
DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2004.1.a03Publicado:
01-01-2004Número:
Vol. 8 Núm. 17 (2004)Sección:
Artículos de investigación científica y tecnológicaIncidencia de las micorrizas arbusculares y vesículo arbusculares como estrategia adaptativa de especies de páramo y selva altoandina, cordillera oriental de Colombia
Incidence of arbuscular mycorrhizae and arbuscular vesicles as an adaptive strategy for species of páramo and jungle altoandina, eastern cordillera of Colombia
Palabras clave:
Mycorrhiza, incidence, adaptation, páramo, andean jungle, states of development (en).Palabras clave:
Micorriza, incidencia, adaptación, paramo, selva altoandina, estados de desarrollo (es).Descargas
Referencias
AZCON, C. & BÁREA, J. 1996. Interacciones de las micorrizas arbusculares con microorganismos de la rizósfera. Departamento de Microbiología. Estación Experimental del Zaidín, CSIC. Granada, España. En: Guerrero, E. 1996. Micorrizas. Recurso biológico del suelo. Fondo FEN Colombia. 208 pp.
BORIE F. 1983. Fósforo orgánico y micorrizas "VA" en suelos volcánicos de Chile. Universidad de la Frontera, Temuco (Chile). En: Técnicas de investigación de micorrizas, Fundación Interamericana para la ciencia (FIC), Centro Agronómico Tropical de Investigación y Enseñanza.
CARRIZOSA, J. y HERNÁNDEZ J. 1990. Selva y futuro en Colombia. El sello editorial. Bogota.
COBA, B. Y COGUA 1995. Reconocimiento de MVA en el Páramo y Bosque Altoandino en la Región de Monserrate; En : Estudios Ecológicos del Páramo y del Bosque Altoandino, Cordillera Oriental de Colombia. p. 439.
CORREA RESTREPO, M. ; MORALES, M., COBA de G., B y E. SIEVERDING 1990. Presencia de Micorriza vesículo arbuscular (MVA) en un relicto de bosque andino, en la vereda " Alta Charco" departamento de Cundinamarca, Colombia. En : Resúmenes del V Congreso Latinoamericano de Botánica y I Simposio Latinoamericano de Micorrizas. Impreso Palacio de las comunicaciones. Cuba. 399 pp.
CORREA DE RESTREPO , M., MONTENEGRO , J., MARTÍNEZ, J., TORRES, E. y A. CAICEDO 1993. Análisis sobre la actividad de hongos formadores de micorrizas vesículo arbusculares. En : " Aspectos ambientales para el ordenamiento territorial del occidente del departamento del Caquetá ". Tomo II. Sección 4 :698-735. Instituto Geográfico Agustín Codazzi (IGAC). Ed.
GUERRERO, E.1993. Evaluación de las micorrizas en ecosistemas andinos del Parque Nacional Natural Chingaza primera etapa: Páramo. Bogota, Colombia. Departamento de Biología ,Facultad de Ciencias Pontificia Universidad Javeriana.
HERRERA, J. F., & F. E. RUIZ. 1981. Algunos aspectos de la ecología y de los efectos inmediatos del fuego sobre la antropofauna asociada a Espeletia grandiflora H & B. Páramo de Monserrate, Bogotá, Colombia. Tesis Departamento de Biología Universidad Nacional de Colombia. Bogotá.
SIEVERDING. E. 1991: Vesicular Arbuscular Mycorrhyza Management Tropical Agrosystems. Technical Cooperation Federal Republic of Germany. GTZ No. 224.
DEPARTAMENTO DE AGRICULTURA Y SUELOS DE LOS ESTADOS UNIDOS. 1999. Soil taxonomy.
UNIDAD ADMINISTRATIVA ESPECIAL DEL SISTEMA DE PARQUES NACIONALES NATURALES, (UAESPNN) MINISTERIO DEL MEDIO AMBIENTE. 2000. Plan Estratégico y de Manejo para el Parque Nacional Chingaza y su Zona de influencia 2000-2005. p. 10-35.
VALENCIA H, MURILLO, MOYANO. 1995. Micorrizas Vesículo-Arbusculares asociadas con tres especies nativas de páramo y bosque altoandino en la región de Monserrate, Colombia. En : Estudios Ecológicos del Páramo y del Bosque Altoandino, Cordillera Oriental de Colombia. p. 449.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Artículo de Investigación científica y tecnológica
Colombia Forestal, Diciembre del 2004 vol:8 nro:17 pág:43-59
Incidencia de las Micorrizas Arbusculares y Vesículos Arbusculares como estrategia adaptativa de especies de páramo y selva Altoandina, Cordillera Oriental de Colombia
Juan Francisco García Romero
Daniel Camila García Fernandéz
Ingenieros forestales, Universidad Distrital Francisco José de Caldas.
Marina Correa de Restrepo
Directora de la Investigación, Profesor Asociado Departamento de Biología, Universidad Nacional de Colombia.
Resumen
La presente investigación determinó mediante un análisis cualitativo y cuantitativo la incidencia de las micorrizas arbusculares y vesículo - arbusculares (MA-VA), en la adaptación de especies vegetales de páramo y selva altoandina, evaluando tres estados de desarrollo de la planta en dos épocas climáticas. En un área natural, Parque Nacional Natural Chingaza y en un área con disturbios Páramo el Granizo, Región de Monserrate Se emplearon las técnicas descritas para investigación en micorrizas (Correa et al., 1993).
Se evidenció claramente la importancia de la relación de los hongos MA-VA con las especies en estudio en función de su estado de desarrollo. Se observó como la dinámica del hongo se ve influenciada por las variaciones climáticas, siendo muy marcada su actividad en la época seca. La incidencia de los MA-VA como estrategia de adaptación de estas especies a las condiciones medioambientales extremas fue uno de los aportes más relevantes del presente estudio, debido a que se comprobó la prevalencia de la simbiosis en los estados iniciales en la mayoría de las variables de respuesta evaluadas.
