35-49. En: Churchill, S. P., H. Balslev, E. Forero, J. L. Luteyn (eds.). Biodiversity and Conservation of Neotropical Montane Forests. Proceedings of the Neotropical Montane Forest Biodiversity and Conservation Symposium. The New York Botanical Garden. Bronx. New York.
Hooghiemstra, H., T. Van der Hammen & A. M. Cleef. 2002. Paleoecología de la lora boscosa, pp.: 43-58. En: Guariguata E. & G. Kattan (eds.). Ecología y conservación de bosques neotropicales. Editorial Tecnológica de Costa Rica. Costa Rica.
Ingeominas. 1998. Mapa geológico generalizado del Departamento de Santander. Bogotá.
Jiménez A. 2009. Caracterización isionómica y estructural de la vegetación natural en claros de roble, en la reserva biológica Cachalú, Municipio de Encino (Santander). Trabajo de grado en Ingeniería Forestal. Universidad Distrital Francisco José de Caldas. Bogotá. 63 p.
Kappelle, M. 1992. Phytogeography of Talamanca montane Quercus forests. Journal of Biogeography 19: 219-315.
Kappelle, M. 2006. Changes in diversity and structure along a succesional gradient in a Costa Rican Montane Oak Forest, pp.: 223-233. En: Kappelle, M. (ed.). Ecology and conservation of Neotropical Montane Oak Forests. Ecological Studies, Vol. 185. Springer-Verlag. Berlin, Heidelberg.
Kappelle, M., A. Cleef & A. Chaverri. 1989. Phytosociology of montane Chusquea-Quercus forests, Cordillera de Talamanca, Costa Rica. Brenesia 32: 73-105.
Kappelle, M., T. Geuze, M. Leal & A. Cleef. 1996. Succesional age and forest structure in a Costa Rica Upper Montane Quercus Forest. Journal of Tropical Ecology 12:681-698.
Kessler, M. & J. Kluge. 2008. Diversity and endemism in tropical montane forests-from patterns to processes, pp.: 35-50. En: Gradstein, S. R., J. Homeier & D. Gansert. (eds.). The Tropical Mountain Forest. Patterns and processes in a Biodiversity Hotspot. Göttingen Centre for Biodiversity and Ecology. Biodiversity and Ecology Series 2.
López, W. & A. Duque. 2008. Diversidad alfa de especies arbóreas en remanentes de bosques montanos en la región norte de la Cordillera Central de Colombia, pp.: 35-56. En: León., J. D. (ed.). Ecología de bosques andinos. Experiencias de investigación. Universidad Nacional de Colombia. Medellín.
Losos, E. & CTFS Working Group. 2004. The structure of tropical forests, pp.: 69-78. En: Losos, E. & E. Leigh, (eds.). Tropical Forest Diversity and Dynamism. Findings from a Large-Scale Plot Network. University of Chicago. Chicago.
Lozano, G. & H. Torres. 1965. Estudio itosociológico de un bosque de Roble (Quercus humboldtii Bonpl.) de la Merced, Cundinamarca. Trabajo de grado Biología. Universidad Nacional de Colombia. Bogotá. 216 p.
Lozano, G., N. Ruíz, F. González & M. T. Murillo. 1996. Diversidad vegetal del Parque Nacional Natural Munchique, Cauca (Colombia), entre 1800 y 3050 msnm, pp.: 8-71. En: Pinto, P. (ed.). Notas sobre biodiversidad. Instituto de Ciencias Naturales-Museo de Historia Natural. Bogotá.
Magurran, A. 1989. Diversidad ecológica y su medición. Ediciones Vedra. Barcelona. 200 p.
Marín-C., C. & J. Betancur. 1997. Estudio lorístico en un robledal del Santuario de Fauna y Flora de Iguaque (Boyacá, Colombia). Revista de la Academia Colombiana de Ciencias 21: 249-259.
Mendoza-Cifuentes, H. & B. Ramírez-Padilla. 2000. Flora de La Planada: Guía Ilustrada de Familias y Géneros. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Fundación para la Educación Superior, World Wildlife Fund. Bogotá.
Ocaña, Y. 2005. Caracterización lorística y estructural de unidades de bosque andino en las veredas Minas y Patios Altos del municipio de Encino, Santander. Colombia Forestal 9: 7086.
Pan-Amazonian. 2009. Project for the advancement of networked science in Amazonian, Protocols for ield measurements at Caxiunana, Brasil and Tambopata, Perú (Preliminary version). 14 p. Disponible en: http://www.eci. ox.ac.uk./panamazonia/rainfor_ield_manual_ spanish.pdf
Paredes, A. & A. Martínez. 2005. Diversidad y caracterización lorística y estructural de la vegetación secundaria de la reserva biológica Cachalú, con propósitos de restauración. (Encino Santander). Trabajo de grado. Ingeniería Forestal. Facultad del Medio Ambiente y Recursos Naturales. Universidad Distrital Francisco José de Caldas. Bogotá. 238 p.
Phillips, O. & J. Miller. 2002. Global patterns of plant diversity: Alwyn H. Gentry's forest transect data set. Missouri Botanical Garden Press. St. Louis. 186 p.
Pulido, M. T., J. Cavelier & P. Cortés. 2006. Structure and composition of Colombian Montane Oak Forests, pp.: 141-151. En: Kappelle, M. (ed.). Ecology and Conservation of Neotropical Montane Oak Forests. Ecological studies 185. Springer-Verlag. Berlin Heidelberg.
Rangel-Ch., O. & A. Garzón. 2000. Aspectos de la estructura, de la diversidad y de la dinámica de la vegetación del Parque Regional Natural Ucumarí: Instituto de Ciencias Naturales. Universidad Nacional de Colombia. Bogotá.
Rangel-Ch., O., P. D. Lowry-C., M. Aguilar-P. & A. Garzón-C. 1997. Tipos de vegetación en Colombia. Una aproximación al conocimiento de la terminología itosociológica, itoecológica y de uso común, pp.: 59-87. En: Rangel-Ch., O., P. D. Lowry-C., M. Aguilar-P. (eds.). Colombia Diversidad Biótica II. Tipos de vegetación en Colombia. Instituto de Ciencias Naturales. Universidad Nacional de Colombia. Bogotá.
Rangel-Ch., O. & G. Lozano. 1986. Un peril de vegetación entre la Plata (Huila y el Volcán Puracé). Caldasia 14: 503-547.
Rangel-Ch., O., & A. Velásquez. 1997. Métodos de estudio de la vegetación, pp.: 59-88. En: RangelCh., O., P. D. Lowry-C., M. Aguilar-P. (eds.). Colombia Diversidad Biótica II. Tipos de vegetación en Colombia. Instituto de Ciencias Naturales. Universidad Nacional de Colombia. Bogotá.
Rincón, A. & N. Bernal, 2007. Factores antrópicos asociados e interrelaciones con el estado de los ecosistemas andinos, pp.: 10-128. En: Armenteras, D. & N. Rodríguez, (eds.). Monitoreo de los ecosistemas andinos 1995-2005: Síntesis y perspectivas. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá.
Samper K., C. & M. I. Vallejo. 2007. Estructura y dinámica de poblaciones de plantas en un bosque andino. Revista de la Academia Colombiana de Ciencias 31: 57-68.
Solano, C. 2006. Reserva Biológica Cachalú: 10 años de investigación en bosques de roble, pp.: 11-23. En: Solano, C. & N. Vargas (eds.). Memorias del I Simposio Internacional de Roble y Ecosistemas Asociados. Fundación Natura Pontiicia Universidad Javeriana. Bogotá.
Vallejo-Joyas M. I., Londoño-Vega A. C. LópezR. Camacho, G. Galeano, Álvarez-E. Dávila & Devia-W. Álvarez 2005. Establecimiento de parcelas permanentes en bosques de Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá. 310 p.
Vallejo, M. I. & G. Galeano. 2009. Cambios a corto plazo en los patrones de distribución espacial en nueve especies de plantas comunes en un bosque nublado al sur-occidente de Colombia. Caldasia 31: 76-98.
Van der Hammen, T. & O. Rangel-Ch. 1997. El estudio de la vegetación en Colombia, pp.: 17-57. En: Rangel-Ch., O., P. D. Lowry-C., M. Aguilar-P. (eds). Colombia Diversidad Biótica II. Tipos de vegetación en Colombia. Instituto de Ciencias Naturales. Universidad Nacional de Colombia. Bogotá.
Van der Hammen, T., R. Jaramillo & M. T. Murillo. 2008. Oak Forests of the Andean Forest zone of the Eastern Cordillera of the Colombian Andes, pp.: 595-614. En: Stuttgart, J. (ed.).Studies in Tropical Andean Ecosystems 7. Universidad de Amsterdam. Amsterdam.
Villarreal H., M. Álvarez, S. Córdoba, F. Escobar, G. Fagua, F. Gast, H. Mendoza, M. Ospina & A. M. Umaña. 2006. Manual de métodos para el desarrollo de inventarios de biodiversidad. Programa de Inventarios de Biodiversidad. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá. 236 p.
Zernig, K. & J. Betancur. 1994. Flora de Iguaque. pp.: 93. En: Cavelier, J., & A. Uribe (eds.). Resúmenes del Simposio Nacional "Diversidad Biológica, Conservación y Manejo de los Ecosistemas de Montaña de Colombia. Universidad de los Andes. Bogotá.
How to Cite
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Download Citation
 |
Ávila, F.A., Ángel, S.P., & López-C., R. (2010). Diversidad y estructura de un robledal en la Reserva Biológica Cachalú, Encino (Santander-Colombia). Colombia Forestal, 13(1), 87-116. |
DIVERSIDAD Y ESTRUCTURA DE UN ROBLEDAL EN LA RESERVA BIOLÓGICA CACHALÚ, ENCINO (SANTANDER–COLOMBIA)
Diversity and structure of an oak community in the Cachalú Biological Reserve, Encino (Santander–Colombia)
Diversidade e estrutura de um robledal na reserva biológica Cachalú, Encino (Santander-Colômbia)
Fabio A. Ávila1, Sonia Patricia Ángel2 & René López C.3
1Universidad Distrital Francisco José de Caldas.
fabio.botaniske@gmail.com. Autor para correspondencia.
