
DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2004.1.a04Publicado:
01-01-2004Número:
Vol. 8 Núm. 17 (2004)Sección:
Artículos de investigación científica y tecnológicaEl banco de semillas germinable de especies leñosas en dos bosques subandinos y su importancia para la restauración ecológica (reserva biológica Cachalú - Santander. Colombia).
The germinable seed bank of woody species in two sub - Andean forests and its importance for ecological restoration (Cachalú biological reserve - Santander, Colombia)
Palabras clave:
Subandean forest, Ecological restoration, vegetable succession, Germinate seeds bank, Seeds dispersion, Seeds rain. (en).Palabras clave:
Bosque subandino, Restauración ecológica, Sucesión vegetal, Banco de semillas germinable, Dispersión, lluvia de semillas (es).Descargas
Referencias
BAKER, H. G. 1989. Some aspects of the natural history of seed banks. En: M.A.P. Leck, V.T. Parket; Simpson, R.L (eds.) Ecology of soil seed bank. San Diego, California. Academic Press; Inc., San Diego, California. pp. 9-21.
BARRERA, J. I.; RÍOS, H. F. 2002. Acercamiento a la ecología de la restauración. PerezArbelaezia. 13(2): 33-46.
BASKIN, J.M. & BASKIN, C.C. 1989. Physiology of dormancy and germination in relation to seed bank ecology. En: M.A.P. Leck, V.T. Parker & Simpson (eds.); Ecology of soil seed bank. Academic Press; Inc. San Diego, California, pp. 53-65.
BROKAW, N.V. 1986. Seed dispersal, gap colonization, and the case of Cecropia insignis. En: Frugivores and seed dispersal. Estrada, A. y Fleming (eds.), T. H. Panamá. Cap. 24: 323-331.
CARDONA, A. 2004. Potencial de regeneración del banco de semillas germinable en dos tipos de bosque subandino: implicaciones para la restauración ecológica Reserva Biológica Cachalú — Santander (Colombia). Tesis de Grado. Universidad Distrital "Francisco José de Caldas". Bogotá, Colombia.
FAHRIG, L. 2003. Effects ofhabitat fragmentation on biodiversity. Annu. Rey. Ecol. Evol. Syst. 34:487-515
FENNER, M. 1995. Ecology of seed banks. En: Kigel, J. & Galili, G. (eds). Seed development and germination. Inc. New York.
FUNDACIÓN NATURA. 2002. Plan de restauración ecológica en el corredor de bosques altoandinos de roble. Encino (Santander). Colombia. 45 p.
GARWOOD, N.C. 1989. Tropical soil seed banks: a review. En: M.A. Leck; V.T. Parker & R.L. Simpson (Eds); Ecology of soil seed bank. Academic Press; Inc. San Diego, California. pp. 149-209.
GENTRY, A. 2001. Patrones de diversidad y composición fiorística en los bosques de las montañas neotropicales. En: Maarten Kappelle & Alejandro Brown. (eds.) Bosques nublados del neotrópico. Instituto nacional de biodiversidad. INBIO. Santo Domingo de Heredia, Costa Rica. pp. 85-124.
GRIME, J.P. 1989. Seed bank in ecological perspective. En: M.A. Leck; V.T. Parker & Simpson (Eds). Ecololgy of soil seed bank. Academic Press; Inc. San Diego, California, pp. xv-xxii
GUARIGUATA, M.R. 2002. Pautas conceptuales y metodológicas para la planificación, ejecución y monitoreo de faenas de restauración ecológica. CATIE, Costa Rica.
HOLTHUIJZEN, A. M. & BOERBOOM, J. H. 1982. The Cecropia seed bank in the Surinan lowland raind forest. Biotropica. 14(1): 6268.
JAIMES, V. & RIVERA, D., 1990. Banco de semillas y tendencias en la regeneración natural de un bosque altoandino en la región de Monserrate (Cundinamarca, Colombia). Tesis de Grado. Universidad Nacional de Colombia. Bogota, Colombia.
LUDWIG, J. A. & REYNOLDS, J.F. 1988. Statistical ecology, a primer on methods and computing. John wiley & sons. Estados Unidos. pp.177.
MAGURRAN, A. E., 1989. Diversidad ecológica y su medición. ed. Vedra. Barcelona España. pp. 200.
MELI, P. 2003. Restauración ecológica de bosques tropicales. 20 años de investigación académica. Interciencia. 28(10): 581-589.
MONTENEGRO, A.L. 2000. Estrategias de dispersión y regeneración por banco de semillas en dos comunidades de bosque alto andino. Bogota, Colombia. Tesis de grado. Universiadad Nacional.
MORENO, C. E. 2001. Métodos para medir la biodiversidad. Centro de investigaciones biológicas, Universidad Autónoma del estado de Hidalgo. México. pp. 83.
QUINTANA-ASCENCIO, P.F., GONZÁLEZESPINOSA, M., RAMÍREZ-MARCIAL, N.; DOMÍNGUEZ-VASQUEZ, G. & MARTÍNEZ-ICÓ, M. 1996. Soil seed bank and regeneration of tropical rain forest from milpa field at the selva lacandona, Chiapas, Mexico. Biotropica, 28(8): 192-209.
RICO, V. G. & GARCÍA, J. F. 1986. Vegetation and soil seed bank of successional stages in tropical lowland deciduos forest. Journal of Vegetation Science. 3. pp. 617-624.
SIMPSON, R.L.; LECK, M. A.; PARKER, T. 1989. Seed bank: general concepts and methodological issues. En: M.A.P. Leck, V.T. Parker; R.L Simpson. (eds). Ecology of soil seed bank. San Diego, California. Academic Press; Inc. San Diego, California. pp. 3-8.
SORZANO, C. 2001. Regeneración reproductiva en un sitio abandonado e invadido por Pteridium aquilinum (L.) Khun. Implicaciones para la restauración. Reserva biológica Cachalú, EncinoSantander. Colombia. Tesis de grado. Pontificia Universidad Javeriana. Facultad de Ciencias. Bogotá, Colombia.