Palabras Clave: Micorriza, incidencia, adaptación, páramo, selva altoandina, estados de desarrollo.
Abstract
The incidence of the arbuscular and vesicular - arbuscular mycorrhiza fungi (A-VAM) in the adaptation of vegetables species from the paramo and high andean jungle measured by three states of development of the plant in two climatic conditions, using quantative and qualitative analysis. The techniques used are described by Correa et al., 1993.
The importance of the relation of the fungus AVAM to the studied species, acording to the states of development, was clearly evident. It was observed that the dynamics of the fungus is influenced by the climatic variations, having marked activity during the dry season. The indidence of the A-VAM as strategy of adaptation of this species in extreme enviromental conditions was one of the createst contribution of the present studie because it was proved the prevalence of symbiosis in the initial states of in most of the avalueted variables.
Key Words: Mycorrhiza, incidence, adaptation, páramo, andean jungle, states of development.
INTRODUCCION
En la actualidad los ecosistemas de páramo y selva . altoandina presentan un alto grado de intervención por el desarrollo de actividades económicas como agricultura, ganadería y minería, así como el establecimiento de asentamientos humanos, lo cual ha repercutido en un rápido deterioro de las condiciones medioambientales.
En lugares de alta montaña donde la alteración antrópica se ha detenido y no se presentan daños irrecuperables, se puede observar un avance de la vegetación de páramo (fenómeno de paramización); además en algunos lugares se observa también un reestablecimiento de especies inductoras propias de la selva altoandina.
Para desarrollar actividades encaminadas a la recuperación de los ecosistemas altoandinos es preciso considerar el suelo y su microbiota asociada como la base del sustento para el establecimiento, adaptación y desarrollo de las comunidades vegetales allí encontradas.
La investigación de los recursos biológicos en países megadiversos como el nuestro ha venido concentrándose en especies de fauna y la flora, lo cual constituye un aporte valioso pero que desconoce otro componente del mundo biótico como lo son los microorganismos del suelo (hongos filamentosos, micorrizas, bacterias, etc.).
El papel de las micorrizas arbusculares y vesículoarbusculares, demuestra ser un factor determinante en el ciclaje de nutrientes y la adaptación a condiciones extremas de clima y suelos, lo cual se refleja en el alto número de especies vegetales que establecen simbiosis con estos hongos. (Correa et al., 1990).
En los ecosistemas andinos se ha establecido la presencia de asociaciones micorrícicas en una gran cantidad de especies (Coba et al., 1995; Guerrero, 1993; Valencia et al., 1995). Los estudios realizados se han centrado principalmente en el reconocimiento de la asociación, pero no se ha determinado la influencia de ésta en los procesos de adaptación (objeto del presente estudio) así como el desarrollo de las especies vegetales a lo largo de sus diferentes etapas de vida, lo cual seria el componente básico de procesos encaminados a la restauración ecológica de estos ecosistemas.
OBJETIVO GENERAL
Determinar la incidencia de las Micorrizas Arbusculares (MA) y Vesículo-Arbusculares (VA), como estrategia adaptativa de especies vegetales de páramo y selva altoandina, en un área intervenida con procesos de paramización y en otra en estado natural.
OBJETIVOS ESPECÍFICOS
• Cuantificar los porcentajes de infección y diversidad de hongos MA-VA a partir de un muestreo de raíces y rizósfera de las especies seleccionadas en ambos ecosistemas.
• Relacionar los porcentajes de infección, la diversidad y número de esporas en tres estados de desarrollo (regeneración, intermedios y adultos) de las especies vegetales en estudio.
• Relacionar las características físico-químicas del suelo (textura, pH, contenido de fósforo y aluminio), con los porcentajes de infección y la diversidad de morfoespecies de esporas de hongos MA-VA, en las raíces y rizósfera de las especies vegetales estudiadas.
• Relacionar datos de precipitación en dos épocas del ario (seca y húmeda), con los porcentajes de infección y a la diversidad de morfoespecies de esporas de hongos MAVA en las raíces y rizósfera de las especies vegetales en estudio.
• Comparar los porcentajes de infección y la diversidad de hongos MA-VA en los tres estados de desarrollo de un área intervenida y otra con baja intervención.
HIPÓTESIS DE TRABAJO
Las micorrizas tienen la propiedad de absorber agua y nutrientes y transportarlos hacia las plantas, no sólo con aquellas que forman simbiosis si no también con las que se conectan a través de la estera de raíces que se forman en las comunidades vegetales asociadas.
La actividad de las MA-VA es mayor en los estados iniciales de desarrollo y en áreas con baja o nula intervención, por tanto inciden en la adaptabilidad de las especies en estudio.
CONCEPTOS BÁSICOS
La presencia de especies de Espeletiay Espeletiopsis dentro de la selva altoandina con frecuencia se debe a la intervención humana, pues a menudo, como sucede con Espeletiopsis corymbosa en los cerros de Bogotá, es común en etapas subseriales o degradadas de la selva altoandina, en las cuales las comunidades son apenas arbustivas. Pero en otros casos, si bien la tendencia heliófila de dichos géneros propicia la ocupación de sitios degradados de la selva altoandina, es posible que algunas áreas que actualmente pueden a primera vista considerarse como verdaderos páramos, no sean otra cosa que etapas subseriales de selva altoandina cuyo arbolado fue eliminado por el hombre. Dicho de otra manera parte de los páramos y subpáramos actuales no correspondería al clímax sino a etapas subseriales, y las especies vegetales que allí se desarrollan serán elementos heliófilos oportunistas con amplia tolerancia ecológica que usualmente se hallan dentro de la selva altoandina clímax, pero sin alcanzar allí la importancia fitosociología que pueden lograr cuando se elimina el dosel arbóreo y se degrada el sotobosque. (Carrizosa y Hernández, 1990).
