DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2005.1.a06Published:
2005-01-01Issue:
Vol. 9 No. 18 (2005)Section:
Scientific articleCaracterización florística y estructural de unidades de bosque altoandino en las Veredas Minas y Patios Altos, Enciso - Santander
Floristic and structural characterization of high Andean forest units in the Veredas Minas and Patios Altos, Enciso - Santander
Keywords:
bosque altoandino, composición florística, estructura, diversidad, Encino, Santander (es).Keywords:
Altoandino forest, floristic compositions, structure, diversity, Encino, Santander (en).Downloads
References
ANDRADE, G. 1993. Paisaje y biodiversidad en las selvas de los Andes. Proyecto Reserva Biológica Carpanta. Fundación Natura, EAAB & INDERENA. Bogotá, D.C.
BRAUN BLANQUET, J. 1979. Fitosociología: Bases para el estudio de las comunidades vegetales. Ed. Blume, Madrid.
CANTILLO, H. 2002 Diversidad y caracterización florística y estructural de la vegetación en la zona de captación de aguas de la microcuenca El Tigre -- municipio de Norcasia, Caldas. Tesis de grado - Maestría en Ecología. UNAL. Bogotá D.C.
CLEEF et al., 1984. En J.O. Rangel Ch, Lowy. P y Aguilar. M. 1997. Colombia Diversidad Biótica II. Instituto de Ciencias Naturales Universidad nacional de Colombia. Bogotá D.C.
CUATRECASAS, J. 1958 Aspectos de la vegetación natural de Colombia. Rev. Acad. Col. Cs. Ex. Fis. Nat. 10 (40): 221-268.
GALINDO & CARDENAS. 1999. Estudio de Vegetación en el Santuario de Fauna y Flora Guanentá-Alto río Fonce, departamentos de Santander y Boyacá. Tesis de Grado. Universidad Distrital Francisco José de Caldas. Bogotá D.C.
GENTRY, · A. 1993. Vistazo General a los Bosques Nublados Andinos y a la Flora de Carpanta. En: Andrade, G. Carpanta, Selva Nublada y Páramo. Colombia. Fundación Natura.
HILL, M. 1979. Twinspan -- A Fortran program for arranging multivariate data in an ordered two-way table by classification of the individuals and attributes. Cornell University, Ithaca. New York.
HOLDRIDGE, L. 1978. Ecología basada en zonas de vida. Traducción de H. Jiménez IICA. Serie de libros y materiales educativos No 34.
IGAC. 1996. Diccionario Geográfico de Colombia. Tomo II. Tercera Edición Colombia.
KUHRY, 1988. En J.O. Rangel Ch, Lowy. P y Aguilar. M. 1997. Colombia Diversidad Biótica II. Instituto de Ciencias Naturales UNAL. Bogotá D.C.
MATTEUCI, S. & COLMA, A. 1982. Metodología para el estudio de la vegetación. OEA. Estado de Falcón, Venezuela.
RANGEL, J.O. & GARZÓN, A. 1994. Aspectos de la estructura, de la diversidad y de la dinámica de la vegetación del parque regional Ucumari. Instituto de Ciencias Naturales Universidad nacional de Colombia. Bogotá, D.C.
RANGEL, J.O. & LOZANO, C. 1986. Un Perfil de Vegetación entre La Plata Huila y el Volcán Puracé. Caldasia 14 (68-70). Bogotá D.C.
RANGEL, J.O. & VELÁZQUEZ, A. 1997. Métodos de estudio de la vegetación. En: J.O. Rangel Ch, Lowy. P. y Aguilar. M. 1997. Colombia Diversidad Biótica II.
ROMERO, J. 1998. Comparación de los sistemas de producción (agrosistemas) en un gradiente altitudinal en el municipio de EncinoSantander. Universidad Javeriana. Bogotá, D.C.
STURGES, H.G, 1926. The Choice of class interval. Journal of the American Statistical Association.
TORRES, D. & TORRES L. 1997. Estructura, diversidad florística y crecimiento inicial de un bosque secundario y un bosque de roble en el santuario de fauna y flora de Iguaque. Universidad Distrital Francisco José de Caldas. Bogotá D.C.