THOMPSON, K. 1987. Seed and seed banks. The New Phytologist. 106: pp. 23-34
OLAYA, A. 2000. Efecto de borde sobre el banco de semillas germinable del bosque altoandino (Embalse de San Rafael. La Calera, Cundinamarca). Tesis de grado. Pontificia Universidad Javeriana. Facultad de Ciencias. Bogotá, Colombia.
WIJDEVEN, S. M. & KUZZE, M.E. 2000. Seed availability as a limiting factor in forest recovery processes in Costa Rica. Restoration Ecology. 8 (4): 414-424
POSADA, C. & CARDENAS, C. 1999. Ecología de los bancos de semillas en una comunidad vegetal de páramo sometida a disturbios por quema y pastoreo. Tesis de grado. Universidad Nacional de Colombia. Bogota, Colombia.
ZIMMERMAN, J. K.; PASCARELLA, J. B.; AIDE, T. M. 2000. Barriers to forest regeneration in an abandoned pasture in Puerto Rico. Restoration Ecology. 8 (4): 350-360
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Artículo de Investigación científica y tecnológica
Colombia Forestal, Diciembre del 2004 vol:8 nro:17 pág:60-74
El Banco de semillas germinable de especies leñosas en dos bosques subandinos y su importancia para la restauración ecológica (Reserva Biológica Cachalú-Santander.Colombia).1
Angélica Cardona Cardozo
Ingeniero Forestal, Universidad Distrital "Francisco José de Caldas", Facultad del Medio Ambiente y Recursos Naturales. Proyecto Curricular de Ingeniería Forestal. Tel. (1)7794946. E-mail:angelicardonaca@yahoo.com
Orlando Vargas Ríos
Biólogo. Profesor Dpto. de Biología. Universidad Nacional de Colombia. jovargasr@unal.edu.co
Resumen
En dos bosques subandinos, en diferente estado sucesional, se evaluó la densidad y abundancia del Banco de Semillas Germinable (BSG) del suelo, para las especies leñosas. Se contabilizaron en total 1091 plántulas de especies leñosas, distribuidas en 18 familias, 22 géneros y 33 especies. El mayor número de plántulas se registró en el BSG del bosque secundario con 599 individuos, mientras que en las muestras del bosque maduro germinaron 492 individuos. Las especies con densidades mayores para el bosque maduro fueron Cestrum tomentosum, Cecropia telealba y Cecropia angustifolia, para el secundario las especies fueron Clethra fagifolia, Tibouchina lepidota y Cecropia telealba. No se encontraron diferencias significativas en el BSG de los dos bosques estudiados, en cuanto a la diversidad (t=0.05, g1=63, p=0.95), tamaño (t=0.05, g1=64, p=0.566) y composición; y los valores obtenidos por estos demuestran el alto grado de diversidad de los dos sitios estudiados. En comparación con otros bosques andinos el BSG encontrado es abundante y diverso.
Palabras clave: Bosque subandino. Restauración ecológica, Sucesión vegetal, Banco de semillas germinable, Dispersión, Lluvia de Semillas.
Abstract
In two subandean forests, in different state successional, was evaluated the density and abundance of the Germinate Seeds Bank (GSB) of the soil, for the woody species They were recorded in total 1091 seedlings of woody species, distributed in 18 families, 22 genus and 33 species. The greater number of seedlings was registered in the GSB of the secondary forest with 599 individual, while in the samples of the mature forest germinateed 492 individual. The species with greater densities for the mature forest were Cestrum tomentosum,. Cecropia telealba and Cecropia angustifolia, for the secondary the species were Clethra fagifolia, Tibouchina lepidota and Cecropia telealba. They were not found meaningful differences in the GSB of the two studied forests, concerning the diversity (t=0.05, g1=63, p=0.95), size (t=0.05, g1=64, p=0.566) and composition; and the values obtained by these demonstrate the high degree of diversity of the two studied sites. In comparison with other Andean forests the found BSG, is abundant and various.
Key Words: Subandean Forest, Ecological Restoration, Vegetable Succession,Germinate seeds bank, Seeds dispersion, Seeds rain.
INTRODUCCIÓN
En general, la fragmentación y pérdida de hábitats son las causas del remplazo de bosques nativos por ecosistemas antrópicos, lo cual produce parches remanentes segregados en una matriz de pasturas y/o cultivos (Fahrig, 2003), disminuyendo las posibilidades de restauración de estos hábitats, además ,de la consecuente pérdida de la biodiversidad y de la variabilidad genética de las poblaciones (Meli, 2003). Debido a esta situación cada vez más creciente, en los últimos arios se han dirigido esfuerzos hacia el desarrollo de la restauración ecológica, entendida según varios autores, como la practica con la cual un ecosistema degradado, dañado o destruido, es asistido en el proceso de sucesión ecológica, hasta alcanzar nuevamente parte de su estructura y función, utilizando como marco de referencia el ecosistema original o predisturbio y la teoría ecológica (Sorzano, 2001; Barrera & Ríos, 2002; Guariguata, 2002; Meli, 2003).
El BSG, se define como un reservorio de semillas en el suelo, no germinadas y que representa diversas especies, genotipos y fenotipos, las cuales son potencialmente capaces de remplazar plantas adultas, una vez dadas las condiciones de humedad, luz y temperatura para su germinación; de esta manera, la vegetación inicial después de un disturbio, estará determinada por la composición del banco de semillas, que puede ser muy diferente de la vegetación predisturbio (Baker, 1989; Garwood, 1989), no obstante, la composición y abundancia de las especies dentro del banco de semillas varia en espacio y tiempo, debido en gran parte a los modelos de dispersión de semillas y emergencia de plántulas (Brokaw, 1986). Esta reserva de semillas viables y dormantes, asegura que las especies van a estar en el lugar y tiempo indicado cuando se presente la oportunidad para su germinación (Simpson et al., 1989; Grime, 1989), dando inicio al proceso de sucesión vegetal y proporcionando una herramienta para la restauración ecológica; además los bancos de semillas juegan un papel importante en el mantenimiento de la vegetación, creación de hábitats y manejo diferencial de especies (Posada & Cárdenas, 1999).