2Universidad Distrital Francisco José de Caldas.
spangel00@gmail.com
3Universidad Distrital Francisco José de Caldas.
rlopezc@udistrital.edu.co
Recepción: Septiembre 22 de 2009/Aprobación: Noviembre 3 de 2019
RESUMEN
Se analiza la diversidad, la estructura y los aspectos relacionados con la dinámica (datos preliminares de mortalidad y de reclutamiento) de un robledal, provenientes de una parcela permanente de 1 ha, en la Reserva Biológica Cachalú, entre 2100 y 2200 m de altitud. Durante el establecimiento en el 2007 se marcaron, midieron y mapearon los individuos con DAP≥10 cm (primera categoría), y en 0.2 ha con 1≥ DAP< 10 cm (segunda categoría). Durante el primer censo (julio de 2009) se encontraron 453 individuos, 34 especies, 26 géneros y 23 familias en la primera categoría y 624 individuos, 80 es pecies, 60 géneros y 34 familias en la segunda. La curva especies-área mostró un valor R 2=0.97. En estructura, la distribución de diámetros tuvo una evidente acumulación de individuos en las prime ras clases de tamaño que en su mayoría corresponden a arbolitos, mostrando una distribución de “J” invertida, lo cual evidencia los estados tempranos de crecimiento del bosque. Entre el establecimien to y el primer censo realizado (1.5 años) se encontró que el área basal disminuyó 1.61% (13.99 m2ha-1 a 13.76 m2ha-1) en la primera categoría y aumentó 36.96% (0.51 a 0.70 m2) en la segunda. Las especies más importantes de acuerdo al IVI fueron Quercus humboldtii, Clusia schomburgkiana y Blakea cuatrecasii. Las tasas de mortalidad y reclutamiento fueron de 0.96% y 1%, en la primera categoría y de 2.80% y 12.20% en la segunda. Du rante este periodo de tiempo se presentaron nuevos claros en la parcela, lo cual se refleja en la dismi nución del área basal total. A pesar de la similitud florística del robledal estudiado con otros bosques montanos neotropicales, difieren algunas características estructura y de dinámica.
Palabras clave: bosques de roble, diversidad, estructura de la vegetación, mortalidad y reclutamiento, parcelas permanentes, Santander– Colombia.
ABSTRACT
We analyzed diversity, structure and preliminary plant dynamics features (mortality and recruit ment) of a one ha permanent plot in an oak forest, at the Reserva Biológica Cachalú, at an altitude of between 2100 and 2200 m. The plot was established in 2007, when all plant individuals with dbh ≥ 10 cm (first category) were tagged, measured and plotted. All individuals 1 ≥ and < 10 cm dbh (second category) were sampled in an area of 0.2 ha within the plot. In the last census (July 2009), we found 453 individuals, 23 families, 26 genera and 34 species from the first Category and 624 individuals, 34 families, 60 genera and 80 species from the second. The species-area curve showed a value of R2= 0.97. According to the structure analysis, most of the individuals, mainly treelets, were accumulated in the first diameter, showing an up-turned “J” distribution, which illustrates early forest stages. Between the plot establishment and the first census (1.5 yr) the basal area of the first category decreased 1.61% while the second increased 36.6%. The most important species ac cording to IVI were Quercus humboldtii, Clusia schomburgkiana and Blakea cuatrecasii. Mortal ity and recruitment rates for the first category were 0.96% y 1%, respectively and in the second cate gory these values were 2.8% y 12.2% respectively. During 1.5 yr new gaps appeared within the plot, which is associated with the decrease in total basal area. Despite high similarity in species composi tion compared with other neotropical montane forests, the dynamics and structure of the community in the present study appear to be distinct.
Key words: oak forest, diversity, vegetation structure, mortality and recruitment, long-term plots, Santander– Colombia.
RESUMO
Se analiza a diversidade, a estrutura e os aspectos relacionados com a dinâmica (dados preliminares da mortalidade e do recrutamento) de um robledal, provenientes de uma parcela permanente de 1 ha, na Reserva Biológica Cachalú, entre 2100 e 2200 m de altitude. Durante o estabelecimento em 2007 se marcaram, mediram e mapearam os indivíduos com DAP≥10 cm (primera categoría), e em 0.2 ha com 1≥ DAP< 10 cm (segunda categoría). Durante o primeiro censo (julho de 2009) se encontraram 453 indivíduos, 34 espécies, 26 gêneros e 23 famílias na primeira categoría e 624 indivíduos, 80 espécies, 60 gêneros e 34 famílias na segunda. A curva espécies-área mostrou um valor de R2=0.97. Na estrutura, a distribuição de diámetros teve uma evidente acumulação de individuos nas primeiras clases de tamanho que na sua maioria correspondem a arvorezinhas, mostrando uma distribuição de “J” invertida, o qual evidencia os estados precoces de crescimento do bosque. Entre o estabelecimento e o primeiro censo realizado (1.5 anos) se encontrou que a área basal diminuiu 1.61%(13.99m2ha-1 a 13.76 m2ha-1) na primeira categoría e aumentou 36.96% (0.51 a 0.70 m²)na segunda. As espécies mais importantes de acordó ao IVI foram Quercus humboldtii, Clusia schomburgkiana e Blakea cuatrecasii. As taxas de mortalidade e recrutamento foram de 0.96% e 1%, na primeira categoria e de 2.80% e 12.20% e na segunda. Durante este período de tempo se apresentaram novos claros no lote, o quel reflete uma diminuição da área basal total. Apesar da semelhança floral do rebledal estudado com outros bosques montanhosos neotropicais, diferem algumas características estruturais e de dinâmica.
Palavras chave: Bosques de roble, diversidade, estrutura da vegetação, mortalidade e recrutamento, lotes permanentes, Santander- Colômbia
INTRODUCCIÓN
Los Andes en Colombia comprenden cerca del 25% de la superficie nacional, dentro de los cuales se estiman más de 150 ecosistemas diferentes (Cabrera & Ramírez 2007). Según Rincón & Bernal (2007), esta región ha estado sometida a procesos de inmigración y asentamientos humanos, convir tiéndose en un foco de presión debido a la demanda de servicios ambientales, económicos y culturales. Los avances en el conocimiento de la composición, la estructura y la función de estos ecosistemas, así como de las alternativas de manejo de especies aún son limitados, lo que ha contribuido a la homoge nización del paisaje, la erosión y la irregularidad hidrológica (Etter & Wyngaarden 2000, Cavelier et al. 2001).
A pesar del escenario anterior, aún se encuentran en el país áreas representativas de la flora andina relevantes por su diversidad e importancia ecológica. Una de las formaciones boscosas más repre sentativas de los bosques andinos lo constituyen los bosques dominados por roble (Quercus humboldtii Bonpl.) o robledales. Esta especie de origen holártico colonizó el noreste de los Andes cerca de 350000 años atrás (Hooghiemstra et al. 2002, Van der Hammen et al. 2008) generando bosques recientes dominantes entre los 1000 y 3600 m de altitud de las tres cordilleras.
Los robledales al igual que la mayoría de bosques andinos, fueron objeto de un intenso aprovecha miento y disminución de su área de distribución, razón por la cual su uso fue vedado. No obstante, actualmente se considera que la extracción puede realizarse por medio del levantamiento de vedas locales (e.g. resolución 0316 de 1974, resolución 1408 de 1975, resolución 096 de 2006). Teniendo en cuenta que el rango de distribución del roble ha sido reducido al 42% del original, Cárdenas & Sa linas (2007), categorizaron la especie como especie vulnerable (VU) de acuerdo con los estándares de la UICN.
Los estudios en robledales han abordado aspectos de diversidad asociada, estructura, silvicultura y algunos aspectos de dinámica. En cuanto a estructura, los trabajos adelantados en el Parque Nacio nal Natural los Nevados, La Plata-Huila, Sumapáz y Tatamá, han definido sintaxones referentes a la asociación de Q. humboldtii, con especies de los géneros Dodonaea, Clusia, Alfaroa, Drimys, principalmente (Rangel-Ch. & Lozano 1986, Rangel-Ch. et al. 1997, Rangel-Ch. & Garzón 2000, Phillips & Miller 2002, Pulido et al. 2006, Cantillo 2007, Van der Hammen et al. 2008). En cuanto a composi ción y diversidad asociadas en los bosques andinos a diferentes escalas ecológicas, se encuentran los adelantados por Gentry (1982, 1995); Kessler & Kluge (2008) y López & Duque (2008). En estos se ha destacado la importancia de las condiciones ambientales (e.g. clima, topografía, sustrato) como aspectos a tener en cuenta en la definición de posibles hipótesis acerca del comportamiento de es tos bosques en el tiempo (Lozano & Torres 1965, Becerra 1989, Galindo-T. et al. 2003, Barrera-L. & Díazgranados 2006, Galindo-T. & Correa 2006, González et al. 2006).
En la Cordillera Oriental se encuentran los bosques de roble más representativos del país, en los cuales se han realizado estudios de estructura y composición de la vegetación. Por ejemplo, los estudios de Lozano & Torres (1965), Marín & Betancur (1997), Phillips & Miller (2002), Galindo-T- et al. (2003), Barrera-L. & Diazgranados (2006), Galindo-T. & Correa-G. (2006) y Galindo-T- et al. (2006) confirman la dominancia de Q. humboldtii, en los bosques de Boyacá y Santander, contiguos o inmersos en el Santuario de Fauna y Flora de Iguaque.
A pesar del sin-número de estudios en robledales, los estudios a largo plazo han estado pobremente representados en los Andes con únicamente 75 ha bajo el esquema de parcelas permanentes (Álvarez et al. 2007). Las perspectivas en los estudios de largo plazo han involucrado temáticas de manejo de especies de importancia ecológica y de cambio climático. El caso de la Red Bosco fue de las principales contribuciones redescriptivas de diferentes ecosistemas del país (Álvarez et al. 2002 en: Vallejo et al. 2005). Un aporte relevante en tér minos de superficie se llevó a cabo en la parcela permanente de La Planada en el Departamento de Nariño, en donde se han descrito aspectos importantes de composición (Mendoza-Cifuentes & Ramí rez-Padilla 2000), estructura, crecimiento y patrones de distribución de especies (Samper & Vallejo 2007, Vallejo & Galeano 2009). Contribuciones desarrolladas en el departamento de Antioquia, han abor dado componentes de diversidad y de estructura asociados a variables ambientales, lo que ha permi tido comprender procesos ecológicos en los bosques de roble (Duque et al. 2008, López & Duque 2008).