HONGOS FORMADORES DE MICORRIZA
Micorriza es la asociación mutualista entre algunos hongos del suelo y la raíz de las plantas. Es el producto de un proceso de coevolución entre plantas y hongos, como parte del avance colonizador de las plantas acuáticas primitivas hacia el medio ambiente terrestre. En ella, el micelio del hongo infecta la corteza radical a modo de endófito y proyecta sus hifas tanto al interior como al exterior de la raíz. La micorriza es la infección fúngica más extendida en el reino vegetal y los hongos micorrizógenos contribuyen de manera sustancial a la biomasa del suelo (Guerrero, 1993).
La función principal de esta asociación desde el punto de vista de la planta, es aumentar la superficie de absorción de la raíz de tal forma que ésta absorba y asimile más agua y minerales en comparación con una raíz no micorrizada. En consecuencia la micorriza es una adaptación biológica que favorece a las plantas en su nutrición mineral, balance hídrico y desarrollo (Azcón y Bárea, 1996).
El carácter mutualista supone también un beneficio para el hongo, que consiste en el suministro, por parte del macrosimbionte, de compuestos orgánicos y de un hábitat protegido. Esto significa que la simbiosis tiene un costo energético para la planta en términos de producto fotosintético, costo que normalmente está compensado por las ventajas que trae la asociación.
La micorriza favorece en particular la absorción de iones poco móviles como H 2PO4- , NH4+, Zn", Cu", gracias a la elongación del micelio externo más allá de la zona de agotamiento de nutrientes alrededor de la raíz (Guerrero, 1993). Esto explica la alta dependencia de la micorriza que manifiestan las plantas cuando crecen sobre suelos pobres en nutrientes.
FUNCIONAMIENTO DE LA RELACIÓN PLANTA — HONGOS MA-VA.
La simbiosis representa un proceso sucesivo de intercambios de sustancias nutritivas, metabólitos esenciales, sustancias hormonales, etc., entre dos partes, resultando en un beneficio mutuo para ambos. Esta asociación se rige fundamentalmente por los genomas de la planta y el hongo, condicionada por el medio ambiente (Guerrero, 1993). En la infección del hongo en las raíces se produce:
• Incremento de la síntesis de proteínas, clorofila, sustancias de crecimiento y metabolitos secundarios.
• Favorecimiento de la absorción, translocación de nutrientes y agua.
• Aumento de la tasa fotosintética.
• Aumento en la diferenciación de los tejidos vasculares.
• Generación de nuevos organelos y del grado de vacuolación de las células corticales.
ZONAS DE ESTUDIO PÁRAMO EL GRANIZO
Se encuentra localizado en los cerros orientales de la Sabana de Bogotá, región de Monserrate, en jurisdicción del Distrito Capital y el municipio de La Calera, entre los 4°45'-4 057" de latitud norte y los 73°8-74°1' longitud oeste, entre 3.000 y 3.200 m.s.n.m. y perteneciente a la cuenca media del río Bogotá y a la microcuenca de la quebrada "El Granizo". Forma parte de la Reserva Forestal de la Empresa de Acueducto y Alcantarillado de Bogotá y terrenos de propiedad privada (Herrera y Ruiz, 1981).
El área de estudio presenta evidentes signos de intervención de épocas pasadas, y aun en la actualidad aunque en una menor escala, por lo que se puede apreciar el avance de procesos de paramización.
La temperatura media anual oscila entre 9° y 14°C. El promedio multianual mensual de la temperatura media en el páramo abierto presenta una oscilación menor de 2°C.
La precipitación media anual es de 700 a 2500 mm/año. El promedio multianual más bajo se presenta en enero (43.52 mm). La precipitación más alta ocurre en los meses de mayo (91.48 mm) y octubre (95.41 mm). Estos máximos multianuales promedio señalan también las dos estaciones anuales de lluvia del páramo y el régimen bimodal de las precipitaciones; en cuanto que dos estaciones menos lluviosas (secas) alternan con dos estaciones con mayor precipitación.
De acuerdo a Departamento de Agricultura y Suelos de los Estados Unidos (1999) los suelos pueden pertenecer a los órdenes Inceptisoles y Andisoles de acuerdo a su grado de evolución. En la región del páramo y de la selva se clasifican como Dystric cryandept. Los cuales presentan alta acidez, bajo nivel de nutrientes y baja concentración de fósforo lo que lo identifica como distrófico. (Coba y Cogua, 1995).
En la región del Granizo se han presentado intervenciones antrópicas en la mayor parte de su extensión, ocasionando la pérdida de la vegetación natural, sin embargo se encuentran relictos de selva altoandina en diferentes estados de sucesión presentando asociaciones típicas como Weinmannia-Drimys y en las partes altas alternado con la asociación de Calamagrostis-Espeletia.
PARQUE NACIONAL NATURAL CHINGAZA
Se ubica en la Cordillera Oriental al Nor-oriente de Bogota, entre los 73°30' y los 73°55' de longitud oeste y los 4°20' y 4°50' de latitud norte. Cuenta con aproximadamente 76.600 ha de superficie, entre los 1.000 y los 4.000 m.s.n.m.
La temperatura media para el cuerpo principal del parque oscila entre 6 y 7°C y sobre la franja Suroriental la variación se produce desde los 6 hasta los 11°C a medida que se desciende altitudinalmente. La distribución dentro del ario de las altas y bajas temperaturas operan en forma simétrica e inversa a la de la precipitación, observándose sin embargo que la diferencia de temperaturas del mes más frío y al más cálido no alcanzan los 5°C. (UAESPNN, 2000).