Aunque desde hace algunos arios se sabe que los suelos de las regiones tropicales contienen un considerable número de semillas viables de especies secundarias (Holthuüzen y Boerboom, 1982; Garwood, 1989), son pocos los estudios que se han realizado con un enfoque hacia la restauración ecológica, tan solo un 4% de estos estudios se desarrollan en países como India, Venezuela, Ecuador, Argentina, Nigeria, República Dominicana y Colombia (Meli, 2003). En Colombia los estudios del BSG se han centrado en bosques altoandinos y el páramo, debido a su fragilidad y a la necesidad de acciones rápidas para su preservación, dando a conocer la existencia e importancia del BSG en este tipo de ecosistemas (Jaimes & Rivera, 1990; Posada & Cárdenas, 1999; Olaya 2000; Montenegro, 2000; Sorzano, 2001;), sin embargo, es necesario investigar el BSG en otras comunidades vegetales y su aplicación en el proceso de restauración ecológica (Garvvood, 1989).
La presente investigación hace parte del primer estudio referente al BSG de bosques subandinos en el país y es un aporte a la ecología de dicha comunidad vegetal, permitiendo identificar las especies presentes en el BSG y contribuir de esta manera en el entendimiento de la dinámica de la vegetación del bosque. Conocimiento que puede ser útil en prácticas de uso sostenible y de ordenación de bosques subandinos tropicales (a través del manejo de la regeneración natural), además de la restauración ecológica.
OBJETIVO GENERAL
Caracterizar el banco de semillas germinable (BSG) de las especies leñosas, en comunidades de bosque subandino, identificando el potencial de estas especies en procesos de restauración ecológica.
OBJETIVOS ESPECÍFICOS
• Identificar y comparar las especies arbóreas y arbustivas presentes en el BSG del bosque maduro y del bosque secundario.
• Determinar la densidad y diversidad en cuanto a las especies arbóreas y arbustivas presentes en el BSG de los dos tipos de bosques estudiados.
• Evaluar el potencial de las especies arbóreas y arbustivas encontradas en el BSG, para ser utilizadas en procesos de restauración ecológica en comunidades vegetales subandinas.
METODOLOGIA
SITIOS DE MUESTREO
La Reserva Biológica Cachalú, donde se realizó la presente investigación, es una figura de conservación perteneciente al Sistema Nacional de Áreas Protegidas, clasificada como Reserva de la Sociedad Civil y administrada por la Fundación Natura. Hace parte de una Ecorregión Nacional según política ambiental y se encuentra ubicada dentro de la zona de amortiguación del Santuario de Fauna y Flora Guanentá - Alto Río Fonce.
Bosque Maduro: Se seleccionó un bosque ubicado aproximadamente a 5 kilómetros desde el centro de investigaciones de la Reserva, a los 06° latitud N y 73° longitud W y a una altitud de 1.700 m.s.n.m. Este sitio se encuentra en el límite entre la Reserva Biológica Cachalú y el Santuario de Fauna y Flora Guanentá-Alto Río Fonce. Se escogió este bosque como maduro por presentar características florísticas que así lo evidencian, como el tipo de vegetación, altura y DAP de los individuos, además del testimonio de habitantes de la región que aseguran no haberse presentado extracción maderera en este lugar debido a lo distante y a las características del terreno.
Bosque Secundario: Corresponde a un bosque ubicado a 15 m. al lado izquierdo de la Torre de observación de la Reserva, entre los 06° latitud N y 74° longitud W y a una altitud de 1.860 m.s.n.m. Este es un bosque secundario tardío que ha estado en regeneración por más de veinte años, después de ser intervenido mediante entresaca selectiva de maderas finas, según investigaciones realizadas entre los habitantes del lugar, por la Fundación Natura (2002).
BANCO DE SEMILLAS GERMINABLE
En cada tipo de bosque a evaluar se tomaron muestras de suelo a lo largo de cuatro (4) transectos de 10 m. de longitud, los cuales se localizaron siguiendo una figura en zig-zag, con un ángulo de 45°. A partir del vértice de los transectos, cada tres (3) metros se localizaron puntos de muestreo, en los cuales se colectaron tres (3) subunidades muestrales de 10 cm de profundidad, cercana una de otra, estas tres subunidades se unieron conformando una única unidad muestral de 3.981 cm3 (Cardona, 2004) para cada punto de muestreo. Para la medición exacta del volumen de suelo en campo, se empleó un cilindro metálico de 10 cm. de altura y 13 cm. de diámetro, coh una capacidad de 1.327 cm3 con el que se colectaron las subunidades muestrales. Al momento de colectar las muestras se retiró la capa de hojarasca (Horizonte 0) y debido al gran volumen de raíces sin descomponer, se tamizo el suelo a través de un tamiz de ojo de malla de 5 mm.
El volumen de suelo por unidad muestral se transportó a un invernadero, localizado contiguo al centro de investigaciones de la Reserva, con las condiciones ambientales adecuadas de temperatura, luz y humedad relativa para la germinación de las semillas. Al interior del invernadero se adecuaron mesones cubiertos con velo blanco, para evitar contaminación por lluvia de semillas y permitir la entrada normal de luz, en estos mesones se ubicaron las bandejas de germinación con el suelo muestreado y entre estas se distribuyeron 20 bandejas de control con solo arena, para detectar contaminación por lluvia de semillas dentro del invernadero.
En cada bandeja de germinación se utilizó un sustrato de 2 cm. de arena lavada de río, previamente esterilizada con agua hirviendo; el suelo se colocó sobre este sustrato en cada bandeja, en una capa no mayor a 1 cm, para facilitar la germinación de las semillas (Posada y Cárdenas, 1999). Las bandejas se regaron según el requerimiento, para mantener una humedad constante y suficiente para la germinación.