Frente a la escasez de estudios dinámicos, en el marco del proyecto “Corredor de Conservación de Robles: una estrategia para la conservación y el manejo forestal en Colombia”, la Fundación Natura en convenio con la Universidad Distrital Francisco José de Caldas, adelanta una iniciativa enfocada en el establecimiento de parcelas permanentes de vegetación que permiten la generación de información básica relativa a la diversidad, la estructura y la dinámica de los robledales. En el presente estudio se evaluó la estructura y la composición florística de una hectárea de bosque de roble en la Reserva Biológica Cachalú–Santander, mediante el establecimiento de una parcela per manente, aportando también un primer escenario acerca de la dinámica de estos bosques.
El establecimiento de esta parcela permanente es de especial interés ya que los estudios estructurales y dinámicos son relevantes para entender la eco logía de los bosques de roble y generar directrices en el manejo y la conservación de la especie. Las principales preguntas de este estudio estaban relacionadas con conocer el estado de la estructura y la diversidad de una hectárea de bosque de roble, cómo se distribuye la diversidad al interior de la parcela y cómo es la dinámica de crecimiento basal de las especies y cómo varía la mortalidad y el crecimiento en el corto plazo.
METODOLOGÍA
El estudio se llevó a cabo en la Reserva Biológica Cachalú de la Fundación Natura, ubicada en la Cordillera Oriental, en el área de amortiguación del Santuario de Flora y Fauna de Guanentá–Alto Río Fonce, en el departamento de Santander, municipio de Encino, situado a 06°08’27” de latitud norte y 73°06’07” longitud oeste (Figura 1). Su altitud varía de 1820 a 2650 m, la precipitación es de 3000 mm anuales, con un régimen bimodal; la temperatura oscila entre 8 y 18° C, la nubosidad es frecuente, generando humedad con fenómenos de lluvia horizontal (Solano 2006). Estas condiciones corresponden a la zona de vida de bosque muy húmedo montano bajo (bmh-MB).
La región se encuentra sobre la formación Rosa Blanca compuesta en su parte inferior por capas de caliza y yeso con oolitos, ostrácodos y dolomitas; hacia la parte superior consta de areniscas y lo dolítas calcáreas. En la parte inferior se presentan depósitos evaporíticos como yeso y polihalita que indican una hipersalinidad y tranquilidad en las condiciones de depositación (Ingeominas 1998). Los suelos son jóvenes, con condiciones de acidez (pH= 3.88 ± 0.39) y textura arenosa principalmente (González en este número).
ESTABLECIMIENTO Y CENSOS DE LA PARCELA PERMANENTE
La parcela permanente se estableció en un rango altitudinal entre 2032 y 2199 m (06° 04’34.5”N– 73°07’56.4”O) en un bosque dominado por roble (Q. humboldtii), con cambios considerables de pendiente (31%–93%), dados principalmente por la presencia de una cuenca antigua en los primeros transectos. Es una zona de suelos jóvenes con alta presencia de rocas en proceso de meteorización.
En noviembre de 2007, se instaló la parcela con una estación topográfica Topcom®, de alta preci sión. Se trazó el perímetro y se ubicaron puntos cada 10 m de distancia, en los que se obtuvieron valores de altitud y coordenadas X y Y; cada uno de estos puntos se marcó empleando tubos blancos de PVC de ½”, y cada 20 metros tubos naranja de 2.5”. La parcela se dividió en 10 transectos de 0.1 ha, encerrando 10 cuadrantes de 100 m2, nombrados de A a J y de 1 a 10 (Figura 1).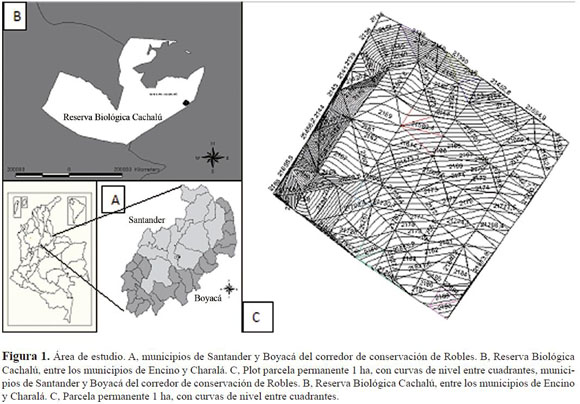
El período entre los dos censos fue de un año y medio. En el primer censo en 2007, se censaron todos los individuos con diámetro a la altura del pecho (DAP) ≥ 10 cm, se estimaron alturas totales (ht) y alturas fus tales (hf), diámetros de copa y se ubicaron dentro de los cuadrantes de 10 x 10 m. En dos transectos (B y H), equivalentes a 0.2 ha se censaron los individuos con 1≥ DAP < 10 cm con calibrador digital, se esti maron alturas, y se ubicaron en cada cuadrante. Adicionalmente, en las 0.2 ha, se describieron las formas de vida epífitas, hemiepífitas y lianescentes, en 6 unidades de 5 x 5 m. Se colectó al menos una muestra botánica de cada especie censada en la parcela bajo los números de colección AA, RL y SPA.
Todos los individuos medidos se marcaron según la metodología propuesta por Condit (1998). Esta metodología consiste en una marcación perimetral con pintura asfáltica de cada individuo a una altura de 1.3 m. También se instaló en cada individuo una placa metálica con la numeración en relieve. Para los individuos mayores se clavaron puntillas de acero de ½ pulgada y en los menores las placas se insertaron en una porción de alambre dulce. Con el fin de obtener mediciones homogéneas futuras y minimizar la desviación estándar de los diámetros de los individuos mayores, se instalaron dendrómetros elaborados de acuerdo manual metodológi co del proyecto PAN-AMAZONIAN (2009).
Se incluyeron casos especiales en los que sobresalen atributos de difícil medición, como lo establecen Vallejo et al. (2005). En individuos estranguladores, la medición se realizó en la porción superior libre. Los diámetros de individuos con tallos múltiples se tomaron en promedio. La medi ción en individuos bifurcados se realizó a la altura de 1.3 m y cuando coincidió con la bifurcación se tomó por encima. Los individuos con raíces tablares se midieron por encima de estas estructuras.
Las muestras botánicas se procesaron y se determinaron las especies con ayuda de claves en grupos especializados o por comparación con especímenes de las colecciones de los herbarios UDBC y COL. Algunas familias fueron determinadas por el equipo encargado de los catálogos preliminares de flora vascular (Medina et al. y Reina et al. en este número), en el marco del proyecto del corredor de conservación de roble.
PROCESAMIENTO DE DATOS
Para la distribución de la abundancia se realizaron curvas de acumulación de especies respecto al área y al número de individuos (Villareal et al. 2006). Con los valores de Alpha de Fisher y con la re lación especies-individuos (número Z), se analizó el comportamiento de la riqueza relativa, como lo exponen López & Duque (2008). Se utilizó el algoritmo de especies esperadas propuesta por Gentry (1995) para 0.1 ha, teniendo en cuenta la altitud.
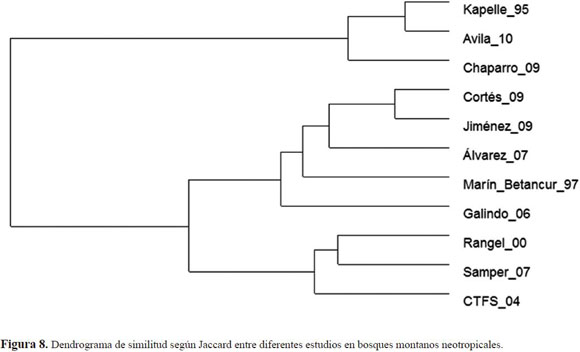
En diversidad, se tuvo en cuenta los índices de Shannon-Wiener (H’), trabajando con logaritmo base 10 y en algunos casos se calculó con base e para efectuar comparaciones con otros estudios, el índice de Simpson (1-D) y la serie logarítmica Alpha de Fisher (Magurran 1989), que evalúa eficazmente la diversidad en función del número de individuos y del número de especies (Condit et al. 1996). A partir de datos de ausencia y presencia a nivel de géneros, se elaboró un dendrograma de similitud con estudios realizados en bosques neotropicales montanos en 0.1 ha (Tabla 2).
En estructura se utilizaron los métodos propuestos por Rangel-Ch. & Velásquez (1997), para describir estratificación, dominancia, frecuencia, abundancia, cobertura e índice de importancia (IVI). Se siguió la clasificación de los individuos por estratos según la propuesta de Rangel-Ch. & Lozano (1986): Arbus tivo (Ar): 2-5 m; Arbolito (A): 5-12 m; Arbóreo inferior (Ai): 12-25 m. Se halló la pendiente promedio de cada cuadrante. Con base en las cotas registradas por la estación topográfica, se obtuvo un valor me dio por transecto, el cuál fue útil en el análisis de componentes principales.
Las variables utilizadas en dinámica fueron: mortalidad, reclutamiento y crecimiento, según las ecuaciones citadas por Samper & Vallejo (2007). El crecimiento se expresó como incremento medio anual (IMA) en centímetros de diámetro.
Se implementó un análisis factorial de componentes principales para encontrar correlación entre va riables según unidad de área (transectos) y según categoría taxonómica (especies), utilizando en éste último las diez especies con mayores valores de IVI (Anexo 2 y 3). Para el primer análisis, se utilizaron las variables de diversidad de Shannon (H’), área basal (G), pendiente (m), mortalidad (M), reclutamiento (R) e incremento medio anual (IMA). En el segundo caso, se emplearon número de individuos en categorías diamétricas uno, dos y tres (CI, CII, CIII) obtenidas en el programa Past® para los individuos, área basal (G), mortalidad (M), reclutamiento (R) e incremento medio anual (IMA). Los datos obtenidos en campo fueron digitalizados y procesados en los programas Excel®, Past® , SPAD® y ArcMap®.
