
DOI:
https://doi.org/10.14483/2256201X.3209Publicado:
01-01-2001Número:
Vol. 7 Núm. 14 (2001)Sección:
Artículos de investigación científica y tecnológicaDinámica temporal y espacial de la fragmentación del bosque nativo en el sur de Chile
Palabras clave:
Fragmentación, Indice de diversidad de Patton, Dimensión fragtal, Factor forma, Osorno, Chile (es).Descargas
Referencias
Burrough P. 1981. Fractal dimensions of landscapes and other envrionmental data. Nature Vol 294 :240-242.
Corporación Nacional Forestal, CONAF & Comisión Nacional del Medio Ambiente CONAMA- 1997. Catastro y evaluación de los recursos vegetacionales de Chile. Base de datos SIG- Xa Región.
Donoso C. 1981. Tipos forestales de los bosques nativos de Chile. Corporación Nacional Forestal y Organización de las Naciones Unidas para la Agricultura y la Alimentación, Documento de trabajo N° 38.
Donoso C. & Lara A. 1997. Utilización de los bosques nativos en Chile: pasado, presente y futuro. En: Ecología de los bosques nativos de Chile. Armesto J., C. Villagrán & M.K. Arroyo (Eds). Editorial Universitaria S.A. Santiago, Chile. 477.
Fallas J. 1998. Sistemas integrados de Información geográfica. Laboratorio de teledección y sistemas de información geográfica. Universidad Nacional. Heredia, Costa kica.
Forman R T.T. 1995. Land Mosaics, the ecology of landscapes and regions. Cambridge University Press, Great Britain.
Forman R.T.T. & M. Godron. 1986. Landscape ecology. John Wiley and Sons.
Fuentes Q.E. 1994. ?Qué futuro tienen nuestros bosques¿. Hacia la gestión sustentable del paisaje del centro y sur de Chile. Ediciones Universidad Católica de Chile. Santiago, Chile.289 p.
Gantz A & J. Rau. 1999. Relación entre el tamaño mínimo de fragmentos boscosos y su riqueza de especies de aves en el sur de Chile. Anales del Museo de Historia Natural de Valparíso 24:85-90.
Harris L.D. & G. Silva-López. 1992. Forest fragmentation and the conservation of biological diversity. In: Conservation biology: the theory and practice of nature conservation and management. Fiedler P.L & 5.1K. Jain, Editors. Chapman & Hall. New York, U.S.A 507 p.
Harris L.D. 1984. The fragmented forest. The University of Chicago Press. 200 p. Chicago, U.S.A.
Helzer Ch & D. E. Jelinski. 1999. The relative importance of patch arca and perimeter-area ratio to grassland breeding birds. Ecological Applications 9(4):1448-1458.
Henao S. 1988. Introducción al manejo de cuencas hidrográficas. Universidad de Santo Tomas. Centro de Enseñanza Desescolarizada. Ediciones Usta. Bogotá, Colombia. 395 p.
Instituto Nacional de Estadísitcas INE. 1997. VI Censo Nacional Agropecuario, Resultados preliminares 1997. INE. 443 pp.
Krummel J.R.; R.H Gardner; G. Sugihara; R.V O'Neill & P.R. Coleman. 1987. Landscape patterns in a disturbed environment. Oikos 48 : 321 - 324.
Laurance WF. & R.O. Bierregaard. 1997. Tropical Forest Remnants Ecology, Management and Conservation of Fragmented Communities (Abstracts on une www.press.uchicago.edu/ Images/Chicago/rbullet.gif).
Laurance W.F & E. Yensen. 1991. Predicting impacts of edge effects in fragmented habitats. Biological Conservation 55:77-92.
Laurance WE; L.V Ferreira; J.M Rankin-de Merona; S.G. Laurance; R.W. Hutchings & T.E. Lovejoy. 1998. Effects of forest fragmentation on recruitment patterns in amazonian tree communities. Conservation Biology 12(2):460-464.
Lord J.M. & D.A. Norton. 1990. Scale and the spatial concept of fragmentations. Conservation Biology 4:197-202.
Lovejoy TE. 1982. Area-perimeter relation for rain and cloud arcas. Science Vol 216:185 :187.
Lovejoy T.E.; R.0 Bierregaard; A.B. Rylands; J.R. Malcom; K.S. Brown; C.E. Quintela; L.H. Harper; A.H. Powell; G.V. PowelL; H. Shubart & M.B. Hays. 1986. Edge and other effects of isolation in amazon forest fragments. In: Conservation Biology : the science of scarcity and diversity. Soulé M.E (Ed.). Sinauer Associates. Sunderland, Massachussets, U.S.A 584 p.
MacArthur R.H. & E.O. Wilson. 1967. The theory of island biogeography. Monographs in Population Biology N° 1. Princeton University Press. New Jersey. 215 p.
Mansilla, E. 1995. Auditoría ambiental a las actividades humanas de la cuenca del Río Damas. Seminario de título para optar al título de Técnico Universitario en Control Ambiental. Universidad de Los Lagos. Sede Puerto Montt. Puerto Mont, Chile.
McGarigal, K & B. Marks. 1995. Fragstat's, spatial patterns analysis program for quantifying lansdscape structure. Portland OR: U.S. Department of Agrigulture, Forest Service, Pacific Northwest, Research Station. 122 13.
Meffe G.K. & C.R. Carroll. 1994. Principies of conservation biology. Sinauer Associates, Inc. Sunderland, Massachusetts. 600 p.
Moreno, L.A. 1994. Estudio hidroambiental del Río Damas (X Región): Análisis de la situación actual. Tesis de grado, Facultad de Ingeniería. Departamento Ingeniería Civil. Universidad de concepción. 182 pp.