Semanalmente se contaron y marcaron las plántulas germinadas con banderitas rotuladas, según la fecha de germinación. Inicialmente las plántulas se clasificaron a nivel de morfoespecie, algunas de estas se transplantaron con el fin de identificarlas posteriormente a nivel de familia, género y especie» Concluidos 6 meses no se presentó la germinación de ningún individuo, dando por terminado el registro. Las especies de dificil identificación, se buscaron en estado adultos en campo y se compararon con especímenes de vivero para su identificación.
• Composición del BSG
La composición del BSG se evaluó a través de la abundancia y la densidad, estas dos medidas estian la diversidad y permiten hacer un diagnóstico confiable. La densidad se halló dividiendo la abundancia absoluta de las especies por el área de la unidad muestreal, que se define como 3(3.1416)m 2, ya que se extrajo suelo tres veces con el mismo cilindro, para l;a obtención de la muestra
Lo cual da un área de 0.04 m 2 para cada unidad muestral y de 0.48 m 2 en total por bosque.
• Diversidad del BSG
Para el análisis de alfa diversidad se utilizaron los índices de riqueza y abundancia, referidos en la Tabla 1 (Magurran, 1989). La diversidad beta se calculó a través del Coeficiente de Sorensen y el cuantitativo de Sorensen, descritos en la Tabla 2 (Magurran, 1989; Moreno, 2001).
Se realizaron análisis con la prueba de t-Student, para determinar diferencias entre sitios con respecto a la abundancia y densidad de especies, y los índices de diversidad de Simpson y Shannon. Se realizó una prueba de chi-cuadrado .X2 utilizando la corrección de Yates' s, para determinar diferencias entre sitios a partir de las abundancias de las especies dominantes. La corrección se utilizó debido a que las diferencias se hallaron entre pares de especies dominantes (g1=2), lo que genera que el resultado del test estadístico en estos casos sea sesgado (Ludwig y Reynolds, 1988).
RESULTADOS
BANCO DE SEMILLAS GERMINABLE
• Composición del BSG En total en el banco de semillas de los dos sitios muestreados germinaron 1091 plántulas de especies leñosas, pertenecientes al 8 familias, distribuidas en 22; géneros y 33 especies. En el Banco de Banco de Semillas Germinable Germinable (BSG) del bosque secundario se registraron 599 individuos de especies leñosas, un poco mas que en el bosque maduro donde se registaron 492 individuos (Tabla 3). Germinaron 19 especies de tipo arbóreo y 13 de tipo arbustivo, iguales para los dos estados sucesionales, y una especie arbustiva que únicamente se registró en el BSG del bosque secundario.



La familias de las especies encontradas en el BSG, presentaron igual número de especies en los dos estados sucesionales, a excepción de Asteraceae que registró una especie más en el BSG del bosque secundario. La familia con mayor riqueza fue Solaneceae con 5 especies, seguida por Cecropiaceae y Asteraceae con 3 especies cada una; Clusiaceae, Melastomataceae, Moraceae, Piperaceae, Rubiaceae y Myrtaceae, registraron dos especies cada una. En general con los datos de los dos estados sucesionales, las familias más dominantes fueron Cecropiaceae con 280 individuos, Solanaceae con 221 individuos, Clethraceae con 113 individuos, Melastomataceae con 109 individuos y Rubiaceae con 51 individuos. Para el BSG del bosque maduro las familias más dominantes fueron en orden descendente Cecropiaceae, Solanaceae, Rubiaceae y Melastomataceae; este orden varió un poco en el BSG del bosque secundario donde después de Cecropiaceae, le siguen Clethraceae, Solanaceae y Melastomataceae (Figura 1).
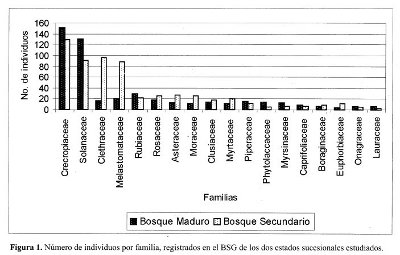
Entre las especies arbóreas, las tres del género Cecropia, (Cecropia angustifolia, Cecropia telealba, Cecropia telenitida) están dentro de las primeras cinco más abundantes en los dos bosques. Para el bosque maduro además son importantes en términos de su abundancia Guettarda sabiceoides que ocupa el tercer lugar y del quinto en adelante se encuentran en

orden descendente Clethra fagifolia (Figura 2), Myrsine coriaceae, Tibouchina lepidota (Figura 3), Vismia baccifera, Viburnum cornifolium, Solanum sp, Piper sp y Ficus sp (Tabla 4). En el bosque secundario se encuentran Clethra fagifolia y Tibouchina lepidota, ocupando el primero y segundo lugar respectivamente, seguidas de las tres especies de Cecropia, del quinto lugar en adelante
están en orden descendente Ficus sp, Solanum sp, Croton smithianus, Cordia cilindrostachya, Vismia baccifera, Vismia guianensis, Guettarda sabiceoides, Viburnum cornifolium y Myrsine coriaceae.
Dentro de los arbustos las especies más dominantes en el BSG, para el bosque maduro fueron en orden descendente Cestrum tomentosum, Rubus sp, Phytolacca dioica, Psidium guineensis, Ludwigia peruviana, Palicourea sp, Solanum quitoensis, Eupatorium sp y Psidium guyanense. Para el bosque secundario las especies fueron Cestrum tomentosum, Rubus sp, Palicourea sp, Solanum quitoensis , Psidium guineensis , Baccharis trinervis, Baccharis lehemannii, Psidium guyanense, Baccharis decussata, Ludwigia peruviana y Phytolacca dioica (Tabla 4).