VARGAS, W. 2002. Guía ilustrada de las plantas de las montañas del Quindío y Los Andes Centrales. Universidad de Caldas. Manizales. 803p.
VÁSQUEZ, G. 1987. Crecimiento de un bosque de Guandal explotado en el Litoral Pacífico colombiano. Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias. Medellín.
How to Cite
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Download Citation
Artículos de investigación científica y tecnológica
Colombia Forestal, 2005-11-00 vol:9 nro:18 pág:70-86
Caracterización florística y estructural de unidades de bosque altoandino en las veredas Minas y Patios Altos, Encino-Santander
Yira Gelit Ocaña Figueroa
Ingeniera Forestal, Universidad Distrital Francisco José de Caldas briz_o@hotmail.com
Resumen
En el bosque altoandino de las Veredas Minas y Patios Altos se logró clasificar florísticamente la alianza Clusio multiflorae-Quercoion humboldti, conformada por las asociaciones Miconio theaezandis-Quercetum humboldtii y Myrsino guianensis-Quercetum humboldtii. Además, se identificó la comunidad Hedyosmum bonplandianum-Quercus humboldtii.
El estrato arbóreo inferior resultó ser el de mayor cobertura. La especie de mayor IPF (Índice de Predominio Fisionómico) e IVI (Índice de Valor de Importancia) fue Quercus humboldtii, para individuos mayores de 10 cm de DAP . (Diámetro a la Altura del Pecho). Las especies que registraron los mayores valores en regeneración natural son Vismia guianensis y Hedyosmum bonplandianum; Quercus humboldtii y Cecropia telenitida, y resultaron relevantes en cuanto a posición sociológica.
Las distribuciones de cobertura y diamétrica reflejaron una tendencia de "J" invertida. La distribución de altura indicó un comportamiento normalizado. La diversidad se manifestó con un total de 128 especies y 78 géneros pertenecientes a 41 familias. En el estrato herbáceo se encontró la mayor riqueza en especies. La dominancia del gremio ecológico heliófito en los estratos inferior y medio, evidenció un estado sucesional secundario del bosque, condicionado por el cambio de las características ambientales de acuerdo con el ascenso en altitud y al grado de intervención humana.
Palabras clave: bosque altoandino, composición florística, estructura, diversidad, Encino, Santander.
Abstract
In the altoandino forest of the Sidewalks Minas and Patios Altos, The floristic classification defined the alliance Clusio multifloraeQuercoion humboldtii conformed by the associations Miconio theaezandis-Quercetum humboldtii, and Myrsino guianensisQuercetum humboldtii. Also, the community Hedyosmum bonplandianum Quercus humboldtii was identified.
The stratum arboreal inferior turned out to be that of more covering. The species of more PPI (Physiognomic Predominance Index) and IVI . (Importante Value Index) were Quercus humboldtii, for individuals bigger than 10 cm diameter to the height of the chest. The species that register the biggest values in natural regeneration are Vismia guianensis and Hedyosmum bonplandianum; Quercus humboldtii and Cecropia telenitida, are important as for sociological position.
The diametric and covering distributions reflected a tendency of invested "J". The distribution of height shows a normalized behavior. The diversity is manifested with a total of 128 species and 78 genera belonging to 41 families, being in the herbaceous stratum is the biggest wealth in species. The dominance of the heliolilos ecological society in the inferior and half strata, evidences to secondary successional state of the forest, conditioned by changing of the environmental characteristics according to the ascent in altitude, and the incidence of human intervention.
Key words: Altoandino forest, floristic composition, structure, diversity, Encino, Santander.
INTRODUCCIÓN
En este estudio se buscó integrar varios conceptos con el fin de determinar y medir variables representativas de los caracteres fisonómicos y estructurales de la vegetación, como abundancia, frecuencia, cobertura y demás, que pueden ser interpretados mediante métodos numéricos para el análisis estadístico y espacial. Este estudio permitió establecer el estado sucesional del bosque, así como las migraciones y áreas de distribución de las comunidades vegetales encontradas y su estado de conservación, a través del cálculo de índices de diversidad y regeneración natural, entre otros.