Myers N. 1994. Global biodiversity II : losses and threats. In: Principies of Conservation Biology. Meefe G.K. & C.R. Carroll. (eds). Sinauer Associates Pulishers. Sunderland, Massachusetts, U.S.A 600 p.
Noss R.F. & B Csuti. 1994. Habitat fragmentation. In: Principies of Conservation Biology. Mefee G.K. & Carroll (eds).).Sinauer Associates Publishers. Sunderland Massachusetts, U.S.A. 600 p.
Patton D.R. 1975. A diversity index for quantifying habitat edge. Wildlife Society Bulletin 3: 171 - 173:
Pimm L.S. 1998. The forest fragment classic. Nature 393: 23-24.
Preston F.W. 1962. The canonical distribution of communnes and rarity. Ecology 43 :185_ 215, 410-432.
Ripple J.; G.A Bradshaw & TA Spies. 1991. Measuring forest landscapes patterns in the Cascade Range of Oregon, USA. Biological Conservation 57: 73-88
Shafer C.L. 1990. Island theory and conservation practice. Smithsoniam Institution Press. Washington and London. 189 p.
Sugihara G. & May R. 1990. Applications of fragtal in ecology. Trends in Ecology and Evolution. Vol 5, N° 3: 79: 86.
Turner M.G. 1989. Landscape ecology: the effect of pattern on process. Annu. Rey. Ecol. Syst. 20: 171-197.
Willson, E.O. & Willis E.O. 1975. Applied biogeography . In: ,Cody, M.L. & Diamond J.M. (Eds). Ecology and the evolution of communities. Harvard Univ. Press. Cambridge, -M.A pp 522-534.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Artículos científicos
Colombia Forestal, 2001-12-00 vol:7 nro:14 pág:71-84
Dinámica temporal y espacial de la fragmentación del bosque nativo en el sur de Chile
Leyla M. Montenegro Calderón
Universidad Distrital Francisco José de Caldas
Palabras claves:
Fragmentación, Indice de diversidad de Patton, Dimensión fragtal, Factor forma, Osorno Chile.
INTRODUCCIÓN
Las actividades humanas sobre el planeta tales como agricultura, silvicultura, ganadería y establecimiento de asentamientos humanos, han determinado que la mayoría de los paisajes contemporáneos presenten algún grado de fragmentación (Myers, 1994; Krummel et al., 1987). Los bosques nativos se cuentan entre los ecosistemas con mayor grado de fragmentación a nivel mundial. La fragmentación ha sido definida como la transformación de un bosque originalmente continuo, en unidades más pequeñas, rodeadas por matrices, generalmente antrópicas, que les son hostiles (Lord & Norton, 1990; Shafer, 1990; Harris & Silva-López, 1992; Noss & Csuti, 1994; Myers, 1994; Pimm, 1998). Estos fragmentos se comportan como "islas virtuales" inmersos en un océano antrópico y frecuentemente son analizados en el contexto de la teoría del aislamiento biogeográfíco propuesta por MacArthur & Wilson (1967).
El resultado de la fragmentación es un paisaje en el cual se mezclan áreas manejadas y transformadas por el hombre con fragmentos de vegetación nativa, es decir, parches de coberturas de diferentes tamaños y formas. La relación entre la forma y el tamaño de los fragmentos influye en. un número importante de procesos ecológicos (Laurance & Yensen, 1991; Harris & Silva-López, 1992; Noss & Csuti, 1994) como son la proliferación de especies heliófitas a lo largo de las márgenes de los fragmentos (Lovejoy et al., 1986), el incremento en la mortalidad de árboles y plantas juveniles, lo cual afecta diferencialmente la regeneración de las especies (Harris, 1984), alteración del microclima afectando con ello la temperatura y humedad al interior del fragmento (Laurance,,1997) y procesos de especiación y extinción de especies (Krummel et al., 1987) entre otros.
Un paisaje fragmentado puede describirse por atributos tales como: número, frecuencia, tamaño, forma, grado de aislamiento, densidad, conectividad y tipo de matriz que rodea a los fragmentos (Harris, 1984; Ripple et al., 1991; Harris & Silva-López, 1992; MacGarigal & Marks, 1995). Se ha demostrado que estas propiedades influyen directa o indirectamente sobre la fauna y la flora (Harris, 1984; Laurance et al., 1998; Gantz & Rau, 1999) debido a que algunas especies son más vulnerables que otras ante una reducción del área, incremento del aislamiento entre fragmentos, efectos de borde y otros factores que acompañan el proceso de fragmentación (Noss & Csuti, 1994).
Investigaciones ecológicas hansugerido que la forma de .los fragmentos de vegetación (Vgr islas virtuales) es un parámetro importante que debe ser estudiado, debido al "efecto península" que aumentaría el "efecto de borde" de fragmentos con formas elongadas y peninsulares, comparativamente con formas circulares; la forma también ha incidido en el diseño de áreas silvestres protegidas (MacArthur & Wilson, 1967; Wilson & Willis, 1975; Forman & Godron, 1986; Forman, 1995).
Usualmente la forma de los fragmentos se analiza como la relación área-perímetro, la cual permite inferir un incremento en la complejidad de la forma de éste, en consecuencia, si el fragmento se aleja del modelo circular ideal estará más expuesto a efectos ecológicos de la fragmentación, peninsularización y efectos de borde (Harris, 1984; Forman & Godron, 1986).
Por otra parte, el análisis mediante registros aerofotográficos, cartografía temática o imágenes provenientes de sensores remotos, se ha utilizado con el fin de comparar cambios de cobertura vegetal a través del tiempo (Ripple et al., 1991; Fallas, 1998), pero son pocas las medidas precisas relativas a estos cambios, menos aún. las que tienen que ver con la variación en la forma de los parches de los paisajes fragmentados. En tal sentido resulta importante comprender cómo el tamaño y la forma geométrica de los fragmentos de cobertura, se relacionan con procesos naturales o antrópicos a través de análisis espaciales y temporales (Wilson & Willis, 1975).