RESULTADOS
COMPOSICIÓN FLORÍSTICA
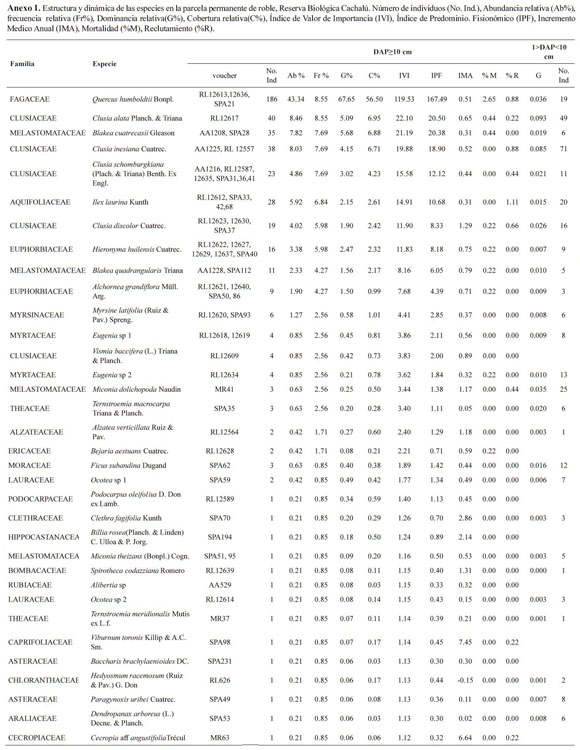
En el primer censo se registraron 452 individuos con DAP≥10 cm (primera categoría), representados en 23 familias, 26 géneros y 34 especies, se logró un porcentaje de determinación a nivel de especie de 87.5%. Las familias Fagaceae, Clusiaceae y Melastomataceae representaron un 78.58% de los in dividuos de la parcela. La familia más diversa en géneros y especies correspondió a Melastomataceae (2.4) seguida por Clusiaceae (1.4); los géneros más diversos fueron, Clusia con cuatro especies, seguidos de Blakea, Eugenia, Miconia, Ocotea y Ternstroemia, con dos especies. La abundancia del roble correspondió al 48.12% de la parcela. En el segundo censo se encontró el aporte de dos nuevas familias, Cecropiaceae y Caprifoliaceae, con las especies Cecropia aff. integrifolia y Viburnum toronis.
Con respecto a los individuos con 1≥DAP< 10 cm (segunda categoría), se encontró un total de 466 en los dos transectos (B: 255 y H: 211 individuos), representados en 65 especies, 49 géneros y 30 fami lias. Las familias más diversas según el número de especies fueron Ericaceae (8), Clusiaceae (4), Ru biaceae (3) y Araliaceae (3) (Anexo 1). Ericaceae, Lauraceae y Melastomataceae representaron gran diversidad de especies en la segunda categoría; se encontraron 58 especies en ésta categoría que no se encuentran registradas en la primera. Aquí, en el segundo censo ingresaron tres nuevas familias, Bombacaceae, Piperaceae y Cunoniaceae, se reclutaron 13 especies y 10 géneros.
Elementos florísticos relevantes presentes en la parcela permanente considerados como plantas no leñosas fueron los siguientes: Guzmania triangularis, Elaphoglossum spp., Epidendrum spp., Marcgraviastrum aff. gigantophyllum, Peperomia aguilae, Pleurothallis perijaensis, Diogenesia floribunda, Sphyrospermum cordifolium, Solanum argenteum y Stenospermation ellipticum.
DIVERSIDAD
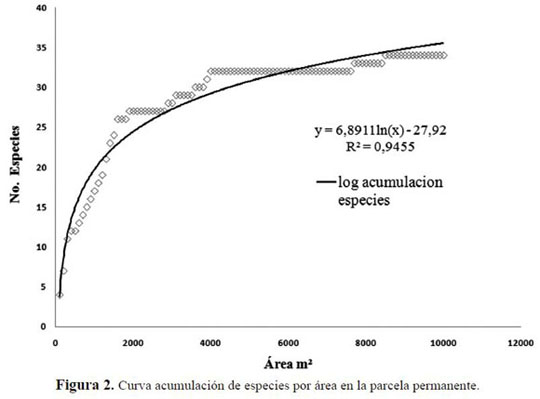
La curva de acumulación de especies respecto al área, mostró que aproximadamente para 6000 m2 se acumula el 96% de las especies, indicando que el área muestreada fue representativa para estimar la diversidad (Figura 2). La curva especies-área estuvo sustentada por un R cuadrado de 0.967 (p< 0.0001) y un análisis de residuales que no mostró tendencia alguna, lo cual permitió validar el modelo.
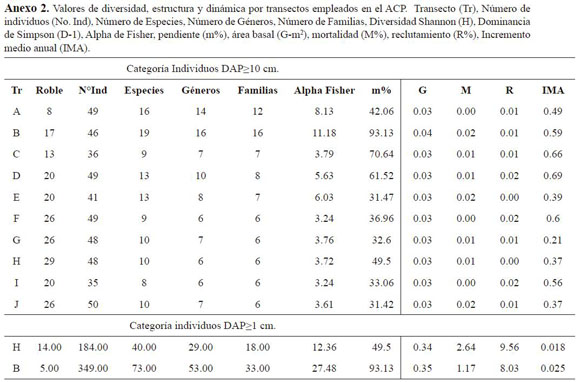
La riqueza relativa de la parcela permanente tuvo un comportamiento similar entre los individuos menores y los mayores. Para el caso del número z, expresado como la relación entre número de es pecies y de individuos en función del aumento del área, el incremento de especies se presentó a los ca. 400 individuos con DAP ≥10 cm y para la segunda categoría hasta casi. 600 individuos. A partir de este punto, el incremento describió un comportamiento asintótico, reflejado en el corte con el eje x (Figura 3). Del mismo modo, la relación entre la diversidad según Alpha de Fisher y la distribu ción de la abundancia en las especies, mostró un comportamiento similar al que presentó la curva de especies-área, ya que cerca del mismo número de individuos explica la diversidad de la parcela per manente (ca. 400 en la primera categoría y ca. 600 en la segunda). La diversidad según esta serie es de 31.42 para la segunda categoría. En la primera categoría se identificaron dos valores relacionados con la distribución de la pendiente en la parcela; los transectos A y B presentaron un Alpha de 8.1 y 11.2, respectivamente, mientras que los transectos de C a J, oscilaron entre 3.2 y 6.0 (Anexo 2).
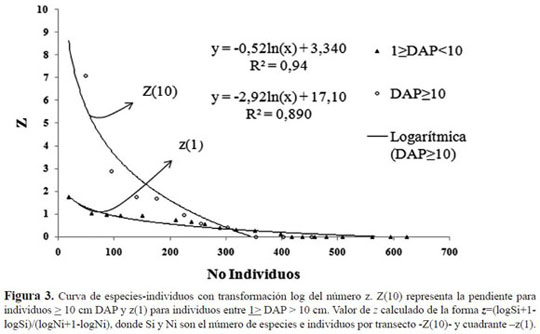
El análisis de similitud a partir de la abundancia de especies por transectos, mostró diferencias signifi cativas a lo largo de la parcela. Se encontró que los transectos A y B son los más diferentes con valo res inferiores al 50%. En esas dos franjas, estuvo concentrada la mayor diversidad de la parcela, los individuos de roble fueron menores y los valores de pendiente fueron más altos. Mientras tanto, en los transectos C a J, en los que coincidieron bajos valores de pendiente, se encontró mayor similitud con un porcentaje superior al 50%.
ESTRUCTURA
La distribución de diámetros para las dos categorías presentó un comportamiento de “J” invertida para los datos de establecimiento y primer censo, evidenciando la tendencia general de los bosques naturales en los primeros estadios de crecimiento (Figura 4a y b). Los incrementos de diámetro más relevantes fueron los de la primera categoría en las clases 2 y 3, así como para la segunda en las clases 1, 2 y 3. El diámetro promedio de la parcela permanente fue de 17.9±8.3 para los individuos ma yores; sin embargo, al considerar únicamente los individuos de roble, el diámetro promedio fue de 22.95±9.6 cm.
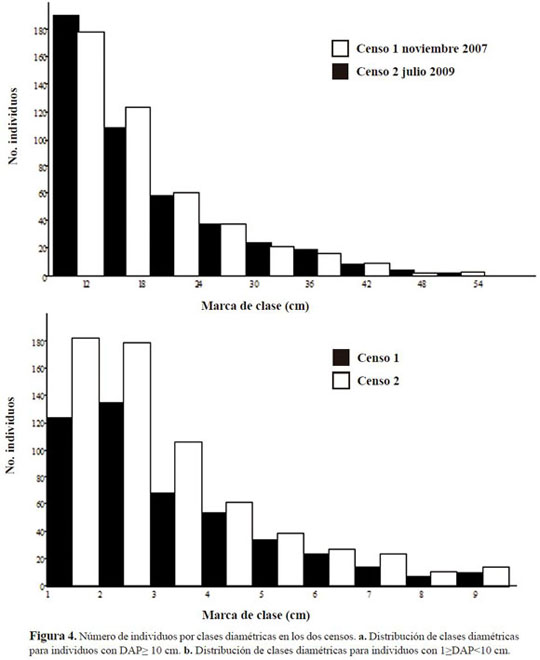
En cuanto a la estratificación, se encontró que en todos los transectos el estrato dominante fue el de arbolitos, seguido del estrato subarbóreo. Los elementos arbustivos fueron poco representativos, únicamente en Hieronyma huilensis y en Clusia schomburgkiana.
El área basal de la parcela permanente disminuyó 1.61%, pasando de 13.99 m 2 en el primer censo a 13.77 m2 en el segundo. Las familias más dominan tes fueron Fagaceae, Clusiaceae y Melastomataceae en la primera categoría, y Clusiaceae, Melastomata ceae, Ericaceae y Aquifoliaceae en la segunda.
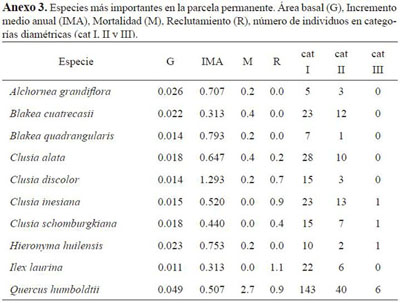
Las especies más importantes de acuerdo al IVI (Figura 5) corresponden a Quercus humboldtii (119.53), Clusia alata (22.10), Blakea cuatrecasii (21.19); estas también fueron las más importantes en el IPF. En los transectos con menor pendiente dominaron las especies Q. humboldtii, Ilex laurina Blakea quadrangularis.

DINÁMICA
En el segundo censo se encontraron 453 individuos con DAP ≥ 10 cm y se reclutaron dos nuevas espe cies Cecropia aff. angustifolia y Viburnum toronis. En los individuos menores, se reclutaron 13 nuevas especies, pertenecientes a 10 géneros y 4 nuevas familias. Las tasas de mortalidad y reclutamiento para los individuos mayores fueron de 0.96% y 1% respectivamente; mientras que para los indivi duos menores los valores fueron de 2.81% y 12.2% (Tabla 1).