El régimen de precipitaciones tiene características particulares que denotan la influencia del tipo de lluvias que se presentan en los llanos orientales. Sobre la franja Sur-oriental caen en promedio unos 4500 mm de precipitación al ario y van disminuyendo gradualmente hasta unos 1200 mm sobre el costado oeste del parque. La distribución dentro del ario de las precipitaciones medias anuales muestra un solo pico de lluvias intensas hacia los meses de Mayo a Agosto, los meses de Diciembre a Febrero son los de menores precipitaciones.
En condiciones de relieve plano o escasa pendiente y buen drenaje, los suelos pueden ser moderadamente evolucionados (Inceptisoles), del orden Tropepts o Andepts cuando en su génesis han participado cenizas volcánicas provenientes de erupciones ocurridas en la Cordillera Central y transportados por corrientes de aire. Donde las pendientes son pronunciadas existen suelos de poco espesor y pobremente evolucionados (Entisoles), que alcanzan a ser ranqueriformes en pendientes próximas a 45%.
ESPECIES EN ESTUDIO
Para la selección de las especies del estudio (Tabla 1) se consideró la posición ecológica y sucesional de éstas, así como su representatividad dentro del ecosistema. Se tuvo en cuenta el tipo de sistema radical para cada especie dada la importancia de la rizósfera en el establecimiento de la simbiosis.
METODOLOGÍA
GENERALIDADES
Para establecer si las MA-VA presentan incidencia en la adaptación de algunas plantas, se planteó evaluar el porcentaje de infección en raíces y número de esporas y morfoespecies presentes en la rizósfera de 3 especies de páramo (Espeletia grandiflora y Espeletiopsis corymbosa asociados a Calamagrostis effusa) y en 3 de selva altoandina (Weinmannia tomentosa y Drimys granadensis asociados a Chusquea scandens) en un área intervenida del páramo "El Granizo" y un área de baja intervención en el Parque Nacional Natural Chingaza.
Las especies E. corymbosa y Ch. scandens no se encontraron en los ecosistemas del Parque Nacional Natural Chingaza, por tanto se tomaron muestras de Espeletia argentea y Chusquea weberbauerii, las cuales presentan una similitud con las especies, presentes en el páramo El Granizo, en cuanto a características fisiológicas y fisonómicas y la posición ecológica y sucesional dentro del ecosistema.
Para la selección de las zonas de muestreo en cada una de las áreas planteadas se consideraron factores como el estado de intervención, composición floristica y altura sobre el nivel del mar.
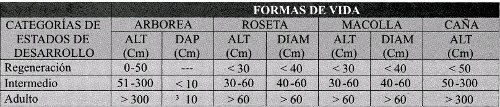
A nivel de especie se seleccionaron individuos en tres diferentes estados de desarrollo, para comparar las variables de la infección en función a éstos. Los parámetros para seleccionar los estados de desarrollo (regeneración, intermedio y adulto), fueron la altura y el diámetro de acuerdo a su forma de vida: arbórea, macollo, rosetas y cañas. (Tabla 2). El diámetro para las especies arbóreas se tomó a la altura del pecho (DAP) y para las rosetas y macollas donde presentaran el mayor diámetro.
En el páramo El Granizo se tomaron muestras en tres zonas diferentes, recolectando 3 repeticiones de cada uno de los estados de desarrollo por especie. El muestreo se realizó en dos épocas del ario que presentaron diferencia de precipitación, (Húmeda, Octubre/2002 y Seca, Febrero/2003)
En el Parque Nacional Natural Chingaza se tornaron muestras de cada especie en los tres estados de desarrollo planteados en dos áreas en condiciones naturales de los ecosistemas páramo (sector de Piedras Gordas) y selva altoandina (Bosque El Encenillo, sector de Monteredondo). Estas muestras se tomaron en época húmeda, Octubre / 2003 (especies de selva altoandina) y en Diciembre /2003 (especies de páramo).
VARIABLES A EVALUAR EN CADA UNA DE LAS ESPECIES
Para evaluar la actividad de las MA-VA a nivel intraespecífico se consideraron los estados de desarrollo de las especies hospederas estudiadas.
Se evaluó el clima (Precipitación) y características físico químicas del suelo (Tabla 3)
EVALUACIÓN DE LA MICORRIZA
Según el tipo de vegetación y suelos se tomaron muestras de material a 5, 10 y 20 cm de profundidad y desde 30 cm a 1 m o más de distancia del fuste de las plantas, según la profundidad a que se encontraron las raíces finas y fueron etiquetadas y transportadas en bolsas de polietileno a los laboratorios para ser procesadas, mediante el método descrito por sierverding (1991), con modificaciones de Correa et al., 1993.
El clareado y tinción de raíces así como la cuantificación de los hongos en las mismas y el aislamiento de esporas se hizo según el método de Sieverding, con modificaciones de Borie, (1983), para suelos con influencias de cenizas volcánicas.
Para el análisis de datos se utilizó el paquete SAS 8.0. Se realizó un análisis de varianza de una vía con un nivel de significancía del 0.1. Se tomaron tres tratamientos a nivel intraespecifico de los estados de desarrollo (regeneración, intermedios y adultos) y como variables respuesta, el número de esporas / 100 gr de suelo, y el porcentaje de infección total, por hifas intramatricales y extramatricales, vesículas y arbúsculos. Los datos expresados en porcentaje se transformaron mediante la aplicación de la función ¨Arcoseno¨ X¨.
Cuando se encontraron diferencias significativas entre tratamientos se realizó una prueba de comparación de medias de Tukey a un nivel de significancía de 0.1 para establecer en que estados de la planta se encuentran las diferencias.
RESULTADOS
PÁRAMO EL GRANIZO
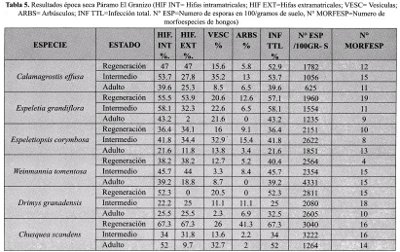
Los resultados encontrados de las variables evaluadas en los tres estados de desarrollo para la época húmeda se presentan en la Tabla 4 y los correspondientes a la época seca se encuentran en la Tabla 5.