Esta investigación tuvo como objetivo cuantificar el grado de fragmentación a través del tiempo en remanentes de bosque nativo, a nivel de una escala espacial correspondiente a la cuenca hidrográfica. El estudio se realizó en la cuenca hidrográfica del Río Damas, Provincia de Osorno, sur de Chile. Con base en cartografía temática se analizó la dinámica de fragmentación del bosque nativo comparando los fragmentos de 1966 (N=165) con los de 1996 (N=121). Se emplearon tres estadígrafos espaciales para cuantificar la ecología de paisajes fragmentados: área, perímetro y forma de los fragmentos para ambos períodos comparados. Para cuantificar la forma se utilizó el Indice del Factor Forma, el Indice de Diversidad de Patton y la Dimensión Fractal.
Se encontró que en tres décadas, el número de fragmentos aislados disminuyó en un 30% y el :área total ocupada por éstos en un 6%. En. 1966 existía una mayor frecuencia de fragmentos pequeños (< 20 ha) de formas simples (ovales y oblongos), comparativamente los de 1996 fueron amorfos. Esto último se verificó también por un aumento de la dimensión fractal. Se presentan sugerencias para el manejo de paisajes fragmentados.
PLANTEAMIENTO DEL PROBLEMA
La fragmentación de los bosques nativos es un proceso que ha congregado una gran atención de la comunidad científica debido a sus implicaciones para la conservación de la biodiversidad (Myers, 1994; Laurance & Yensen, 1991; Laurance & Birregard, 1997). Estudios recientes en Costa Rica (Meffe & Carroll, 1994) y en la Amazonia (Lovejoy et al., 1986; Laurance et al., 1998) evidenciaron que los bosques tropicales se han reducido y fragmentado dramáticamente en los últimos 50 años, producto del incremento de la actividad agropecuaria.
Este mismo patrón se observa en el sur de Chile, donde los bosques nativos han sido talados y fragmentados debido a la necesidad de habilitar tierras para pastoreo y cultivos, y en algunos casos fueron sustituidos por plantaciones de especies exóticas (Donoso, 1981; Fuentes, 1994; Donoso & Lara, 1997).
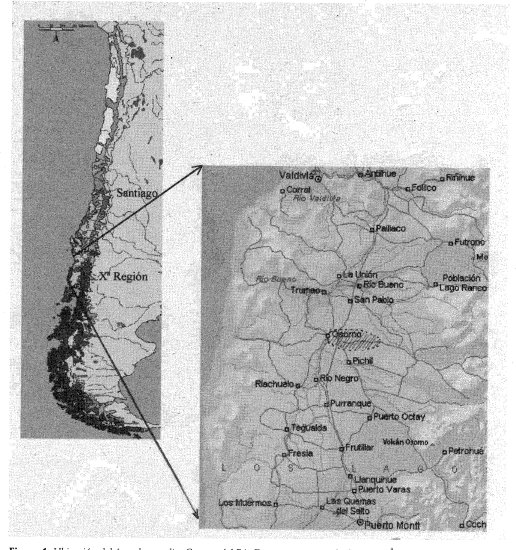
En la cuenca del Río Damas, localizada en la Provincia de Osorno, Décima Región de Los Lagos, Chile (Figura 1), la vegetación nativa boscosa se presenta irregularmente distribuida y fuertemente fragmentada. La forma de los fragmentos y su dinámica de fragmentación a través del tiempo no se han evaluado.
OBJETIVO GENERAL
Evaluar el grado de fragmentación de los remanentes de vegetación nativa en la cuenca hidrográfica del Río _Damas, a través, del tiempo.
OBJETIVOS ESPECÍFICOS
Cuantificar el grado de fragmentación del paisaje de la cuenca del Río Damas, a través del cálculo de la forma y el tamaño de los fragmentos de vegetación nativa y compararla a través del tiempo.
Comparar la habilidad de tres índices en el cálculo de la forma de los fragmentos para detectar modelos espaciales en paisajes fragmentados.
HIPOTESIS
Ho: El tamaño de los fragmentos de vegetación nativa en la cuenca del río Damas no varía significativamente a través del tiempo.
Ho: La forma de los fragmentos no varía significativamente a través del tiempo en la cuenca del Río Damas.
AREA DE ESTUDIO
La investigación se realizó en la cuenca hidrográfica del Río Damas, Provincia de Osorno, Décima Región de Los Lagos, Chile, la cual se sitúa entre los 40 0 33 y los 40 0 45' de latitud sur, y entre los 72° 36' y 73° 09' de longitud oeste. Involucra un área de 513.7 km' y presenta una longitud máxima de 50 km. La diferencia de altura entre el nacimiento y la desembocadura es de 325 m. la topografía en la parte alta está caracterizada por lomajes suaves y ondulados y en la parte baja por amplias planicies (Moreno, 1994; Mansilla, 1995). El uso del suelo es principalmente agropecuario (80%), sus laderas con pendientes muy débiles, de menos de 6 grados, facilitan la ocupación humana y el uso intensivo de sus recursos naturales. Más del 50% de la superficie esta dedicada al cultivo de forrajeras; la tenencia de la tierra está en manos particulares (INE 1997).
METODOLOGIA
Para la investigación se consideró el paisaje de la cuenca del Río Damas de los arios 1966 y 1996, es decir con una diferencia de tres décadas. Los datos de 1966 fueron obtenidos con base en la cartografía del Instituto Geográfico Militar (IGM) a escala 1:50.000. Los datos de 1996 fueron obtenidos de la cartografía del Catastro del Bosque Nativo, elaborado por la CONAF 4 CONAMA5 (1997) a escala 1:50.000.