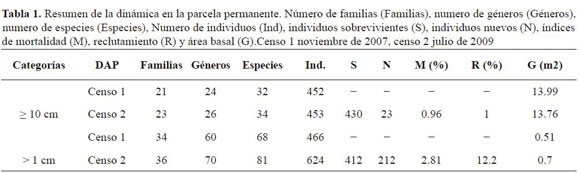
El mayor índice de mortalidad lo presentó Q. humboldtii con un total de 12 individuos muertos, se guido por Blakea cuatrecasii con 3 y Clusia alata con 2; por otra parte, Ilex laurina es la especie que registró mayor número de individuos recluta dos con un total de 5, seguida de Q. humboldtii y Clusia inesiana con 4. La mayor cifra de mortali dad se presentó en la categoría diamétrica II con 9 individuos. En la segunda categoría, el reclutamiento del transecto B fue mayor (109 individuos) que el encontrado en el H (91 individuos).
El análisis a nivel de especies muestra que las mayores tasas de reclutamiento fueron registradas para Clusia discolor e Ilex laurina, y las mayores tasas de mortalidad para A. grandiflora, Blakea quadrangularis, Hieronyma huilensis y C. discolor. En el incremento medio anual (IMA) sobresalieron las especies C. discolor (1.29 cm), B. quadrangularis (0.79 cm) y H. huilensis (0.75 cm). La tasa de mortalidad del roble fue mayor que la tasa de reclutamiento (proporción 3:1) y el valor de IMA fue de 0.50 cm (Anexo 2 y 3). En la categoría de individuos menores las especies Graffenrieda uribei, I. laurina y Miconia dolichopoda presentaron las mayores tasas de reclutamiento y A. grandiflora y B. quadrangularis las mayores de mortalidad.
CORRELACIÓN ENTRE VARIABLES
El análisis de componentes principales empleando las variables reclutamiento, mortalidad IMA, diversidad (H’), área basal y pendiente para los transec tos, permitieron explicar cerca del 76.59% de la inercia total de los ejes. Se lograron definir dos gru pos (Figura 6), uno conformado por los transectos I, F, D, C y A los cuales se encontraron asociados al eje 1 donde las variables, tasa de reclutamiento e IMA fueron las más influyentes sumado al aporte del transecto D. El eje 2, por el contrario, se definió por el efecto de la pendiente, la diversidad y la mortalidad sobre el transecto B, el cual presentó la mayor área basal, la mayor diversidad en transec tos y fue el de mayor pendiente.
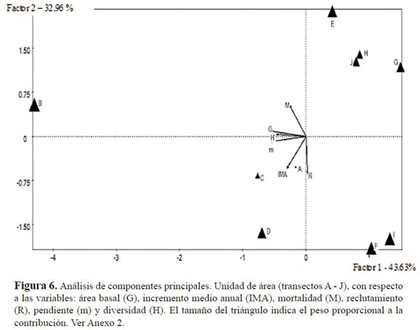
La relación entre las variables registradas en los anexos 2 y 3, mostró una gran influencia de la pen diente sobre las variables de diversidad y área basal. Se encontró que las variables relacionadas con la dinámica de la parcela mostraron un gran aporte a la varianza. La expresión de los componentes ob tenidos en las muestras, sugirió que los transectos A y B fueron los más diversos y los que tuvieron mayor influencia de la pendiente Así mismo, el área basal estuvo relacionada con la tasa de morta lidad, probablemente por la pérdida de individuos de grandes tallas. Del mismo modo, esta relación dependió del aporte de los individuos nuevos en área basal, que no alcanzaron los valores de los in dividuos muertos. Finalmente, el reclutamiento fue considerable en los transectos C y D, debido a la presencia de nuevos individuos de Ilex laurina
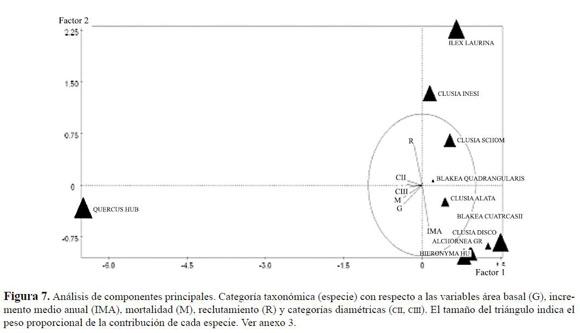
.El análisis factorial de componentes principales de las especies, muestra la tendencia de dos grupos (Figura 7); por un lado, las especies Clusia discolor, Hieronyma huilensis, Blakea quadrangularis y Alchornea grandiflora, influenciadas por el pri mer eje estuvieron relacionadas con el IMA. Otro grupo evidente lo conformaron las especies Clusia alata, Clusia schomburgkiana, Clusia inesiana e Ilex laurina , especies altamente relacionadas con las tasas de reclutamiento. Por su parte, el roble presentó gran influencia de las variables de número de individuos por intervalos de diámetro, mortali dad, área basal y pendiente. Factor que estuvo relacionado con la dispersión barocórica de la especie (González en este número).
DISCUSIÓN DE RESULTADOS
COMPOSICIÓN Y DIVERSIDAD
Varias características de orden fitogeográfico, geológico y ecológico de los bosques neotropicales montanos son compartidas por zonas que comprenden el cordón de montañas del centro de los andes (Bolivia-Perú) y las de Centro América (Nicaragua y Costa Rica). A pesar de que existen espe cies diferentes de roble en estos bosques montanos, las características de composición, diversidad y estructura presentan similitud (Kappelle 1992, Ashton & CTFS Group 2004). En Costa Rica en la Cordillera de Talamanca, se encontró una similitud considerable según la presencia y ausencia de géneros (Figura 8). Las principales semejanzas se encontraron en la presencia de Myrsine, Weinmannia, Oreopanax, Ilex, Nectandra y Ocotea. Cabe resaltar que el género Quercus tiene tres especies dominantes en Costa Rica: Q.costarricensis, Q. copeyensis y Q.seemanii mientras que en Colombia sólo está presente la especie Q. humboldtii (Kappelle et al. 1989, 1996, Pulido et al. 2006).
Se encontró gran similitud entre las familias dominantes de la parcela permanente y las de otros bos ques andinos del país (Figura 8). Los grupos, Clusia(Clusiaceae), Cavendishia, Disterigma, Psammisia, Satyria, Themystoclesia (Ericaceae), Ocotea, Persea (Lauraceae), Blakea, Graffenrieda, Miconia (Melastomataceae) y Alibertia, Hillia (Rubiaceae), estuvieron presentes en los estudios de Lozano et al. (1996), Marín & Betancur (1997), Álvarez et al. (2007), Duque et al. (2008). Además, los géneros Weinmannia (Cunoniaceae) y Monotropa (Monotropaceae), se han destacado en otros bosques andinos con domi nancia de roble (Becerra 1989, Rangel-Ch. & Lozano 1989, Marín & Betancur 1997, Barrera-L & Díazgra nados 2006, Galindo-T. & Correa 2006, Galindo-T. et al. 2006, Galindo-T. et al. 2003, Álvarez et al. 2007). Los géneros Cyathea (Cyatheaceae) y Styloceras (Buxaceae) ausentes en la parcela permanente, restaron similitud florística con otros bosques andinos del país estudiados por Phillips & Miller (2002).
La diversidad en bosques de roble fue mayor en tallas menores, y disminuyó a medida que se encontra ron elementos arbóreos. La abundancia de Quercus humboldtii y la baja diversidad, se debe posible mente a la alelopatía que crea ante otras especies, tal como lo exponen Lozano & Torres (1965). En la segunda categoría, la alta diversidad de la familia Ericaceae, coincidió con los estudios de Betancur & Marín (1997) y Zernig & Betancur (1994), en Iguaque, incluso con casi las mismas especies.
ESPECIES-ÁREA Y DISTRIBUCIÓN DE LA ABUNDANCIA DE ESPECIES
El análisis de la riqueza relativa, explicado en la tasa de incremento del número de especies respecto al número de individuos, indicó que eran necesarios menos de 400 individuos en la primera categoría y 600 en la segunda, para explicar la diversidad en la parcela. Al respecto, López & Duque (2008) en contraron que en una localidad de su estudio, donde incluso fue relevante la dominancia de Quercus humboldtii, era necesario un tamaño de muestra superior a 1000 individuos. Para este tipo de análi sis, es necesario considerar que los bosques andinos varían en su composición, presentando en algunos casos especies dominantes, lo cual permite suponer que la diversidad no depende sólo del área de mues treo, ni del tamaño de la muestra y que no siempre es acertado establecer un límite mínimo de individuos a muestrear para obtener el valor de diversidad.
El uso de la acumulación de la abundancia de las especies expresado en el número z, ayuda a explicar este tipo de comportamientos en los que la diversidad no depende del área, ni del tipo de muestreo, máxi me cuando las unidades de muestreo tienen efecto indirecto con la dinámica de poblaciones (Condit et al. 2004, Kessler & Kluge 2008, Rossenweig & Ziv 1999 en Kessler & Kluge 2008). El número Z y la acumulación de especies tuvo un comportamiento similar a lo registrado por López & Duque (2008), en la localidad de Belmira, a pesar de la diferencia en los tamaños de muestra con este estudio.
DIVERSIDAD
La diversidad en la segunda categoría, según Shannon, fue baja teniendo en cuenta que el va lor máximo fue de 1.14. En la Cordillera Oriental por ejemplo, los valores oscilan entre 0.98 y 4.42 para los sectores de Encino, Cachalú y Virolín en Santander, según lo encontrado en los estudios de estructura y florística de Ocaña (2005), Chaparro et al. (2009) y el de regeneración en claros en la Reserva Biológica Cachalú de Jiménez (2009). En el Santuario de Flora y Fauna de Iguaque, Marín & Betancur (1997), encontraron un valor de 1.14, Galindo-T. & Correa (2006) uno de 1.8 en Bojacá, Cundinamarca (Tabla 2).
Al analizar la diversidad según la serie logarítmica Alpha, se encontró un comportamiento similar. Tal es el caso de los valores registrados por Phillips & Miller (2002), ya que para la Cordillera Oriental, incluyendo individuos con DAP ≥2.5 cm, fueron mayores –Paramillo (21.74), Carpanta (30.52) y Perijá (30.08)– a los de la parcela permanente –primera categoría má;x-11.18 y segunda máx-27.48– (Tabla 2). Vallejo & Samper (2007), registraron alta diversidad en las dos categorías de diámetro: 29.31 para individuos con DAP ≥1 cm y 27.56 para los de DAP≥10 cm, lo que difiere de lo encontrado en la parcela permanente, con cambios de diversidad considerables entre categorías. Esto puede res ponder a la dominancia del roble, como se observó en el índice de Simpson con valores entre 0.6 y 0.9 (1-D) en la parcela.