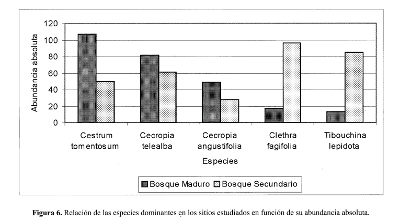
La densidad para el bosque maduro y el secundario fue de 1.025 semillas/m 2 y 1.248 semillas/m2 respectivamente. Las especies con densidades mayores en el bosque maduro fueron, Cestrum tomentosum (223 semillas/m2), Cecropia telealba (171 semillas/m2) y Cecropia angustifolia (102 semillas/m2 ) (Fig. 4); para el secundario las especies fueron Clethra fagifolia (200 semillas/m2), Tibouchina lepidota (177 semillas/m2) y Cecropia telealba (127 semillas/m2) (Fig. 5).

El análisis estadístico de los datos, ratificó que no existen diferencias significativas para los parámetros de abundancia (t=0.05, gl= 64, p=0.566) y densidad (t=0.05, g1=64, p=0.566) entre las muestras de los dos sitios estudiados. La prueba de chi-cuadrado .X2, con la corrección de Yates' s, arrojó que las especies más abundantes presentan diferencias significativas entre los dos sitios de estudio, esto corrobora la dominancia de Cestrum tomentosum (X 2 = 37.58, g1=1, p=0.05), Cecropia telealba (X2 = 9.57, g1=1, p=0.05) y Cecropia angustifolia (X2 = 10.92, g1=1, p=0.05) en el BSG del bosque maduro, y de Clethra fagifolia (X2 = 44.48, g1=1, p=0.05) y Tibouchina lepidota (X2 = 43.03, g1=1, p=0.05) en el BSG del bosque secundario (Figura 6).
Diversidad del BSG
• Índices de diversidad alfa
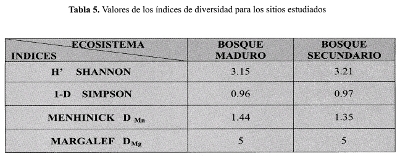
El índice de Shannon-Wiener indica que el sitio más diverso en cuanto a las especies de tipo leñoso,

es el bosque secundario; sin embargo, la diferencia con el bosque maduro es muy escasa, de tan solo 0.06 bits. El índice de Simpson, no presenta ninguna diferencia entre los dos bosques (Tabla 5), aunque su valor, muy cercano a uno (1) demuestra el alto grado de dominancia de las especies en los dos sitios estudiados. Según el índice de Margalef, los dos bosques son igualmente ricos en especies, debido principalmente a que no hay diferencia en el número de especies encontradas en los BSG de los sitios. Por otro lado el índice de Mehinick, indica que el sitio más diverso en cuanto a riqueza de especies es el bosque maduro, aunque por una diferencia no muy grande con el bosque secundario.
Según prueba estadística para la diversidad, medida a través de los índices de Simpson (t=0.05, g1=50, p=0.70) y Shannon (t=0.05, g1=63, p=0.95), no se encontraron diferencias significativas para los dos bosques estudiados.
• Índices de diversidad beta
La beta diversidad medida a través del coeficiente de Sorensen, calculado según número de especies comunes para los dos bosques, dio como resúltado 0.98, señalando de igual manera que los dos sitios son muy similares en cuanto a la composición de especies del BSG. Por otro lado el cuantitativo de Sorensen dio como resultado 1.99, lo que indica un grado de diversidad beta muy alto, esto se debe a que este índice es calculado con base en datos cuantitativos, en este caso de abundancia de especies del BSG, la cual presenta una mayor diferencia entre sitios, en comparación con la riqueza de especies; lo anterior permite deducir para este estudio que el BSG de los sitios estudiados cambia en menor grado en cuanto a su abundancia pero no en cuanto a la composición de especies a través de los dos estados sucesionales.
DISCUSIÓN DE RESULTADOS
El banco de semillas de especies leñosas encontrado en los dos bosques estudiados es muy rico y abundante y corrobora lo expuesto por Grime (1989), quien afirma que inclusive dentro de hábitats con una vegetación perenne cerrada, hay presencia de un número substancial de semillas dormantes en el suelo, con características que impiden su germinación y les permiten subsistir allí. De igual manera, Se puede considerar que el BSG encontrado en los dos estados sucesionales, requiere en general de los mismos factores que estimulan la germinación de semillas, descritos por muchos autores (Grime, 1989; Baskin & Baskin, 1989; Fenner, 1995), ya que las semillas presentes en el suelo muestreado, responden favorablemente a la mayor incidencia de luz, a la fluctuación en la temperatura y al cambio en la humedad relativa, rompiendo la dormancia condicional y permitiendo la germinación (Thompson, 1987); muchas de estas semillas pueden permanecer largos periodos de tiempo en el suelo y germinar bajo condiciones favorables para la especie.
TAMAÑO DEL BANCO DE SEMILLAS GERMINABLE
Comparando la abundancia de las especies leñosas encontradas en el BSG de los dos bosques estudiados, no se presentó una variación significativa en el tamaño y composición, como se esperaba debido a su diferencia sucesional, ya que varios estudios sugieren que el BSG de los bosques maduros es mucho mas pequeño y menos diverso, que los de comunidades sucesionales (Rico & García, 1986). Una de las razones del tamaño del BSG del bosque maduro, puede ser la frecuente apertura de claros que este presenta; ya que con la apertura del dosel se promueve la entrada de semillas desde sitios distantes, gracias a la dispersión por anemocoria principalmente, aunque un número considerable de semillas también puede llegar por zoocoria; estudios han encontrado que algunas especies dispersadas por el viento tenían más semillas por unidad de área en los claros, que en el bosque cerrado (Brokaw, 1986). De esta manera, la influencia de la dinámica de claros y los vectores de dispersión que actúan en el bosque maduro, permiten la entrada de semillas de manera diversa, debido a la dispersión por zoocoria en el bosque en general y anemocoria a través de la dinámica de claros.