En primer lugar se delimitó la cuenca hidrográfica con base en la metodología de la línea divisoria de aguas (Henao, 1988), posteriormente se clasificó el tipo de cobertura distinguiendo principalmente tres tipos de unidades: a) urbano, b) agropecuario, el cual involucró praderas y cultivos, y c) vegetación nativa ("fragmentos"), siendo este último tipo de cobertura el objeto de estudio. La dinámica y el grado de fragmentación del paisaje fue evaluada como una función de la variación a través del tiempo (1966 y 1996) del tamaño y la forma de los fragmentos.
El tamaño de los fragmentos se expresó a través del área, la cual se calculó utilizando un planímetro digital marca Placom KP-80N, con resolución de 0,1 cm' y precisión de ± 0,2%. El perímetro de obtuvo empleando un curvímetro digital marca Mapmeter Cv-9, con resolución de 1 mm. El tamaño por ario se expresó como el promedio del área de todos los fragmentos en dicho período.
La forma de los fragmentos se describió a través de tres índices: a) el Indice de Diversidad de Patton, (Di), b) el Factor Forma (Ff), y c) la Dimensión Fractal (D). Los primeros dos se fundamentan en la geometría euclidiana (líneas "lisas",cuadrados, círculos, esferas, cubos), el tercero en la geometría fractal (líneas "aserradas", formas irregulares) (Mandelbrot, 1977, citado por Sugihara & May, 1990).
El Indice de Diversidad de Patton (1975) expresa la forma de los fragmentos como la relación:

es el área (m2) y Di es el índice de diversidad de Patton de cada fragmento. Di varía entre 1 y 2, cuando Di es 1, la forma del fragmento semeja un círculo, a medida que éste va aumentando la forma se va tornando más compleja. Para cada período analizado, el Di fue agrupado en cinco (5) rangos de acuerdo con Henao (1988), quien. los clasifica en: redondo (Di < 1,25), oval redondo (1,25 > Di < 1,5), oval oblongo (1,5 >Di < 1,7), rectangular oblongo (1,7 > Di < 2) y amorfo (Di > 2).
El Di promedio para cada ario (± error estándar), se obtuvo como el valor medio de todos los Di por período. La prueba estadística no paramétrica de Kolmogorov-Smirnov para dos muestras y la prueba de bondad de ajuste de x2 para tablas de contingencia se emplearon para comparar el Di de los fragmentos entre arios y entre categorías de clasificación, respectivamente, a un nivel de significación a = 0,05. En adelante, a menos que se indique lo contrario, el nivel de significancia para todas las pruebas estadísticas será de α= 0,05.
La Dimensión Fractal: D. Es una medida cuantitativa de complejidad paisajística (Turner, 1989), es invariante de escala y estadísticamente robusta (cf. Ripple et al., 1991), es considerada el descriptor más ad hoc para cuantificar la fragmentación de diferentes paisajes (Sugihara May, 1990; McGarigal & Marks, 1995).
La dimensión fractal, D, para el conjunto total de fragmentos en cada ario, se obtuvo siguiendo el método de cálculo propuesto por Love joy (1982): P., -VAD , esto es: logP 1/2D log A, donde P es el perímetro, (m), log transformado, A es el área (m2), log transformada y D es la pendiente de la regresión. La dimensión fractal usualmente es aplicada a grandes paisajes mediante el uso de la relación área-perímetro; si existen suficientes datos (n >20), la dimensión fractal, D, es dos (2) dividido por la pendiente de la regresión linear obtenida de logP sobre logA. La D obtenida de esta manera ni es igual a la pendiente (Krummel et al.,1987) ni es dos veces la pendiente, como lo informan algunos autores (McGarigal & Marks, 1995). La dimensión fractal de cada fragmento individual discreto, se'obtuvo operando la expresión anterior por su equivalente:
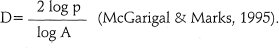
La dimensión fractal expresa el grado de complejidad de los fragmentos, así, en ambas fórmulas (i. e., para el total y para cada fragmento), D oscila entre 1 (formas euclidianas simples) y 2 (formas complejas, amorfas y elongadas) (Krummel et al., 1987, Sugihara & May, 1990; Ripple et al., 1991; McGarigal & Marks, 1995).
La prueba estadística no paramétrica de Wilcoxon-Mann-Whitney para dos -muestras independientes y la prueba de bondad de ajuste de X2 para tablas de contingencia, se emplearon para comparar la dimensión fractal de los fragmentos individuales entre arios y entre categorías de clasificación, respectivamente.
El Factor forma (FF) relaciona el perímetro de un círculo de igual área que el fragmento vs. el perímetro del fragmento, mediante la ecuación: FF = pc / p (Forman, 1995), donde: pc es el perímetro de un círculo de igual área que el fragmento, p es el perímetro del fragmento y FF es el factor forma. El factor forma varía entre O (formas lejanas de la circularidad) y 1 (formas circulares). El valor promedio por ario - se obtuvo como el valor medio de todos los FF por período. La prueba estadística de Wilcoxon-Mann-Whitney para dos muestras y la prueba X para tablas de contingencia se emplearon para comparar el FF de cada uno de los fragmentos entre arios y entre categorías de clasificación, respectivamente.
RESULTADOS
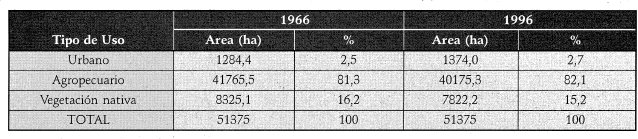
La estimación del cambio en_la cobertura de la vegetación nativa a través del tiempo en la cuénca del Río Damas demostró que el paisaje ha sido modelado por regímenes de perturbación antrópica, los cuales están generando un paisaje ho : mogéneo al interior de ésta. La Tabla 1 muestra que la cobertura de vegetación nativa disminuyó del 16,2% (1966) al 15,2% (1996) y que la cobertura predominante es de tipo agropecuaria (80%).