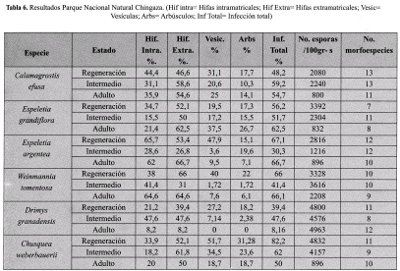
PARQUE NACIONAL NATURAL CHINGAZA
Los resultados encontrados en los tres estados de desarrollo en el área de baja intervención dentro del Parque Nacional Natural Chingaza se presentan en la Tabla 6.
DISCUSIÓN DE RESULTADOS
Las discusiones presentes en este artículo hacen referencia a la comparación de las variables tomando únicamente para el caso de la micotrofía la infección total, número de esporas en 100 gr. de suelo y el número de morfotipos.
MICOTROFIA EN EL PÁRAMO EL GRANIZO
En las especies de páramo estudiadas en El Granizo, la micorrización expresada como el porcentaje de infección en raíces, presentó los niveles más altos en los estados de regeneración e intermedios, para las dos épocas del año. A la vez que los niveles más bajos se encontraron en los estados adultos cumpliendo así con la hipótesis planteada en este estudio.
La excepción a este comportamiento se vio en Calamagrostis effusa, donde la infección en raíces para la época húmeda presentó el mayor porcentaje de infección total en el estado adulto. Esto puede estar dado por ser esta una Gramínea, que presentan un alto grado de micotrofía. En los estados maduros el desarrollo de procesos fisiológicos puede exigir una simbiosis más activa. Sin embargo la variación frente a los otros dos estados no fue estadísticamente significativa lo que evidencia una infección total de hongos MALVA estable con relación a los estados de desarrollo.
La diversidad de morfoespecies asociados a Calamagrostis effusa fue la más alta encontrada entre las especies estudiadas, mostrando una marcada diferencia entre las dos épocas del ario, siendo mayor en la época húmeda que en la época seca. La interpretación a este comportamiento puede estar dada por la presencia de especies más agresivas de MA-VA que presentan una propagación tanto en la época seca como en la húmeda, mientras que otras solo emiten sus propágulos al verse sometidos a una situación *de estrés, en este caso hídrico.
El número más alto de morfoespecies se presentó en el estado intermedio durante la época húmeda (40 morfoespecies), mostrando mayores diferencias con el estado adulto que con el estado de regeneración.
En cuanto al número de esporas / 100 gr. de suelo, se pudo observar una relación existente entre el estado de desarrollo y esta variable para ambas épocas climáticas, siendo mayor en el estado de regeneración seguido de intermedios y adultos. Esto puede estar causado por la presencia de un alto número de esporas latentes en la rizósfera, donde se establecen los estados iniciales disponibles para iniciar el proceso de colonización e infección de los sistemas radicales de los nuevos individuos.
La variación del número de esporas con relación a la época climática fue similar para los diferentes estados de desarrollo ya que todos presentaron para la época seca casi el doble del valor obtenido en la época húmeda lo cual indica la tendencia del hongo a propagarse en mayor proporción ante situaciones ambientales adversas (estrés hídrico)
Para el caso de Espeletia grandiflora la infección total de raíces presentó en época húmeda el comportamiento esperado con una diferencia significativa relacionada con los estados de desarrollo siendo mayor en el estado de regeneración seguido de los intermedios y adultos, presentó una diversidad mayor en la época húmeda que en la época seca, encontrando el número más alto de morfoespecies en el estado de regeneración (32 morfoespecies).
Espeletiopsis corymbosa, no presentó diferencias significativas en ninguna de las dos épocas, sin embargo se mantiene la relación de esta variable con los estados de desarrollo siendo mayor en regeneración, seguido de intermedios y adultos para la época húmeda. En la época seca la tendencia es de mayor valor para estados intermedios, seguido por regeneración y adultos, pudiendo deberse este comportamiento a una mayor actividad y desarrollo de procesos fisiológicos en el estado intermedio que condiciona la simbiosis.
En las especies de selva altoandina muestreadas en el páramo El Granizo, en la época húmeda, no se encuentra ninguna diferencia significativa en las especies arbóreas evaluadas, mientras que en la especie pionera Chusquea scandens hay correlación entre los estados de desarrollo y en la diversidad de hongos y el número de esporas.
En Weinmannia tomentosa se presentaron diferencias significativas durante la época seca, siendo mayor para los estados intermedios, seguidos de adultos y regeneración. Lo cual indica que en el caso de los estados iniciales de la planta, la simbiosis está más enfocada en satisfacer las necesidades nutricionales del hongo mediante el flujo de fotosintatos desde la planta hacia el simbionte; mientras que en los estados intermedios se presenta un mayor flujo de nutrientes minerales captados desde el suelo por el hongo hacia la planta, dado principalmente por el aumento de la superficie de absorción producida por las hifas extramatricales y una mayor transferencia de elementos por medio de los arbúsculos, a la vez que se desarrolla una alta colonización del hongo al interior de la raíz. Estos resultados se pueden interpretar desde el punto de vista fisiológico de las especies vegetales ya que en estados intermedios hay una mayor demanda de nutrientes para el desarrollo de procesos reproductivos (formación de flores y frutos) típicos de la fenología de las especies y el incremento en biomasa.
En cuanto a la diversidad de hongos MAVA asociados a la rizósfera de Weinmannia tomentosa se encuentran los mayores valores en la época húmeda en el estado intermedio con 22 morfoespecies. Para la época seca se presenta una disminución en el número de morfoespecies, presentando el menor valor de todas las especies estudiadas (4 morfoespecies). Esto demuestra como ante condiciones ambientales poco favorables sólo las especies de hongos más persistentes y adaptadas prevalecen.