La riqueza de especies leñosas encontradas en el BSG de este estudio, es bastante grande comparada con otros estudios realizados en comunidades tropicales a mayor altitud, donde las especies de tipo arbóreo o arbustivo suman menos del 30% del total de las especies germinadas (Jaimes & Rivera, 1990; Posada & Cárdenas, 1999; Olaya 2000; Montenegro, 2000; Sorzano, 2001); sin embargo, esta característica parece revertirse en comunidades a menor altitud, como lo encontrado por un estudio realizado en Costa Rica a 1100 m de altitud en un bosque de sucesión avanzada, donde se halló que el 26% de las especies encontradas en el BSG eran árboles y en total el 86% pertenecían a especies leñosas (Brokaw, 1986).
Dentro de las especies arbóreas germinadas en el banco de semillas de los dos sitios estudiados, el género Cecropia fue el que mayor número de especies presentó, con Cecropia angustifolia, Cecropia telealba y Cecropia telenitida, además estas fueron las especies más abundantes en el BSG del bosque maduro y se encontraron entre las de mayor abundancia en el bosque secundario; lo cual hace que estas especies sean unas de las pioneras más importantes en este tipo de comunidad. Un estudio realizado en Surinam sobre el BSG de las especies del género Cecropia, encontró un total de 73 semillas/m2 de este género, un número mucho menor que el encontrado en el presente estudio, donde se hallaron 315 semillas/m 2 en el bosque maduro y 268 semillas/m2 en el bosque secundario, adicionalmente, el mismo estudio encontró que la abundancia de especies de este género varía durante el ario, encontrando que la menor densidad se presenta al final de la primera época de lluvias en julio (33 semillas/m2) y la mayor al principio del segundo periodo de lluvias en diciembre (127 semillas/m2), en cuanto a la viabilidad, reportaron que las semillas de C. sciadophylla y C. obtusa, pierden viabilidad después de 62 y 48 meses de estar enterradas respectivamente (Holthuijzen y Boerboom, 1982); otro estudio realizado en Panamá señala que la dispersión de Cecropia insignis es realizada por un gran un número de animales dispersores de aproximadamente 100 géneros, y que el 58% de las trampas utilizadas para medir dispersión recibieron semillas de esta especie durante 4 meses de muestreo, obteniéndose un porcentaje grande de individuos de este género (Brokaw, 1986).
Los resultados obtenidos por todos los estudios, como los señalados anteriormente, demuestran que existe un considerable número de semillas dormantes de este género a lo largo del ario y aunque la entrada de semillas al banco de semillas puede variar dependiendo del pico de fructificación durante el ario, la dispersión nunca es nula. En virtud a esa persistente dispersión algunos estudios sugieren, que la floración de estas especies puede ocurrir en cualquier época del ario, debido a que son polinizadas por el viento, lo que generaría que la producción de semillas sea continua y abundante, permitiendo que las especies de este género sean pioneras eficientes de gran importancia en la recuperación de bosques tropicales.
COMPOSICIÓN Y DIVERSIDAD DEL BANCO DE SEMILLAS GERMINABLE
La similitud en la composición de especies del BSG de los dos estados sucesionales estudiados, puede estar influenciada por la sucesión avanzada del bosque secundario, aunque presente un dosel menos cerrado y una densidad y altura de individuos menor a la del bosque maduro, su estructura vegetal, si puede influir significativamente en el tipo y cantidad de dispersión, de esta manera el bosque secundario recibe las mismas especies dispersadas por zoocoria que entran al banco de semillas del bosque maduro, originando que la riqueza en los dos estados sucesionales sea similar.
Por otro lado, la afinidad entre la abundancia de semillas del BSG y la edad de la vegetación en pie, es afectada por los atributos sucesionales de los parches de vegetación que los rodean y por la complejidad regional (Quintana-Ascencio et al. 1996), que para el caso del bosque maduro, ésta es predominantemente de bosques secundarios y pastizales, lo que permitiría la dispersión y entrada al BSG de gran cantidad de semillas de especies pioneras como es el caso de Cestrum tomentosum , Cecropia angustifolia y Cecropia telealba, que son las especies dominantes en el BSG de este bosque, haciendo que la composición y tamaño del BSG del bosque maduro sea muy similar a la del bosque secundario.
Asimismo las semillas que entran al bosque maduro a través de los claros y no germinan, pueden ser redispersadas dentro del bosque por gravedad y escorrentía, debido principalmente a la topografía con fuertes pendientes en los alrededores del sitio de muestreo, que harían posible la concentración de un gran número de semillas en este lugar. Este puede ser el caso de especies como Clethra fagifolia, Tibouchina lepidota, Tibouchina Miconiaminutiflora, Myrsine coriaceae, Baccharis lehemannii, Eupatorium sp y Baccharis trinervis, que son dispersadas por el viento y se encontraron en el BSG del bosque maduro, lo cual de algún modo también podría explicar su baja abundancia en este bosque, debido a que la presencia allí, estaría condicionada por la ocurrencia de claros y la redispersión de las semillas. Lo contrario ocurre en el bosque secundario donde las especies dispersadas por el viento son las mas dominantes como es el caso de Clethra fagifolia y Tibouchina lepidota, debido a que las condiciones del lugar y la estructura de la vegetación favorecen este tipo de dispersión.
IMPLICACIONES DE LOS BSG EN LA RESTAURACIÓN ECOLÓGICA
Son muchas las implicaciones que tienen los BSG en la restauración ecológica, ya que las especies contenidas en estos son pioneras capaces de colonizar sitios degradados e iniciar el proceso de sucesión. Para el caso del BSG de los bosques estudiados, este representa un potencial genético de especies leñosas, que la mayoría de los sitios disturbados han perdido y que difícilmente recuperarían de manera natural (Zimmerman et al., 2000).