El número de especies esperado según el algoritmo propuesto por Gentry (1995) para 0.1 ha, relacio nado con la altitud, estaría cerca de 107 especies. En la parcela permanente el máximo número de especies con DAP≥10 cm en 0.1 ha, fue de 21 y de 61 para los de 1≥DAP<10 cm. A pesar de que exista una relación entre el número de especies con la gradación de la altitud (Gentry 1995, Van der Hammen & Rangel 1997, Galindo et al. 2003), se deben considerar condiciones como la dominancia de especies, ya que estas pueden limitar las oportunidades de crecimiento de otras y limitar la expre sión de la diversidad (Pulido et al. 2006).
No sólo la diversidad en especies fue baja, ya que el número de familias géneros y especies escasamen te superó otros sitios, incluso en los que dominaba el roble. Valores inferiores a los de este estudio, los reportaron únicamente Galindo-T. & Correa (2006) con 8 familias para 0.1 ha. En términos generales para la zona y la para la cordillera oriental, se registraron valores superiores a 30 familias por 0.1 ha, y a 50 en 1 ha (Paredes & Martínez 2005, Álvarez et al. 2007, Chaparro et al. 2009, Jiménez 2009).
ESTRUCTURA
En este estudio el número de individuos por unidad de área (453 DAP ≥10 cm; 623, 1≤DAP>10 cm) es bajo si se compara con el promedio que se reporta para los bosques montanos tropicales (866 DAP ≥10 cm; ±1065, 1≤DAP>10 cm), (Losos & CTFS working Group 2004). Valores similares se encuentran en Costa Rica en los bosques de Quercus, en donde el número de individuos varía entre 455 y 555 en la categoría de DAP ≥10 cm (Tabla 3). Del mismo modo, al comparar por transectos de 0.1 ha, en la Cordillera de Talamanca – Costa Rica-, obtenidos por Kappelle (2006), se encontra ron valores de 45±6.54 individuos, y así mismo las condiciones de topografía y clima y algunas espe cies, coinciden en varios aspectos.

En el país, los valores encontrados por Álvarez et al. (2007), para las tres cordilleras -occidental (717), central (660) y oriental (809- son superiores a los de la parcela permanente. Mientras que el número registrado por Samper & Vallejo (2007) para La Planada (576±48) y Rangel-Ch. & Garzón (2000), para Ucumarí, no difieren de lo encontrado en la parcela permanente. Por otra parte, en el Santuario de Flora y Fauna Iguaque se registraron para la categoría de 1≤DAP>10 cm valores desde ±171 en la Sierra, has ta 745 en el sector de Chontales bajo (Galindo-T. et al. 2003) en una superficie de 0.1 ha.
Puntualmente, en la Reserva Biológica Cachalú, se han encontrado valores similares a los encon trados en este estudio. Se encontró que el número de individuos de roble en la parcela es superior a lo reportado por González (2009) para una hectárea, pero el diámetro promedio para los 121 individuos encontrados fue de 35.75±15.23 cm superior al de los individuos de roble de la parcela permanente (17.9±8.3 cm). El bajo número de individuos de roble, puede estar relacionado con lo expuesto por Guerrero et al. (en este número), al encontrar un patrón con tendencia a la uniformidad en plántulas de roble, lo que coincidió con el bajo número de individuos de roble en la segunda categoría en 0.1 ha (9 ind.), frente a la primera (20 ind.).
La distribución de diámetros es similar a la de otros estudios adelantados en robledales. Por ejemplo, González (2009), en la Reserva Biológica Cachalú, encontró también una distribución en “J” invertida, a pesar de haber encontrado pocos individuos en la primera clase diamétrica (11-20 cm), lo que pudo estar relacionado con el muestreo de individuos de roble únicamente. Sólo los resultados presentados por Cortés (2009), difieren del patrón anterior, pues algunos de estos bosques se encuentran en estados avanzados de sucesión. En la mayoría de estudios de bosque andino, la abundancia disminuye a me dida que aumentan los tamaños de diámetro (Rangel & Lozano 1989, Galindo et al. 2003, Barrera & Diazgranados 2006, Galindo & Correa 2006).
La estratificación de este estudio mostró una estructura vertical pobre. Esto se evidenció en un estrato arbóreo inferior escaso, teniendo en cuenta que las alturas del roble no superaron los 20 m. Re sultados similares, con alturas inferiores al estrato de arbolitos, los registraron Marín & Betancur (1997) en Iguaque. A pesar de que la presencia de estratos superiores es limitada en estos bosques an dinos, en localidades como La Plata en Huila, pueden ser los de mayor cobertura (Rangel & Lozano 1989). Por otra parte, Rangel & Garzón (2000) y Chaparro (2009) registraron dominancia del estra to arbóreo inferior.
Los valores de estructura horizontal también fueron bajos, al comparar por ejemplo el promedio de área basal para bosques montanos tropicales en una hectárea. Losos & CTFS Working Group (2004), registran valores entre 40.8 y 109.6 m2 para individuos con DAP≥ 10 cm y entre 92.6 y 134.7 m2 para individuos de 1≤DAP>10 cm. Del mismo modo, registran valores de 5.08 a 29 m 2 de área basal, para las cinco especies más importantes de cada parce la permanente, valores que incluso superan el área basal de este estudio. Esta baja dominancia estuvo relacionada con el dominio de una sola especie, con el estado sucesional del robledal y probable mente por la juventud suelo.
La dominancia también tuvo valores inferiores a los registrados en el país. En la parcela permanente el valor de área basal en este estudio pasó de 13.77 a 13.99 m2. Los valores encontrados en este estu dio corresponden a uno de los valores más bajos reportados para los bosques montanos, teniendo en cuenta que para la cordillera oriental se encontraron cifras entre 23.5 m2 y 27.3 m2 (Álvarez et al. 2007) y en La Planada de 23.41 m2 (Phillips & Miller 2002, Samper & Vallejo 2007).
Estos bajos valores pueden estar relacionados con dos aspectos. El primero tiene que ver con la baja es tratificación en los elementos arbóreos ya que se encuentran pocos individuos con alturas mayores a 12 m, y los diámetros no superan los 50 cm de DAP. El segundo corresponde a la interacción de condiciones del entorno como variables ambientales principalmente las características edáficas y a la topografía, que en este estudio condicionaron la estructura y la diversidad en algunos sectores de la parcela.
Los índices de importancia ecológica expresaron la relevancia del roble en este bosque. A pesar de la dominancia de ésta especie, la cobertura presentó valores que se consideraron bajos, debido a la baja densidad de copas. Rangel-Ch. & Lozano (1989), consideraron que mejores condiciones ambientales permitirían desarrollar atributos más sobresalientes. Es importante considerar que en pocos robledales el IVI del roble superó el 200% (Barrera & Diazgranados 2006). En este estudio el IVI de la especie fue de 119.53%, similar al valor del índice de la especie en otros sitios de la cordillera oriental que oscila entre 50 y 150% (Marín & Betancur 1997, Galindo-T. et al. 2003, Ocaña 2005, Galindo-T. et al. 2006, Galindo & Correa 2006, Cortés 2009).
En contraste con Costa Rica, la familia Fagaceae es la única que comparte valores de importancia con este estudio. En estos bosques de Centro América, los mayores valores en índices de importancia están dados por especies del género Quercus y de especies de las familias Aquifoliaceae, Symplocaceae, Theaceae y Juglandaceae. En este estudio los mayores aportes se encontraron en las familias Fa gaceae, Clusiaceae y Melastomataceae, las cuales representaron el mayor VIF para la parcela (Anexo 1). Aunque las familias Theaceae, Aquifoliaceae y Caprifoliaceae están en la parcela, no representa ron un mayor aporte al componente estructural.
DINÁMICA
Para la primera categoría, la diferencia que se presenta entre la tasa de mortalidad (0.96%) y la de reclutamiento (1%) definió un aparente equilibrio en la dinámica de la parcela. Por el contrario en la segunda categoría el cambio es amplio entre la tasa de reclutamiento (12.2%) y la de mortalidad (2.81%). Este comportamiento estuvo relacionado con las estrategias de reproducción de cada una de las especies, la aparición de claros, así como posibles condiciones en disponibilidad de espacio, luz, y recursos en general.
El valor de crecimiento para la parcela permanente fue mayor al de otro bosque andino. En la parcela permanente se encontraron valores de 0.73 cm/año en los individuos de categoría mayor y 0.81 cm/ año en la menor. En contraste, Samper & Vallejo (2007), registraron valores para La planada de 0.14 y 0.27 cm/año respectivamente.
Es importante tener presente que este valor de crecimiento superior es relativo, ya que depende del plazo en el que se desarrollan los censos, así como de las condiciones y estrategias que mantienen las plantas para supervivir y adaptarse. Un ejemplo de esto fue lo encontrado por Samper & Vallejo (2007) en casos en los que un individuo se considera muerto, pero en un censo posterior se ha rege nerado haciendo que ingrese un individuo nuevo a una clase diamétrica superior. Este nuestro estudio esto se observó con las especies Themystoclesia epyphytica e Ilex laurina.
En términos generales se pudo notar la dinámica de las especies del sotobosque, debido a los mayores valores en reclutamiento y mortalidad, al compararlos con los de la primera categoría. Es impor tante destacar los valores de reclutamiento de I. laurina, que en las dos categorías de individuos fue el mayor registrado; esto se debe probablemente a sus altos valores de abundancia y a su estrategia de propagación. El roble por su parte, en los individuos arbóreos, mantuvo valores constantes de área basal a pesar de que la mortalidad fuera la más alta; el valor de IMA registrado para ésta especie en la parcela fue de 0.5 cm/año, relativamente bajo si se tiene en cuenta el valor de 0.58 cm/año reportado por Becerra (1989).
CORRELACIÓN ENTRE VARIABLES
En escala geológica la estructura, la composición y la función de áreas naturales dependen de la va riación de las condiciones ambientales (Kessler & Kluge 2008). Dichas condiciones pueden afectar la presencia o no de elementos florísticos, como de sus atributos. La formación andina genera condiciones muy específicas y es posible encontrar fenómenos muy variables entre localidades relativamente cer canas. Al respecto, varios autores aseguran que la topografía y el clima son los generadores de de terminados patrones en las variables de estructura horizontal y vertical de las formaciones montaño sas del neotrópico (Hooghiemstra & Cleef 1993, Van der Hammen & Rangel 1997, Kessler & Kluge 2008). Losos & CTFS working group (2004), han encontrado que la densidad de individuos por cla se de tamaño no está relacionada con la pendiente promedio de cada parcela, aunque no desconocen que la topografía puede condicionar otros aspectos que llegaran a afectar la estructura (drenaje, lixi viación, interacción con vientos, entre otros).