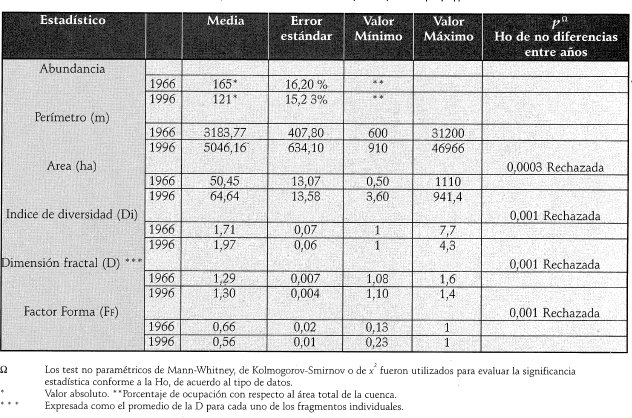
El grado de fragmentación, como una medida de las estadísticas del área, perímetro y forma de los fragmentos al interior de la cuenca del Río Damas, varió significativamente entre 1966 y 1996 (Tabla 2).
En total se contabilizaron 165 fragmentos en. el ario 1966 y 121 en el ario 1996. De los fragmentos registrádos en 1966 sólo 109 (66%) fueron contabilizados en 1996, registrando una pérdida del 34% (56) de fragmentos, en tanto que en 1996 el 90% (109) de éstos se contabilizaron en el paisaje inicial y el 10% (12) se registraron como nuevos.
La dinámica temporal (i.e., 1966 a 1996) de la fragmentación, reveló que el proceso al interior de la cuenca no aumentó, es decir, no se registró un mayor número de fragmentos en el intervalo de tiempo analizado. Así mismo, la desaparición de 56 fragmentos sugiere que existe un impacto antrópico diferenciado sobre la vegetación nativa, pues los fragmentos que desaparecieron se localizaban en la cuenca baja del río (cerca de la ciudad de Osorno), que coincide con los mejores tipos de suelos, demostrando con ello que el proceso de deforestación en la cuenca no es aleatorio.
El valor medio del área de los fragmentos registró un aparente incremento entre 1966 y 1996, pues varió de 50,5 a 64,6 ha, respectivamente; sin embargo el área total ocupada por los remanentes de vegetación nativa disminuyó significativamente (P=0,0003), lo cual podría deberse a que 1966 presentó un mayor porcentaje de fragmentos de área pequeña (< 20 ha.) y a la desaparición de éstos en 1996. En términos de área, esta disminución representó una reducción de 502,9 ha., (6%) lo cual podría sugerir una tasa de deforestación de 16,8 ha/ ario ( =1,8 fragmentos/año).
El valor medio del área de los fragmentos registró un aparente incremento entre 1966 y 1996, pues varió de 50,5 a 64,6 ha, respectivamente; sin embargo el área total ocupada por los remanentes de vegetación nativa disminuyó significativamente (P=0,0003), lo cual podría deberse a que 1966 presentó un mayor porcentaje de fragmentos de área pequeña (< 20 ha.) y a la desaparición de éstos en 1996. En términos de área, esta disminución representó una reducción de 502,9 ha., (6%) lo cual podría sugerir una tasa de deforestación de 16,8 ha/año (=1,8 fragmentos/año).
La distribución por intervalos de clase del tamaño de los fragmentos, mostró que 1966 presentó una mayor abundancia (61,8 %) de fragmentos pequeños (< 20 ha.) que 1996 (49,6%). Esta situación se invierte en los sucesivos rangos de área, pues 1996 presentó mayores frecuencias de fragmentos de áreas grandes que 1966. En otras palabras, en el paisaje de 1966 los fragmentos variaron en tamaños, fue un paisaje más hetereogéneo; con el paso del tiempo la dinámica de fragmentación modeló un paisaje homogéneo (1996).
El índice de diversidad de Patton, la dimensión fractal y el factor forma reflejaron una diferencia estadísticamente significativa en el incremento de la complejidad de la forma de los fragmentos a través del tiempo (Tabla 2).
El índice de diversidad de Patton (Tabla 3) demostró que en promedió la forma de los fragmentos del ario 1966 fue oval-oblonga (Di =1,7 _± 0.07) en tanto que el año 1996 fue amorfa (Di = 1,97 -1-.0.06) (Tabla 2) . La prueba de Wilcoxon-Mann-Whitney demostró que esta diferencia fue estadísticamente significativa (P<0,001). El análisis de x2 permite concluir que existen diferencias significativas (P < 0,001) en la distribución por rangos de forma, entre 1966 y 1996. La prueba de Kolmogorov-Smirnov permitió rechazar la hipótesis y concluir que la forma de los fragmentos varió significativamente entre 1966 y 1996 (P=0,001).
La Dimensión fractal, D, (Tabla 4) muestra que el mayor porcentaje de fragmentos en 1966 se localiza en el rango < 1,24 (22 %), y en 1996 en el rango > 1,35 (>61,2 %), lo cual indica que la forma de los fragmentos del año 1966 fue menos compleja que la del año 1996. El análisis x2 permite concluir que existen diferencias significativas (P< 0,001) en la distribución de fragmentos por rangos de D entre años (i.e., 1966 y 1996).