Estos BSG pueden ayudar principalmente en sitios donde los factores limitantes de tipo biológico, causan retardo en la velocidad del proceso sucesional, tales factores hacen referencia a situaciones donde hay falta de semillas dispersadas naturalmente, alta predación de semillas, defoliación por insectos e invasión de malezas (Wijdeven y Kuzze, 2000; Guariguata, 2002; Meli, 2003), en estos casos, técnicas como: i) la donación de suelo de sitios mejor conservados, tales como bosques maduros y de sucesión avanzada, que poseen especies pioneras, tal como se constató en esta investigación; ii) el manejo de especies que forman BSG, lo cual además facilitaría la recolección y germinación de estas especies, para ser utilizadas posteriormente en procesos de reforestación y iii) el direccionamiento de la sucesión a partir del BSG; pueden ayudar en la recuperación de la vegetación de acuerdo con la intensidad, magnitud, duración y frecuencia del disturbio previo y así obtener resultados con menos costos y en menor tiempo.
De acuerdo con el alto grado de diversidad encontrado en el BSG de los dos estados sucesionales estudiados, estos tendrían el suficiente potencial genético para recuperar la diversidad perdida de sitios degradados en este tipo de ecosistema, en relativamente poco tiempo a través de la sucesión vegetal; sin embargo, la composición de especies de un bosque maduro es mucho mas dificil de recuperar y el papel del BSG en este caso seria el de modificar las condiciones del medio con el fin de facilitar el arribo de las especies que hacían parte del ecosistema antes del disturbio.
No obstante es necesario investigar sobre las necesidades de establecimiento de las especies encontradas en el BSG, ya que difieren de las necesidades de germinación; y en últimas la capacidad para establecerse bajo ciertas condiciones en un hábitat es lo que evalúa la eficiencia de la especie como agente importante en la restauración ecológica. Algunas de las especies del BSG, por ejemplo, podrían establecerse más fácilmente si son germinadas en invernadero y posteriormente transplantada, que si son puestas a germinar directamente en el sitio a restaurar; o su capacidad de establecerse podría también estar muy ligada a las condiciones ambientales del sitio, por lo que es necesario saber cual es el mejor periodo en el año para el desarrollo de cierta especies. Por ejemplo, en un ensayo acerca del establecimiento de Cecropia schreberiana, se demostró que la remoción de la vegetación herbácea no mejora la germinación de esta especie encontrada en el BSG, según el autor esto podría deberse a una alta predación por hormigas y otros predadores; otro experimento con semillas de esta especie puestas en campo, dio como resultado la remoción de todas las semillas en menos de ocho horas (Zimmerman et al, 2000).
En conclusión las semillas del género Cecropia según los resultados obtenidos por diversos estudios, poseen mecanismos de dormancia y la habilidad para detectar y responder a la germinación relacionada con la probabilidad de establecimiento, esto sumado a un largo periodo de fructificación, una abundante producción de semillas pequeñas y un amplio rango de dispersión gracias a la ayuda de numerosos agentes dispersores, hacen que las especies de este género sean las más abundantes dentro del BSG como lo encontrado en este estudio y por lo tanto un efectivo colonizador en comunidades de tierras tropicales bajas después del disturbio del bosque, por otro lado estas especies son de rápido crecimiento y llegan a hacer parte del dosel en pocos arios. Otras de las especies encontradas en el BSG, importantes por su capacidad de colonización son Tibouchina lepidota, Clethra fagifolia, Croton smithianus, Vismia guianensis, Myrsine coriaceae, Vismia baccifera y Viburnurn cornifolium, por que aunque no poseen todas las características presentes en el género Cecropia, si exhiben una amplia dispersión, debido principalmente a que tienen una alta producción de semillas pequeñas.
CONCLUSIONES
• El banco de semillas germinable de especies leñosas encontrado en los dos bosques estudiados es bastante rico y abundante en cuanto a su densidad y diversidad.
• Las semillas presentes en el suelo muestreado, respondieron favorablemente a los cambios en las condiciones ambientales, lo cual es un indicio de que muchas de estas semillas poseen dormancia condicional.
• No se halló una variación significativa en cuanto al tamaño, composición y diversidad del BSG de los dos bosques estudiados, como se esperaba debido a la diferencia sucesional.
• La densidad del BSG hallada en el bosque maduro es superior a la reportada por otros estudios para los bosques maduros tropicales; esto podría estar relacionado con la constante apertura de claros, permitiendo la entrada de semillas de partes distantes, principalmente por anemócoria, dando como resultado el aumento en el tamaño del BSG y por consiguiente la similitud entre los dos estados sucesionales.
• Según el alto grado de diversidad en cuanto a especies leñosas de los BSG estudiados, estos tendrían el suficiente potencial genético para ayudar en la recuperación de sitios degradados, en relativamente poco tiempo a través de la sucesión vegetal, sin embargo este proceso se podría acelerar con el manejo del BSG.
• Las especies del género Cecropia, son las pioneras más eficaces en este tipo de ecosistema según la abundancia y la capacidad de respuesta a los cambios ambientales.
AGRADECIMIENTOS
Los autores agradecen a la Fundación Natura por la financiación de este proyecto y la facilitación para el desarrollo del mismo. A los encargados del Herbario de la Universidad Distrital Francisco José de Caldas, por permitir la revisión de la colección de las muestras botánicas.
Agradecimientos especiales a Carolina Roa,a por la colaboración para la realización de esta investigación; al Ingeniero Forestal Gilberto Mahecha, por la indispensable ayuda en la determinación de las muestras botánicas. A José, Javier yr Omaira por su valioso apoyo en la fase de campo.
Y en general a todas aquellas instituciones y personas que tuvieron que ver directa o indirectamente en el desarrollo de la investigación.
REFERENCIAS BIBLIOGRÁFICAS
BAKER, H. G. 1989. Some aspects of the natural history of seed banks. En: M.A.P. Leck, V.T. Parket; Simpson, R.L (eds.) Ecology of soil seed bank. San Diego, California. Academic Press; Inc., San Diego, California. pp. 9-21.
BARRERA, J. I.; RÍOS, H. F. 2002. Acercamiento a la ecología de la restauración. PerezArbelaezia. 13(2): 33-46.