El número de esporas es menor en la época húmeda que en la época seca, corresponde el mayor valor al estado de regeneración, seguida de intermedios y adultos. Para la época seca si se encuentran diferencias significativas entre los estados de desarrollo, presentando el mayor valor de todas las especies estudiadas en el estado adulto (4331), seguido de la regeneración y los intermedios. Este alto valor puede ser la respuesta del hongo a características muy particulares de los individuos como lo son condiciones micro ambiéntales, edáficas, fenológicas, entre otros.
En el caso de Drimys granadensis para la época seca se hallan diferencias significativas del porcentaje de infección en raíces entre los tres estados de desarrollo, siendo mayor en la regeneración para hifas intramatricales, vesículas e infección total, lo que indica una etapa de establecimiento y colonización por parte del hongo, a la vez que actúa como estrategia de adaptación de la planta en sus estados iniciales.
El número de esporas para la época seca, mostró diferencias significativas entre los tres estados de desarrollo, siendo mayor el valor en los estados de regeneración (2811), seguido por los intermedios y los adultos; el menor valor se encontró en el estado adulto de la época húmeda con 480 esporas / 100 gr. de suelo.
Chusquea scandens presenta una alta actividad de hongos MA-VA relacionada directamente con los estados de desarrollo, siendo mayor en regeneración, seguido de intermedios y adultos, sin embargo solo se encuentran diferencias significativas entre estados para la época seca, indicando los porcentajes de infección total más altos de todas las especies estudiadas. Este alto grado de asociación con hongos MA-VA puede estar relacionado directamente con el papel inductor de esta especie en los estadios sucesionales iniciales de la regeneración de la selva altoandina.
La diversidad de morfoespecies asociados a esta especie observó el mismo comportamiento que Calamagrostis effusa, presentando el mayor valor en el estado intermedio de la época húmeda con 26 morfoespecies y el menor valor en el adulto durante la época seca con 14 morfoespecies.
Se encuentran diferencias significativas entre estados en cuanto a número de esporas para época húmeda, donde conserva el mayor valor el estado de regeneración seguido por el intermedio y el adulto. En la época seca se encuentran as diferencias significativas para esta variable estando el mayor nivel en los estados intermedios seguido por la regeneración y los adultos. Este es un comportamiento atípico con relación al comportamiento seguido por la mayoría de las especies estudiadas, pudiendo deberse a características muy particulares de los individuos muestreados.
COMPARACION DE LA MICOTROFIA EN EL PÁRAMO EL GRANIZO CON EL PARQUE NACIONAL NATURAL CHINGAZA
Con el fin de evaluar y comparar el comportamiento de la simbiosis de hongos MA-VA con relación al grado de intervención del ecosistema, se llevó a cabo la comparación de los resultados obtenidos del muestreo de la época seca en el área intervenida en el páramo El Granizo con los datos procesados del trabajo en áreas conservadas dentro del Parque Nacional Natural Chingaza.
Se compararon las variables porcentaje de infección total, número de esporas y la diversidad de morfoespecies, relacionando a su vez las características edáficas como pH, contenido de Fósforo y contenido de Aluminio de cada una de las zonas. Para este artículo se hace referencia específicamente al porcentaje de infección total para comparar el comportamiento de las especies en las dos áreas.
Las especies vegetales en estudio comunes en las dos zonas fueron Calamagrostis effusa y Espeletia grandiflora entre las especies de páramo y Weinmannia tomentosa y Drimys granadensis entre las especies de selva altoandina.
Teniendo en cuenta el papel sucesional de C. difusa como especie pionera en la colonización de áreas sometidas a intervención en estos ecosistemas se puede considerar el papel que cumple la simbiosis micorrícica con mayor relevancia en las etapas iniciales durante el establecimiento de los individuos en condiciones poco favorables.
Al establecerse los estados más avanzados de desarrollo presentan características que los hacen más persistentes, como puede ser el sistema radicular y mayor capacidad fotosintética, por lo cual la dependencia de la simbiosis puede disminuir. En el caso de ecosistemas de baja intervención la infección puede tender a aumentar a medida que avanza el estado de desarrollo, principalmente por la presencia de una .mayor diversidad de especies vegetales ya establecidas que interactúan a través de la rizósfera y la estera de raíces formada por este tipo de comunidades.(Figura 1).
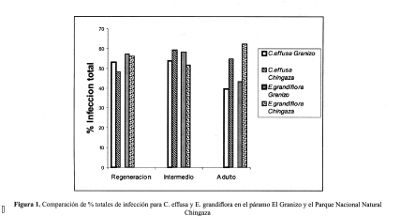
En Espeletia grandiflora los estad-os de regeneración e intermedio no presentan diferencias marcadas entre las dos zonas, siendo ligeramente mayor en el páramo El Granizo. Este comportamiento estable puede estar dado por su ciclo de vida más largo y por . su persistencia en los diferentes estadios de la sucesión. La baja variabilidad entre el área intervenida y el área con baja intervención para los estados de regeneración e intermedio puede estar dada por condiciones más favorables para su establecimiento generadas a partir de especies pioneras precursoras como Calamagrostis effusa, el mayor valor presentado en el estado adulto en Chingaza puede estar dado principalmente por la estera de raíces presente en el horizonte superficial del suelo y al aumento de la producción de necromasa en pie lo cual constituye una estrategia de este frailejón donde se sugiere un mecanismo de ciclaje directo de nutrientes evitando su perdida en el suelo, a través de la asimilación y translocación por parte de los hongos MA-VA. (Guerrero, 1993).