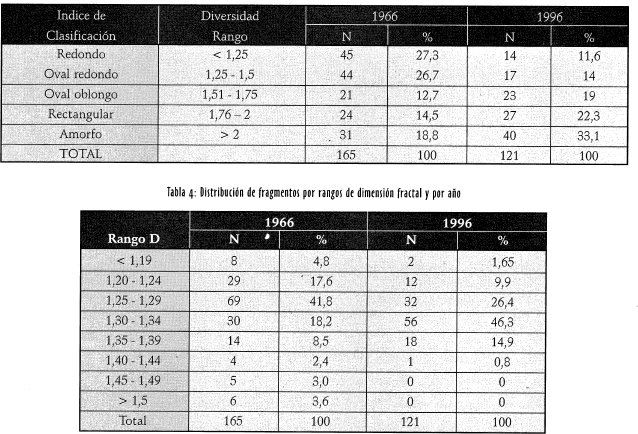
El análisis de la dimensión fractal para el conjunto de fragmentos por ario, calculada a través de la regresión logarítmica, mostró una relación positiva entre el área (m ) y el perímetro (m) de los fragmentos, con las funciones: para 1966: log A= 0,53+ 1,48logP; con F=550,92, g1=164; donde la pendiente fue significativamente distinta de cero (P=0,0001) y la regresión explicó un 77,16% de la varianza (r2 =0, 77). Para este año el valor de la dimensión fractal (D) fue de 1,35 (2 /1,48) (McGarigal & Marks, 1995), (Figura 2). Para 1996: logA= 1,603 + /1,336 logP; con F=881,04, g1=120; donde la pendiente también fue significativamente - distinta de cero (P.---0,0001)2 y la regresión explicó un 88% de la varianza (r =0,88). Para este ario el valor de D fue de 1,49 (Figura 3).
Los valores de la dimensión fractal obtenidos a través de los modelos de regresión logarítmica para 1966 y 1996 (1,35 y 1,50 respectivamente) evidenciaron que la forma de los fragmentos del ario 1996 es más compleja (i.e., amorfa) que los del año 1966. La prueba de Wilcoxon-Mann-Whitne permite concluir que la forma de los fragmentos varió significativamente entre un ario y otro (P=0,001), por tanto se rechaza la hipótesis de nulidad.
El Factor forma (Tabla 5) muestra que el 62% de los fragmentos del ario 1966 fueron circulares (FF »1), en tanto que en 1996 el 60% de éstos se alejaron de dicha forma (FF <0,5), lo cual concuerda con los resultados del índice de Patton (1975) y de la dimensión fractal. El valor promedio del FF (Tabla 2) muestra que en 1966 los fragmentos fueron circulares (FF=0,66 ±0.02) en tanto que 1996 se alejó de la forma circular (FF=0,56 ±0.01).La prueba de Wilcoxon-Mann-Whitney permite concluir que la forma de los fragmentos varió significativamente (P=0,001) entre un año y otro, rechazando la hipótesis nula.
DISCUSIÓN
Un examen de los resultados obtenidos revela la habilidad de los índices para captar las características de la forma de los fragmentos, las cuales tienen implicaciones importantes para el manejo de paisajes fragmentados.
Los índices de forma evaluados (i.e., índice de diversidad, dimensión fractal y factor forma) demostraron que existen diferencias estadísticamente significativas en la forma de los fragmentos a través del tiempo. En general, los índices revelaron que la forma de los fragmentos en 1966 fue menos compleja o más cercana a la forma circular ideal; en contraste, 1996 se caracterizó por fragmentos de formas amorfas y elongadas.
Un factor que podría estar contribuyendo al incremento en la complejidad de la forma de los fragmentos del paisaje del ario 1996, sería atribuido a una mayor presión antrópica sobre el borde de los fragmentos, o a la influencia de la matriz que los rodea, lo cual se reflejó en un incremento del perímetro y de la forma fractal de éstos a través del tiempo
Fragmentos elongados y amorfos son más susceptibles a los procesos que se generan por el "efecto de borde", la pérdida de especies y la acción de la matríz circundante. El incremento en el perímetro de los fragmentos beneficia a algunas especies, pero perjudica a otras; esto indica que especies de condiciones ecológicas del interior de los bosques podrían estar siendo perjudicadas y en consecuencia se estaría favoreciendo especies heliófitas o exóticas que competirían con las especies nativas.
Por otra parte el proceso natural de sucesión ecológica que podría generarse en el perímetro de los fragmentos depende fundamentalmente de la matriz circundante, de la conectividad entre fragmentos, del grado de aislamiento entre éstos y de la presencia de un fragmento "fuente" que aporte el material (semillas) necesario para que el proceso se genere; en tal sentido se hace necesario evaluar el efecto de la matriz, la conectividad entre fragmentos y la riqueza de especies de éstos, a fin de proponer acciones de manejo acordes con la dinámica de fragmentación de la región.
La dimensión fractal y el índice de diversidad de Patton fueron medidas robustas de la forma promedio de los fragmentos, lo cual coincidió con los resultados obtenidos por Ripple et al., (1991), en contraste con el factor forma.
El índice de diversidad de Patton fue consistente con la dimensión fractal y en general fue un buen estimador para cuantificar diferencias aparentes, y de alguna forma evidentes en la forma de los fragmentos, en tanto que la dimensión fractal reveló diferencias finas. y sutiles, coincidiendo con MacGarigal & Marks (1995). En tal sentido es válido aceptar que la dimensión fractal es poco afectada por la escala, comparativamente con el índice de diversidad de Patton, consecuentemente se puede considerar a la dimensión fractal como un estimador estadísticamente robusto cuando se quieren comparar paisajes que difieren en la resolución de sus escalas espaciales y en formas sutiles de los fragmentos (Burrough, 1981; Krummel et al., 1987; Turner, 1989; Sugihara & May, 1990; Ripple et al., 1991; MacGarigal & Marks, 1995).