BASKIN, J.M. & BASKIN, C.C. 1989. Physiology of dormancy and germination in relation to seed bank ecology. En: M.A.P. Leck, V.T. Parker & Simpson (eds.); Ecology of soil seed bank. Academic Press; Inc. San Diego, California, pp. 53-65.
BROKAW, N.V. 1986. Seed dispersal, gap colonization, and the case of Cecropia insignis. En: Frugivores and seed dispersal. Estrada, A. y Fleming (eds.), T. H. Panamá. Cap. 24: 323-331.
CARDONA, A. 2004. Potencial de regeneración del banco de semillas germinable en dos tipos de bosque subandino: implicaciones para la restauración ecológica Reserva Biológica Cachalú — Santander (Colombia). Tesis de Grado. Universidad Distrital "Francisco José de Caldas". Bogotá, Colombia.
FAHRIG, L. 2003. Effects ofhabitat fragmentation on biodiversity. Annu. Rey. Ecol. Evol. Syst. 34:487-515
FENNER, M. 1995. Ecology of seed banks. En: Kigel, J. & Galili, G. (eds). Seed development and germination. Inc. New York.
FUNDACIÓN NATURA. 2002. Plan de restauración ecológica en el corredor de bosques altoandinos de roble. Encino (Santander). Colombia. 45 p.
GARWOOD, N.C. 1989. Tropical soil seed banks: a review. En: M.A. Leck; V.T. Parker & R.L. Simpson (Eds); Ecology of soil seed bank. Academic Press; Inc. San Diego, California. pp. 149-209.
GENTRY, A. 2001. Patrones de diversidad y composición fiorística en los bosques de las montañas neotropicales. En: Maarten Kappelle & Alejandro Brown. (eds.) Bosques nublados del neotrópico. Instituto nacional de biodiversidad. INBIO. Santo Domingo de Heredia, Costa Rica. pp. 85-124.
GRIME, J.P. 1989. Seed bank in ecological perspective. En: M.A. Leck; V.T. Parker & Simpson (Eds). Ecololgy of soil seed bank. Academic Press; Inc. San Diego, California, pp. xv-xxii
GUARIGUATA, M.R. 2002. Pautas conceptuales y metodológicas para la planificación, ejecución y monitoreo de faenas de restauración ecológica. CATIE, Costa Rica.
HOLTHUIJZEN, A. M. & BOERBOOM, J. H. 1982. The Cecropia seed bank in the Surinan lowland raind forest. Biotropica. 14(1): 6268.
JAIMES, V. & RIVERA, D., 1990. Banco de semillas y tendencias en la regeneración natural de un bosque altoandino en la región de Monserrate (Cundinamarca, Colombia). Tesis de Grado. Universidad Nacional de Colombia. Bogota, Colombia.
LUDWIG, J. A. & REYNOLDS, J.F. 1988. Statistical ecology, a primer on methods and computing. John wiley & sons. Estados Unidos. pp.177.
MAGURRAN, A. E., 1989. Diversidad ecológica y su medición. ed. Vedra. Barcelona España. pp. 200.
MELI, P. 2003. Restauración ecológica de bosques tropicales. 20 años de investigación académica. Interciencia. 28(10): 581-589.
MONTENEGRO, A.L. 2000. Estrategias de dispersión y regeneración por banco de semillas en dos comunidades de bosque alto andino. Bogota, Colombia. Tesis de grado. Universiadad Nacional.
MORENO, C. E. 2001. Métodos para medir la biodiversidad. Centro de investigaciones biológicas, Universidad Autónoma del estado de Hidalgo. México. pp. 83.
QUINTANA-ASCENCIO, P.F., GONZÁLEZESPINOSA, M., RAMÍREZ-MARCIAL, N.; DOMÍNGUEZ-VASQUEZ, G. & MARTÍNEZ-ICÓ, M. 1996. Soil seed bank and regeneration of tropical rain forest from milpa field at the selva lacandona, Chiapas, Mexico. Biotropica, 28(8): 192-209.
RICO, V. G. & GARCÍA, J. F. 1986. Vegetation and soil seed bank of successional stages in tropical lowland deciduos forest. Journal of Vegetation Science. 3. pp. 617-624.
SIMPSON, R.L.; LECK, M. A.; PARKER, T. 1989. Seed bank: general concepts and methodological issues. En: M.A.P. Leck, V.T. Parker; R.L Simpson. (eds). Ecology of soil seed bank. San Diego, California. Academic Press; Inc. San Diego, California. pp. 3-8.
SORZANO, C. 2001. Regeneración reproductiva en un sitio abandonado e invadido por Pteridium aquilinum (L.) Khun. Implicaciones para la restauración. Reserva biológica Cachalú, EncinoSantander. Colombia. Tesis de grado. Pontificia Universidad Javeriana. Facultad de Ciencias. Bogotá, Colombia.
THOMPSON, K. 1987. Seed and seed banks. The New Phytologist. 106: pp. 23-34
OLAYA, A. 2000. Efecto de borde sobre el banco de semillas germinable del bosque altoandino (Embalse de San Rafael. La Calera, Cundinamarca). Tesis de grado. Pontificia Universidad Javeriana. Facultad de Ciencias. Bogotá, Colombia.
WIJDEVEN, S. M. & KUZZE, M.E. 2000. Seed availability as a limiting factor in forest recovery processes in Costa Rica. Restoration Ecology. 8 (4): 414-424
POSADA, C. & CARDENAS, C. 1999. Ecología de los bancos de semillas en una comunidad vegetal de páramo sometida a disturbios por quema y pastoreo. Tesis de grado. Universidad Nacional de Colombia. Bogota, Colombia.
ZIMMERMAN, J. K.; PASCARELLA, J. B.; AIDE, T. M. 2000. Barriers to forest regeneration in an abandoned pasture in Puerto Rico. Restoration Ecology. 8 (4): 350-360
Notas
1 Investigación, financiada por la Fundación Natura.
Creation date:




Licencia
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