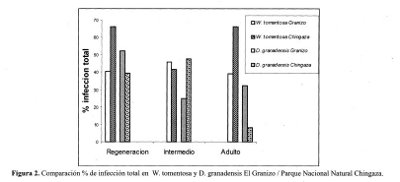
La infección total en raíces de W.tomentosa mostró valores iguales para los estados de regeneración y adulto en las dos zonas solamente la infección del estado intermedio fue superior a las infecciones encontradas en el páramo de Chingaza. Lo cual posiblemente está relacionado con la posición sucesional de esta especie, ya que al ser una dominante climácica de la selva altoandina, en un ecosistema como éste que presenta un estado de sucesión avanzado, ya se han establecido unas condiciones óptimas dadas principalmente por especies precursoras, lo que determina una mayor diversidad y por ende una alta actividad de la microbiota asociada en la rizósfera como es el caso de los hongos MA — VA.
En el caso del estado de desarrollo intermedio la presencia de una mayor infección en el área intervenida puede evidenciar, de acuerdo a la posición sucesional de esta especie, una mayor actividad de procesos fisiológicos enfocados a establecerse y dominar dentro del ecosistema.
El porcentaje de infección total en raíces de Drimys granadensis observó el mayor valor en el estado de regeneración en el área intervenida. El valor más bajo se encontró en el páramo de Chingaza en el estado adulto. Los valores de infección fueron mayores para el área de baja intervención solamente en el estado Intermedio.
Los niveles más altos de micotrofía observados para esta especie en el área intervenida puede tener relación con la posición sucesional, ya que al ser esta una especie inductora preclimácica del Weinmanniatum presentan un mayor actividad en los estados iniciales. El área del Granizo se encuentra en un estadio sucesional secundario temprano donde su papel es de gran relevancia para el acondicionamiento necesario del medio para el establecimiento de las especies dominantes.
El porcentaje de infección para esta especie en el Parque Nacional Natural Chingaza en el estado adulto mostró un valor muy bajo frente al encontrado en el área intervenida, lo cual puede estar en relación directa al estadio sucesional en el que se encuentra el Weinmanniatum alcanzando su estado clímax. Los individuos adultos presentes en esta asociación se caracterizan por sistemas radicales más lignificados y por tanto en su rizósfera se pueden restringir los procesos de intercambio con los hongos MA — VA. (Figura 2).
Las dos gramíneas comparadas en la selva altoandina muestran una alta micotrofía, presentando los porcentajes más altos de infección total en los estados de regeneración Ch. scandens y Ch. weberbauerii, disminuyendo gradualmente en los estados intermedio y adulto. Esta particularidad puede estar dada por la corta longevidad que presentan estas precursoras leñosas dentro de la sucesión, por lo cual es posible que la actividad de la simbiosis se intensifique en los estadios iniciales y su disminución gradual hacia el estado adulto está dada por su senescencia.
Dentro de los frailejones comparados se encontró un comportamiento inverso, donde E. corymbosa presentó niveles más bajos de infección en el estado adulto seguido por regeneración e intermedio, contrario a lo hallado en Espeletia argentea donde los estados de regeneración y adulto presentaron valores muy similares disminuyendo considerablemente para el estado intermedio. (Figura 3).
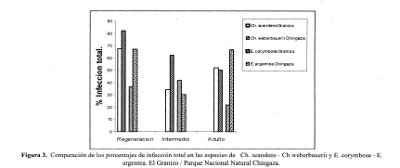
Los bajos niveles de infección en los estados iniciales de Espeletiopsis corymbosa pueden estar influenciados por el establecimiento de esta en zonas recién alteradas donde los disturbios han generado unas condiciones adversas para el desarrollo de la simbiosis por lo que los procesos de infección se pueden ver disminuidos. En el espeletion del Parque Nacional Natural Chingaza los niveles de infección para los estados de regeneración y adultos de E. argentea, pueden estar condicionados a la baja periodicidad de las alteraciones que pueda presentar el ecosistema, lo cual permite un posible equilibrio eh la asociación mutualista para estos dos estados de desarrollo.
Las muestras estudiadas provienen de suelos extremadamente ácidos, siendo mayor en el páramo de Chingaza. La vegetación abierta del Granizo está sometido a un mayor lavado de nutrientes y procesos de lixiviación debido a la precipitación. Estas condiciones pueden estimular un mayor crecimiento radical y por ende una mayor probabilidad de infección por parte de los hongos MA-VA, (Valencia et al., 1995).
En ambas zonas se encuentra una alta micotrofía así como bajos niveles de fósforo, sin embargo en el páramo de Chingaza se observan valores de infección un poco más altos, lo que puede estar relacionado con unas condiciones más estables dentro del ecosistema sin la presencia de disturbios o alteraciones recientes.
El contenido de aluminio presentó niveles altos para ambas áreas, siendo ligeramente mayor en el páramo sin intervención del Parque Nacional Natural Chingaza, lo que sumado a los mayores niveles de actividad micorrícica encontrados para esta zona puede indicar como ante limitantes como la toxicidad causada por la alto cantidad de aluminio los hongos MA-VA pueden determinar una mayor tolerancia de las plantas a estas condiciones.
CONCLUSIONES
• Las ocho especies incluidas en este estudio presentaron micotrofía con hongos formadores de MA-VA en los tres estados de desarrollo evaluados durante las dos épocas climáticas del ario siendo relativamente mayor en el Parque Nacional Natural Chingaza.
• Prevalecen los mayores valores en las variables mencionadas para el estado de regeneración de las especies en estudio lo cual indica el papel que ejerce la simbiosis con hongos MA-VA como estrategia adaptativa.
• Las variaciones en la precipitación tienen una incidencia directa con los porcentajes de infección, siendo mayor para la época húmeda.
• En términos generales las gramíneas tanto de selva altoandina como de páramo presentan una mayor actividad micotrófica, ratificando su papel como colonizador e inductor de la sucesión dentro del ecosistema.