La forma de los fragmentos, calculada a través del factor forma fue un buen indicador con respecto a la circularidad de los fragmentos, ya que utiliza únicamente el perímetro de éstos, sin embargo, debe ser tomada con precaución debido a que este factor es afectado por la escala y a que es menos sensible a formas distintas a círculos.
En opinión de Helzer & Jelinski (1999) la forma de los fragmentos es una medida efectiva de la calidad de éstos, comparada con el área, debido a que ésta refleja además de la forma, el tamaño de los mismos.
El hecho de que el área total de los fragmentos de vegetación nativa disminuyera significativamente a través del tiempo (tres décadas) pone en evidencia que la fragmentación en la cuenca del Río Damas es un proceso lento, pero constante, lo cual indica que, a menos que se tomen medidas correctivas de inmediato, el proceso va a continuar, afectando core ello las condiciones de vida, tanto para la flora como para la fauna de la región, además de procesos climáticos asociados al papel regulador de la vegetación, tales como: regímenes de precipitación, temperatura, humedad relativa y control de caudales.
La hipótesis del aislamiento biogeográfico (Preston, 1962; MacArthur & Wilson, 1967) predice que áreas (o fragmentos) pequeñas sostendrán menos especies que áreas más grandes, debido al balance que existiría entre las tasas de inmigración y extinción de las especies. Con base en esto, se puede predecir una pérdida de especies en la cuenca del Río Damas, sin embargo es necesario desarrollar investigaciones que relacionen el área de los fragmentos con la diversidad y riqueza de especies, de tal forma que aporten evidencia con respecto a la hipótesis del aislamiento biogeográfico.
CONCLUSIONES
La dinámica de fragmentación temporal en la cuenca del Río Damas varió significativamente; el número de fragmentos decreció en un 30 % en el período analizado (30 arios), así como el área total ocupada por éstos (6% con respecto al total). Los fragmentos variaron de formas circulares (1966) hacia formas amorfas y elongadas (1996), esta variación fue significativa a través del tiempo.
El índice de diversidad de Patton se comportó como un buen indicador de formas gruesas; la dimensión fractal reflejó habilidad en captar formas finas, y el factor forma se comportó como un indicador de formas circulares. En caso de requerir evaluar paisajes a diferentes escalas, resulta apropiado el uso de la dimensión fractal en la estimación de la forma de los fragmentos.
La variación en la forma de los fragmentos fue estadísticamente significativa, en la dinámica temporal del paisaje investigado, e indica una tendencia a fragmentos de áreas pequeñas y medianas (entre 5 y 30 ha.) de formas amorfas, elongadas e irregulares con mayor perímetro.
Si el proceso de fragmentación en la cuenca del Río Damas continúa como hasta ahora, la proporción de remanentes de vegetación nativa declinará, en consecuencia, los remanentes de vegetación en un futuro podrían estar dominados por especies de alta movilidad por factores abióticos, invasoras, exóticas y/o heliófitas; además la dinámica de perturbación humana no permitiría procesos de sucesión vegetal; así, las especies incapaces de colonizar áreas relativamente aisladas podrían progresivamente limitar su presencia en los fragmentos lo cual incrementaría su vulnerabilidad a riesgos de extinción local.
Es necesario adelantar investigaciones con respecto a la conectividad entre fragmentos, el grado de aislamiento, la influencia de la matriz circundante y la riqueza de especies de los fragmentos.
RECOMENDACIONES
Con base en las conclusiones anteriores, algunas sugerencias a tener en cuenta para lograr un manejo apropiado de los fragmentos de la cuenca del Río Damas son:
· Impedir que se eliminen los fragmentos de vegetación nativa remanentes, proteger aquellos de áreas grandes y particularmente de áreas pequeñas; éstos dan la oportunidad para mantener poblaciones de especies a través de un "efecto de rescate". La presencia de fragmentos pequeños ofrece al menos la oportunidad de persistencia para ciertas especies, más que paisajes completamente deforestados.
· Modificar la forma de fragmentos elongados de tal forma que se acerquen más a la circularidad, mediante la construcción de cercas de protección y el favorecimiento de la regeneración natural.
· Conservar los fragmentos de vegetación ribereña, los cuales pueden ser utilizados como corredores de conectividad entre fragmentos.
· Considerar el efecto de la matriz que rodea a los fragmentos, pues matrices "amigables" permiten una recuperación y colonización de especies más rápida que matrices "hostiles".
· Fomentar la conectividad de los fragmentos de tal forma que se generen corredores que faciliten la dispersión de las especies.
BIBLIOGRAFÍA
- Burrough P. 1981. Fractal dimensions of landscapes and other envrionmental data. Nature Vol 294 :240-242.
- Corporación Nacional Forestal, CONAF & Comisión Nacional del Medio Ambiente - CONAMA- 1997. Catastro y evaluación de los recursos vegetacionales de Chile. Base de datos SIG- Xa Región.
- Donoso C. 1981. Tipos forestales de los bosques nativos de Chile. Corporación Nacional Forestal y Organización de las Naciones Unidas para la Agricultura y la Alimentación, Documento de trabajo N° 38.
- Donoso C. & Lara A. 1997. Utilización de los bosques nativos en Chile: pasado, presente y futuro. En: Ecología de los bosques nativos de Chile. Armesto J., C. Villagrán & M.K. Arroyo(Eds). Editorial Universitaria S.A. Santiago,Chile. 477.
- Fallas J. 1998. Sistemas integrados de Información geográfica. Laboratorio de teledección y sistemas de información geográfica. Universidad Nacional. Heredia, Costa kica.
- Forman R T.T. 1995. Land Mosaics, the ecology of landscapes and regions. Cambridge University Press, Great Britain.
- Forman R.T.T. & M. Godron. 1986. Landscape ecology. John Wiley and Sons.