La variación de estructura y composición estuvo reflejada en los cambios de pendiente a lo largo de la parcela, tal y como lo mostró el análisis de componentes principales (Figura 6). En este punto, la pendiente estuvo considerablemente relacionada con las variables de IMA y diversidad. Para el caso de la composición, se pudo obtener conclusiones preliminares, que pueden ser apoyadas con el estudio de otras condiciones específicas como el sustrato, microclima, patrones de distribución de especies, entre otros, ya que se logró observar la ausencia o presencia de especies según los cambios de pendiente en la parcela. El roble mostró una evidente correlación con la pendiente, aspecto obser vado en el aumento de individuos de la especie en cuanto se avanza del transecto A al J.
CONCLUSIONES
Se registró un bajo número de especies, géneros y familias para el área estudiada. Sin embargo, la composición florística de la parcela permanente es similar a la de otros bosques andinos con presencia de roble. Comportamiento encontrado no sólo en bosques de Colombia, sino también en los de Cen tro América.
Se comprobó que para este bosque el área evaluada de una hectárea fue suficiente para describir el comportamiento de la diversidad de este robledal. La dominancia de la especie Quercus humboldtii, favoreció el carácter asintótico de la línea de ten dencia de la acumulación de especies. Del mismo modo, se comprobó que el tamaño de la muestra fue significativo para describir la diversidad según el valor de Alpha de Fisher.
La mayor diversidad se presentó en los individuos de menor categoría diamétrica. Mientras tanto, en los individuos mayores se encontró que la diversidad estuvo correlacionada con la pendiente. En los transectos donde la pendiente promedio fue mayor se encontraron más especies.
El área basal encontrada fue baja en comparación con otros bosques montanos neotropicales. Esto debido a que la mayor parte de los individuos estuvo representado en sólo dos estratos.
Los índices de importancia son semejantes a los registrados en estudios de la Cordillera Oriental. Es común la presencia en estos índices de la especie Quercus humboldtii y especies de los géneros Clusia y Blakea.
En un año y medio se evidenciaron cambios en la parcela permanente. En los individuos mayores el número de individuos aumentó, pero el área basal disminuyó; así mismo, en la categoría diamétrica menor, el número de individuos aumentó, así como su área basal.
Las relaciones entre topografía, diversidad, estructura y dinámica, fueron evidentes según el análi sis de componentes principales. Se evidenció la influencia de la pendiente en la composición y la estructura (expresada en área basal). Se encontró que el roble fue abundante en sectores con me nores valores de pendiente, debido a la estrategia de dispersión de la especie y probablemente a los requerimientos de sustrato de la especie para establecerse.
AGRADECIMIENTOS
Esta investigación contó con la financiación y apoyo de la Fundación Natura, en el marco del Pro yecto de Conservación del Corredor de Robles, la Fundación Mac Arthur y la Universidad Distrital Francisco José de Caldas. Expresamos agradecimientos al personal científico y administrativo del Herbario Forestal de la Universidad Distrital (UDBC) y del Herbario Nacional Colombiano ( COL), así como al de la Fundación Natura, especialmente a Luis Mario Cárdenas. Del mismo modo agra decemos a Rocío Cortés, Agustín Rudas y a los evaluadores del manuscrito por sus acertados co mentarios. Finalmente a José Martínez y a Omaira Prieto por su apoyo permanente en el desarrollo del trabajo de campo.
REFERENCIAS BIBLIOGRÁFICAS
Álvarez, E. A. Cogollo, O. Melo, E. Rojas, D. Sánchez, O. Velásquez, E. Jiménez, D. Benítez, C. Velásquez, M. Serna, J. Pérez, F. Cardona & W. Devia. 2007. Monitoreo de los Andes colombianos (2000-3000 msnm) a través del establecimiento de parcelas permanentes, pp. : 75-89. En: Armenteras, D. & N. Rodríguez. (eds. ). Monitoreo de los ecosiste mas andinos 1995-2005: Síntesis y perspectivas. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá
Ashton, P.S. &CTFS Working Group. 2004. Floristics and Vegetation of the Forest Dynam ics Plots, pp.: 94-97. En: Losos, E &E. Leigh. (eds.)Tropical Forest Diversity and Dynamism. Findings from a Large-Scale Plot Network. University of Chicago
Barrera-L, J. &M. Diazgranados-C. 2006. Aspectos estructurales de tres bosques de roble ubicados en el área de amortiguación del Santuario de Fauna y Flora de Iguaque (Boyacá). Pontificia Universidad Javeriana, pp. 73-82. En: Solano C & Vargas N. (eds). Memorias del I Simposio Internacional de Robles y Ecosistemas Asociados. Fundación Natura - Ponti ficia Universidad Javeriana. Bogotá.
Becerra, J. 1989. Estructura y crecimiento de un bosque secundario de Roble. Colombia Forestal 3: 60-64.
Cabrera, E. &D.P. Ramírez. 2007. Estado actual y cambio en los ecosistemas de los Andes co lombianos: 1985-2005, pp.: 39-63. En: Armenteras, D &N. Rodríguez. (eds.)Monitoreo de los ecosistemas andinos 1995-2005: Síntesis y perspectivas. Instituto de Investigación de recursos BiológicosAlexander von Humboldt Bogotá
Cantillo, E.E. 2007. La estructura de la vegetación en Colombia: una síntesis preliminar. Tesis de doctorado en Ecología. Instituto de Ciencias Naturales. Universidad Nacional de Colombia. Facultad de Ciencias. Bogotá. 539 p.
Cárdenas, L.D. &N.R. Salinas. 2007. Roble: Quercus humboldtii Bonpl.. pp.: 202-208. En: Cárdenas L.D & N.R. Salinas (eds). Libro rojo de plantas de Colombia. Volumen 4. Es pecies maderables amenazadas: primera parte. Serie libros rojos de especies amenazadas de Colombia. Instituto Amazónico de Investigaciones Científicas SINCHI- Ministerio de Ambiente, Vivienda y Desarrollo Territorial. Bogotá.
Cavelier, J. D. Lizcano &M.T. Pulido. 2001. Colombia. pp: 443-496. En: Kappelle M. & A. D. Brown (eds. ). Bosques Nublados del Neotrópico. .Instituto Nacional de Biodiversidad (IN Bio). Santo Domingo de Heredia. Costa Rica.
Chaparro, O. 2009. Caracterización estructural de los bosques andinos de la vertiente oriental de la Cuchilla El Fara, Santander. Trabajo de grado en Ingeniería Forestal. Universidad Distrital Francisco José de Caldas. Bogotá. 90 p.
Condit, R. 1998. Tropical forest census plots: Methods and results from BCI, Panamá and a com parison with other plots. Springer. Berlin. 211 p.
Condit, R. S.P. Hubbell, J.V. Lafrankie, R. Sukumar, N.. Manokaran, R.B. Foster & P. S. Ashton. 1996. Species-area and species-individual relationship for tropical trees: a comparison of three 50-ha plots. Journal of Tropical cology 84: 549-562
Condit, R. E. LeighJr, S. LoodeLao &CTFS Working Group 2004. Species-area rela tionship and diversity mesaures in the forest dynamics plots. pp.: 79-81. En: Losos, E. & E. Leigh, (eds.). Tropical Forest Diversity and Dynamism. Findings from a Large-Scale Plot Network. University of Chicago.
Cortés-B, L. 2009. Caracterización de la vegeta ción de cuatro bosques de Roble ubicados en las veredas de Patios Altos y Canadá, Muni cipio de Encino, Santander. Trabajo de grado en Biología. Pontificia Universidad Javeriana. Facultad de Estudios Ambientales y Rurales. 128 p.
Duque, A.J. W. López, F.H. Moreno &J. Toro. 2008. Variabilidad estructural y ambiental en remanentes de bosques andinos. pp.: 17-33. En: León. , J. D. (ed. ). Ecología de bosques andinos. Experiencias de investigación. Universidad Nacional de Colombia. Medellín.
Etter, A. &W. Wyngaarden. 2000. Patterns of landscape transformation in Colombia, with emphasis in the Andean Region. Royal Sweedish Academy of Sciences. Ambio 29: 432-439.
Galindo-T, R. J. Betancur &J. Cadena-M. 2003. Estructura y composición florística de cuatro bosques andinos del Santuario de Flora y Fauna Guanentá-Alto Río Fonce, Cordillera Oriental colombiana. Caldasia 25: 313-325.
Galindo-T, R. J. Betancur &H. Mendoza. 2006. Estructura y composición florística de los bosques de roble en dos parques nacionales naturales en la Cordillera Oriental Colombiana, pp: 95-100. En: Solano C. & N. Vargas (eds. ). Memorias del I Simposio Internacional de Ro bles y Ecosistemas Asociados . . Fundación Natura - Pontificia Universidad Javeriana. Bogotá.
Galindo-T, C. &D.F. Correa-G. 2006. Caracterización fisonómica de un relicto de bosque de roble ubicado en la vereda El Ocaso, municipio de Bojacá, pp.: 95-100. En: Solano, C &N. Vargas. (eds.)Memorias del I Simposio Internacional de Robles y Ecosistemas Asocia dos. Fundación Natura - Pontificia Universidad Javeriana Bogotá
Gentry, A.H. 1982. Neotropical Floristic Diversity: phytogeographical connections between Central and South America, Pleistocene climatic fluctuations, or an accident of the Andean Orogeny? . Annals of the Missouri Botanical Garden 69: 557-593.
Gentry, A.H. 1995. Patterns of diversity and floristic composition in Neotropical Montane Forest, pp.: 103-126. En: Churchill, S.P., H. Balslev, E. Forero & J.L. Luteyn (eds.). Biodi versity and Conservation of Neotropical Montane Forests. Proceedings of the Neotropical Montane Forest Biodiversity and Conservation Symposium. Bronx. New York
González, A. 2009. Producción de frutos del roble Quercus humboldtii Bonpl., en dos bosques an dinos de la cordillera oriental. Trabajo de grado en Ingeniería Forestal. Universidad Distrital Francisco José de Caldas. Bogotá. 37p.