• Los niveles de micotrofía en las especies estudiadas dentro del área de baja intervención en el Parque Nacional Natural Chingaza fueron en términos generales más altos que los presentados en el área del Granizo, a excepción de la diversidad que fue mayor en la zona con intervención.
• Los resultados obtenidos demuestran una relación directa entre la diversidad de hongos MA-VA y número de esporas presentes en la rizósfera con la dinámica sucesional del bosque, donde a medida que se alcanza una condición clímax la diversidad disminuye.
• Se encontró que la textura de los suelos para ambas áreas estudiadas fue franco arenosa, la cual favorece el establecimiento y actividad de los hongos MA-VA.
• Con relación al pH de los suelos estudiados se encontró en un rango de 3.8 a 4.4, el cual influyó directamente sobre la diversidad y número de propágulos encontrados en la rizósfera.
• En cuanto a los niveles de fósforo disponible se encontraron niveles muy bajos en un rango de 0.2 a 5 p.p.m lo cual estimula la actividad simbiótica de las plantas con los hongos micorricicos.
RECOMENDACIONES
• Se recomienda continuar los estudios sobre hongos formadores de micorriza (MAVA) en ecosistemas de alta montaña con el fin de ampliar el conocimiento sobre. la diversidad microbiana de estos ecosistemas abordando nuevos enfoques de investigación. (Ecología, fisiología, dinámica poblacional, entre otros).
• Para evaluar en su totalidad la incidencia de estos hongos en la adaptabilidad de las especies en estudio, es necesario desarrollar investigaciones "Ex situ", con diseños experimentales que permitan cuantificar la efectividad de los hongos MA-VA y su actividad biológica en relación con las plantas.
• Consideramos de gran importancia desarrollar investigaciones especificas que busquen involucrar los hongos MA-VA como elemento bioindicador del estado de conservación de los ecosistemas presentes en un sitio determinado.
• Para los trabajos que se plantean sobre restauración ecológica se recomienda tener en cuenta los microorganismos que se asocian a la rizósfera en especial referencia las MA-VA dado el potencial y función que cumple la simbiosis al interior de la sucesión vegetal.
REFERENCIAS BIBLIOGRÁFÍCAS
AZCON, C. & BÁREA, J. 1996. Interacciones de las micorrizas arbusculares con microorganismos de la rizósfera. Departamento de Microbiología. Estación Experimental del Zaidín, CSIC. Granada, España. En: Guerrero, E. 1996. Micorrizas. Recurso biológico del suelo. Fondo FEN Colombia. 208 pp.
BORIE F. 1983. Fósforo orgánico y micorrizas "VA" en suelos volcánicos de Chile. Universidad de la Frontera, Temuco (Chile). En: Técnicas de investigación de micorrizas, Fundación Interamericana para la ciencia (FIC), Centro Agronómico Tropical de Investigación y Enseñanza.
CARRIZOSA, J. y HERNÁNDEZ J. 1990. Selva y futuro en Colombia. El sello editorial. Bogota.
COBA, B. Y COGUA 1995. Reconocimiento de MVA en el Páramo y Bosque Altoandino en la Región de Monserrate; En : Estudios Ecológicos del Páramo y del Bosque Altoandino, Cordillera Oriental de Colombia. p. 439.
CORREA RESTREPO, M. ; MORALES, M., COBA de G., B y E. SIEVERDING 1990. Presencia de Micorriza vesículo arbuscular (MVA) en un relicto de bosque andino, en la vereda " Alta Charco" departamento de Cundinamarca, Colombia. En : Resúmenes del V Congreso Latinoamericano de Botánica y I Simposio Latinoamericano de Micorrizas. Impreso Palacio de las comunicaciones. Cuba. 399 pp.
CORREA DE RESTREPO , M., MONTENEGRO , J., MARTÍNEZ, J., TORRES, E. y A. CAICEDO 1993. Análisis sobre la actividad de hongos formadores de micorrizas vesículo arbusculares. En : " Aspectos ambientales para el ordenamiento territorial del occidente del departamento del Caquetá ". Tomo II. Sección 4 :698-735. Instituto Geográfico Agustín Codazzi (IGAC). Ed.
GUERRERO, E.1993. Evaluación de las micorrizas en ecosistemas andinos del Parque Nacional Natural Chingaza primera etapa: Páramo. Bogota, Colombia. Departamento de Biología ,Facultad de Ciencias Pontificia Universidad Javeriana.
HERRERA, J. F., & F. E. RUIZ. 1981. Algunos aspectos de la ecología y de los efectos inmediatos del fuego sobre la antropofauna asociada a Espeletia grandiflora H & B. Páramo de Monserrate, Bogotá, Colombia. Tesis Departamento de Biología Universidad Nacional de Colombia. Bogotá.
SIEVERDING. E. 1991: Vesicular Arbuscular Mycorrhyza Management Tropical Agrosystems. Technical Cooperation Federal Republic of Germany. GTZ No. 224.
DEPARTAMENTO DE AGRICULTURA Y SUELOS DE LOS ESTADOS UNIDOS. 1999. Soil taxonomy.
UNIDAD ADMINISTRATIVA ESPECIAL DEL SISTEMA DE PARQUES NACIONALES NATURALES, (UAESPNN) MINISTERIO DEL MEDIO AMBIENTE. 2000. Plan Estratégico y de Manejo para el Parque Nacional Chingaza y su Zona de influencia 2000-2005. p. 10-35.
VALENCIA H, MURILLO, MOYANO. 1995. Micorrizas Vesículo-Arbusculares asociadas con tres especies nativas de páramo y bosque altoandino en la región de Monserrate, Colombia. En : Estudios Ecológicos del Páramo y del Bosque Altoandino, Cordillera Oriental de Colombia. p. 449.
Creation date:




Licencia
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