- Fuentes Q.E. 1994. ?Qué futuro tienen nuestros bosques¿. Hacia la gestión sustentable del paisaje del centro y sur de Chile. Ediciones Universidad Católica de Chile. Santiago,Chile.289 p.
- Gantz A & J. Rau. 1999. Relación entre el tamaño mínimo de fragmentos boscosos y su riqueza de especies de aves en el sur de Chile. Anales del Museo de Historia Natural de Valparíso 24:85-90.
- Harris L.D. & G. Silva-López. 1992. Forest fragmentation and the conservation of biological diversity. In: Conservation biology: the theory and practice of nature conservation and management. Fiedler P.L & 5.1K. Jain, Editors. Chapman & Hall. New York, U.S.A 507 p.
- Harris L.D. 1984. The fragmented forest. The University of Chicago Press. 200 p. Chicago, U.S.A.
- Helzer Ch & D. E. Jelinski. 1999. The relative importance of patch arca and perimeter-area ratio to grassland breeding birds. Ecological Applications 9(4):1448-1458.
- Henao S. 1988. Introducción al manejo de cuencas hidrográficas. Universidad de Santo Tomas. Centro de Enseñanza Desescolarizada. Ediciones Usta. Bogotá, Colombia. 395 p.Instituto Nacional de Estadísitcas INE. 1997. VI Censo Nacional Agropecuario, Resultados preliminares 1997. INE. 443 pp.
- Krummel J.R.; R.H Gardner; G. Sugihara; R.V O'Neill & P.R. Coleman. 1987. Landscape patterns in a disturbed environment. Oikos 48 : 321 - 324.
- Laurance WF. & R.O. Bierregaard. 1997. Tropical Forest Remnants Ecology, Management and Conservation of Fragmented Communities(Abstracts on une www.press.uchicago.edu/Images/Chicago/rbullet.gif).
- Laurance W.F & E. Yensen. 1991. Predicting impacts of edge effects in fragmented habitats. Biological Conservation 55:77-92.
- Laurance WE; L.V Ferreira; J.M Rankin-de Merona; S.G. Laurance; R.W. Hutchings & T.E. Lovejoy. 1998. Effects of forest fragmentation on recruitment patterns in amazonian tree communities. Conservation Biology 12(2):460-464.
- Lord J.M. & D.A. Norton. 1990. Scale and the spatial concept of fragmentations. Conservation Biology 4:197-202.
- Lovejoy TE. 1982. Area-perimeter relation for rain and cloud arcas. Science Vol 216:185 :187.
- Lovejoy T.E.; R.0 Bierregaard; A.B. Rylands; J.R. Malcom; K.S. Brown; C.E. Quintela; L.H. Harper; A.H. Powell; G.V. PowelL; H. Shubart & M.B. Hays. 1986. Edge and other effects of isolation in amazon forest fragments. In: Conservation Biology : the science of scarcity and diversity. Soulé M.E(Ed.). Sinauer Associates. Sunderland, Massachussets, U.S.A 584 p.
- MacArthur R.H. & E.O. Wilson. 1967. The theory of island biogeography. Monographs in Population Biology N° 1. Princeton University Press. New Jersey. 215 p.
- Mansilla, E. 1995. Auditoría ambiental a las actividades humanas de la cuenca del Río Damas. Seminario de título para optar al título de Técnico Universitario en Control Ambiental. Universidad de Los Lagos. Sede Puerto Montt. Puerto Mont, Chile.
- McGarigal, K & B. Marks. 1995. Fragstat's, spatial patterns analysis program for quantifying lansdscape structure. Portland OR: U.S. Department of Agrigulture, Forest Service, Pacific Northwest, Research Station. 122 13.
- Meffe G.K. & C.R. Carroll. 1994. Principies of conservation biology. Sinauer Associates, Inc. Sunderland, Massachusetts. 600 p.
- Moreno, L.A. 1994. Estudio hidroambiental del Río Damas (X Región): Análisis de la situación actual. Tesis de grado, Facultad de Ingeniería. Departamento Ingeniería Civil. Universidad de concepción. 182 pp.
- Myers N. 1994. Global biodiversity II : losses and threats. In: Principies of Conservation Biology. Meefe G.K. & C.R. Carroll. (eds). Sinauer Associates Pulishers. Sunderland, Massachusetts, U.S.A 600 p.
- Noss R.F. & B Csuti. 1994. Habitat fragmentation. In: Principies of Conservation Biology. Mefee G.K. & Carroll (eds).).Sinauer Associates Publishers. Sunderland Massachusetts, U.S.A. 600 p.
- Patton D.R. 1975. A diversity index for quantifying habitat edge. Wildlife Society Bulletin 3: 171 - 173:
- Pimm L.S. 1998. The forest fragment classic. Nature 393: 23-24.
- Preston F.W. 1962. The canonical distribution of communnes and rarity. Ecology 43 :185- 215, 410-432.
- Ripple J.; G.A Bradshaw & TA Spies. 1991. Measuring forest landscapes patterns in the Cascade Range of Oregon, USA. Biological Conservation 57: 73-88
- Shafer C.L. 1990. Island theory and conservation practice. Smithsoniam Institution Press. Washington and London. 189 p.
- Sugihara G. & May R. 1990. Applications of fragtal in ecology. Trends in Ecology and Evolution. Vol 5, N° 3: 79: 86.
- Turner M.G. 1989. Landscape ecology: the effect of pattern on process. Annu. Rey. Ecol. Syst. 20: 171-197.
- Willson, E.O. & Willis E.O. 1975. Applied biogeography . In: ,Cody, M.L. & Diamond J.M. (Eds). Ecology and the evolution of communities. Harvard Univ. Press. Cambridge, -M.A pp 522-534.
Creation date:




Licencia
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