González, C. A. Jarvis &J.D. Palacio. 2006. Biogeography of the Colombian oak, Quercus humboldtii Bonpl. : geographical distribution and their climatic adaptation. International Centre for Tropical Agriculture (CIAT)/Museo de Historia Natural, Universidad del Cauca. Popayán. 10 p.
Hooghiemstra, H. &A.M. Cleef. 1993. Pleistocene climatic change and environmental and generic dynamics in the North Andean Montane Forest and Paramo, pp.: 35-49. En: Churchill, S.P., H. Balslev, E. Forero & J.L. Luteyn (eds.). Biodiversity and Conservation of Neo tropical Montane Forests. Proceedings of the Neotropical Montane Forest Biodiversity and Conservation Symposium. The New York Botanical Garden Bronx. New York
Hooghiemstra, H. T. Van der Hammen &A.M. Cleef. 2002. Paleoecología de la flora bosco sa. pp: 43-58. En: Guariguata E. & G. Kattan (eds. ). Ecología y conservación de bosques neotropicales. . . Editorial Tecnológica de Costa Rica. Costa Rica.
Ingeominas. 1998. Mapa geológico generalizado del Departamento de Santander. Bogotá.
Jiménez, A. 2009. Caracterización fisionómica y estructural de la vegetación natural en claros de roble, en la reserva biológica Cachalú, Municipio de Encino (Santander). Trabajo de gra do en Ingeniería Forestal. Universidad Distrital Francisco José de Caldas. Bogotá. 63 p.
Kappelle, M. 1992. Phytogeography of Talamanca montane Quercus forests. Journal of Biogeo graphy 19: 219-315.
Kappelle, M. 2006. Changes in diversity and structure along a succesional gradient in a Cos ta Rican Montane Oak Forest, pp.: 223-233. En: Kappelle, M. (ed.). Ecology and conser vation of Neotropical Montane Oak Forests. Ecological Studies, Vol. 185. Springer-Verlag BerlinHeidelberg
Kappelle, M. A. Cleef &A. Chaverri. 1989. Phytosociology of montane Chusquea-Quercus forests, Cordillera de Talamanca, Costa Rica. Brenesia 32: 73-105.
Kappelle, M. T. Geuze, M. Leal &A. Cleef. 1996. Succesional age and forest structure in a Costa Rica Upper Montane Quercus Forest. Journal of Tropical Ecology 12: 681-698.
Kessler, M. &J. Kluge. 2008. Diversity and endemism in tropical montane forests-from patterns to processes, pp.: 35-50. En: Gradstein, S.R., R.J. Homeier. & D. Gansert. (eds.). The Tropical Mountain Forest. Patterns and processes in a Biodiversity Hotspot. Göttingen Centre for Biodiversity and Ecology. Biodiversity and Ecology Series 2
López, W. &A. Duque. 2008. Diversidad alfa de especies arbóreas en remanentes de bosques montanos en la región norte de la Cordillera Central de Colombia, pp.: 35-56. En: León, J.D. (ed.). Ecología de bosques andinos. Experiencias de investigación. Universidad Nacio nal de Colombia Medellín
Losos, E. &CTFS Working Group. 2004. The structure of tropical forests, pp.: 69-78. En: Losos, E &E. Leigh. (eds.)Tropical Forest Diversity and Dynamism. Findings from a Large-Scale Plot Network. University of Chicago Chicago
Lozano, G. &H. Torres. 1965. Estudio fitosociológico de un bosque de Roble ( Quercus humboldtii Bonpl) de la Merced, Cundinamarca. Trabajo de grado Biología. Universidad Nacio nal de Colombia. Bogotá. 216 p.
Lozano, G. N. Ruíz, F. González &M.T. Murillo. 1996. Diversidad vegetal del Parque Nacional Natural Munchique, Cauca (Colombia), entre 1800 y 3050 msnm. pp.: 8-71. En: Pinto, P. (ed. ). Notas sobre biodiversidad. . Instituto de Ciencias Naturales-Museo de Historia Natural. Bogotá.
Magurran, A. 1989. Diversidad ecológica y su medición. Ediciones Vedra. Barcelona. 200 p.
Marín-C, C. &J. Betancur. 1997. Estudio florístico en un robledal del Santuario de Fauna y Flora de Iguaque (Boyacá, Colombia). Revista de la Academia Colombiana de Ciencias 21: 249-259.
Mendoza-Cifuentes, H. &B. Ramírez-Padilla. 2000. Flora de La Planada: Guía Ilustrada de Familias y Géneros. Instituto de Investigación de Recursos Biológicos Alexander von Hum boldt, Fundación para la Educación Superior, World Wildlife Fund. Bogotá.
Ocaña, Y. 2005. Caracterización florística y estructural de unidades de bosque andino en las veredas Minas y Patios Altos del municipio de Encino, Santander. Colombia Forestal 9: 70-86.
Pan-Amazonian. 2009. Project for the advancement of networked science in Amazonian, Protocols for field measurements at Caxiunana, Brasil and Tambopata, Perú (Preliminary version), 14 p. Disponible en: http://www.eci.ox.ac.uk./panamazonia/rainfor_field_manual_ spanishpdf
Paredes, A. &A. Martínez. 2005. Diversidad y caracterización florística y estructural de la ve getación secundaria de la reserva biológica Cachalú, con propósitos de restauración. (Encino-Santander). Trabajo de grado. Ingeniería Forestal. Facultad del Medio Ambiente y Recur sos Naturales. Universidad Distrital Francisco José de Caldas. Bogotá. 238 p.
Phillips, O. &J. Miller. 2002. Global patterns of plant diversity: Alwyn H. Gentry's forest transect data set. Missouri Botanical Garden Press. St. Louis. 186 p.
Pulido, M.T. J. Cavelier &P. Cortés. 2006. Structure and composition of Colombian Mon tane Oak Forests. pp: 141-151. En: Kappelle, M. (ed. ). Ecology and Conservation of Neotro pical Montane Oak Forests. Ecological studies 185. . . Springer-Verlag. Berlin Heidelberg.
Rangel-Ch, O. &A. Garzón. 2000. Aspectos de la estructura, de la diversidad y de la dinámica de la vegetación del Parque Regional Natural Ucumarí: Instituto de Ciencias Naturales. Uni versidad Nacional de Colombia. Bogotá.
Rangel-Ch, O. P.D. Lowry-C, M. Aguilar-P &A. Garzón-C. 1997. Tipos de vegetación en Co lombia. Una aproximación al conocimiento de la terminología fitosociológica, fitoecológica y de uso común. pp. : 59-87. En: Rangel-Ch. , O. , P. D.Lowry-C, M. Aguilar-P. (eds.). Colombia Diversidad Biótica II. Tipos de vegetación en Colombia. Instituto de Ciencias Naturales. Universidad Nacional de Colombia. Bogotá.
Rangel-Ch, O. &G. Lozano. 1986. Un perfil de vegetación entre la Plata (Huila y el Volcán Pu racé). Caldasia 14: 503-547.
Rangel-Ch, O. &A. Velásquez. 1997. Métodos de estudio de la vegetación, pp.: 59-88. En: Rangel- Ch, O., P.D. Lowry-C, M. Aguilar-P. (eds.). Colombia Diversidad Biótica II. Tipos de vegeta ción en Colombia. Instituto de Ciencias Naturales. Universidad Nacional de Colombia Bogotá
Rincón, A. &N. Bernal. 2007. Factores antrópicos asociados e interrelaciones con el estado de los ecosistemas andinos, pp.: 10-128. En: Armenteras, D &N. Rodríguez. (eds.)Moni toreo de los ecosistemas andinos 1995-2005: Síntesis y perspectivas. Instituto de Investiga ción de Recursos Biológicos Alexander von Humboldt Bogotá
Samper, K.C. &M.I. Vallejo. 2007. Estructura y dinámica de poblaciones de plantas en un bos que andino. Revista de la Academia Colombiana de Ciencias 31: 57-68.
Solano, C. 2006. Reserva Biológica Cachalú: 10 años de investigación en bosques de roble, pp.: 11-23. En: Solano, C &N. Vargas. (eds.)Memorias del I Simposio Internacional de Roble y Ecosistemas Asociados. Fundación Natura - Pontificia Universidad Javeriana Bogotá
Vallejo-Joyas, M.I. A. Londoño-Vega, R. López-Camacho, G.. Galeano, D. Álvarez-E &D-W. Álvarez. 2005. Establecimiento de parcelas permanentes en bosques de Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá. 310 p.
Vallejo, M.I. &G. Galeano. 2009. Cambios a corto plazo en los patrones de distribución es pacial en nueve especies de plantas comunes en un bosque nublado al sur-occidente de Co lombia. Caldasia 31: 76-98.
Van der Hammen, T. &O. Rangel-Ch. 1997. El estudio de la vegetación en Colombia, pp.: 17-57. En: Rangel-Ch, O., P.D. Lowry-C, M. Aguilar-P. (eds.). Colombia Diversidad Biótica II. Tipos de vegetación en Colombia. Instituto de Ciencias Naturales. Universidad Nacional de Colombia Bogotá
Van der Hammen, T. R. Jaramillo &M.T. Murillo. 2008. Oak Forests of the Andean Forest zone of the Eastern Cordillera of the Colombian Andes. pp: 595-614. En: Stuttgart, J. (ed. ). Studies in Tropical Andean Ecosystems 7. Universidad de Amsterdam. Amsterdam.
Villarreal, H. M. Álvarez, S. Córdoba, F. Escobar, G. Fagua, F. Gast, H. Mendoza, M. Ospina & A. M. Umaña. 2006. Manual de métodos para el desarrollo de inventarios de biodiversidad. Programa de Inventarios de Biodiversidad. Instituto de Investigación de Recursos Biológi cos Alexander von Humboldt. Bogotá. 236 p
Zernig, K. &J. Betancur. 1994. Flora de Iguaque, pp. 93. En: Cavelier, J &A. Uribe. (eds.)Resúmenes del Simposio Nacional "Diversidad Biológica, Conservación y Manejo de los Ecosistemas de Montaña de Colombia. Universidad de los Andes Bogotá




License
Colombia Forestal retains the patrimonial rights (copyright) of the published works, and favors and allows the reuse of the same under the Creative Commons Attribution-ShareAlike 4.0 International license, so they can be copied, used, disseminated, transmitted and exhibited publicly, provided that:
You acknowledge the credits of the work in the manner specified by the author or licensor (but not in a way that suggests that you have their support or that they endorse your use of their work).