OBJETIVO GENERAL
Realizar la caracterización florística y estructural de las diferentes unidades de vegetación encontradas en los bosques altoandinos de las veredas Minas y Patios Altos del municipio de Encino-Santander.
OBJETIVOS ESPECÍFICOS
- Identificar unidades de vegetación en los bosques altoandinos de las veredas Minas y Patios Altos del municipio de EncinoSantander.
- Determinar la composición florística y estructural de la vegetación encontrada.
- Definir la fisionomía y estructura vertical y horizontal de la vegetación, mediante el análisis de variables representativas como abundancia, frecuencia, estrato y cobertura.
- Determinar el estado de conservación del bosque altoandino, mediante el análisis de índices de diversidad.
ÁREA DE ESTUDIO
El municipio del Encino se encuentra ubicado en la vertiente occidental de la Cordillera Oriental, al sur oriente del departamento de Santander. La cabecera municipal está situada a 06°08'27" de latitud norte y a 73°06'07' de longitud oeste a una altura de 1820 msnm. Comprende 13 veredas categorizadas en "bajas" (1400-2000 msnm) y "altas" (2200-3000 msnm). El área de estudio está comprendida entre las veredas Minas y Patios Altos, al suroriente de Encino, entre los 2400 y 3200 msnm; la totalidad de las tierras con cobertura boscosa son de propiedad privada.
En cuanto a las formaciones vegetales, según Romero (1998), estas veredas pertenecen a un mismo sistema agroecológico, denominado zona de montaña alta de papales, que presenta condiciones climáticas y socioeconómicas similares. Esta zona comprende ante todo bos que natural de roble (Quercus humboldtii) de carácter secundario, altamente fragmentado debido a la ampliación de la frontera agropecuaria. De acuerdo con la clasificacsión climática de Holdridge (1978), el área de estudio está comprendida dentro de dos zonas de vida: bosque muy húmedo montano bajo (bmh-MB) y bosque pluvial montan() (bp-M).
El área presenta una temperatura media anual de 18°C y una precipitación media anual de 3281 mm (IGAC 1996). El régimen de lluvias es bimodal, con dos períodos secos que comprenden los meses de enero y febrero, el primero, y los de junio y agosto, el . segundo; clo's períodos de lluvias, el primero entre los meses de abril y mayo, y el segundo, entre los de octubre y noviembre. El uso actual de la tierra corresponde principalmente a áreas de pastos dedicadas a la ganadería extensiva, zonas de pastos enrastrojados, rastrojos bajos y altos, pequeñas áreas destinadas a la agricultura de autoconsumo, relictos de bosque primario y bosque secundario. El relieve del área es en su mayoría pronunciado y fuerte; montañoso con pendientes mayores al 75% y colinado montañoso con pendientes entre 50 y 75% (IGAC, 1996).
MATERIALES Y MÉTODOS
Se recolectó la información de campo a partir del muestreo de unidades aleatorias rectangulares, de 500 m2 y 250 m2 (Rangel, 1995) de acuerdo con la altitud. Para los estratos superiores (fustales) se tomaron datos de DAP, altura total, altura a la primera rama, diámetro de copa, coordenadas (x,y) con respecto al límite preestablecido de la parcela. Esta medición se facilitó delimitando subparcelas de 10 x 10 m; se realizaron observaciones adicionales. Para los estratos inferiores se delimitaron subparcelas de 2 x 2 m y 1 x 1 m para latizos y brinzales, respectivamente. Se recolectaron datos de presencia, abundancia, altura y cobertura de cada categoría. Las espeéies objeto de medición se marcaron adecuadamente y se tomaron muestras de cada una para su posterior identificación en el Instituto de Ciencias Naturales de la Universidad Nacional y el Herbario Gilberto Mahecha Vega de la Universidad Distrital Francisco José de Caldas.
Según la metodología desarrollada por Cantillo (2002), se realizó una definición de comunidades mediante una tabla fitosociológica a partir de una matriz de coberturas entre especies contra levantamientos. Se utilizó el programa Twinspan (Hill, 1979), usando las coberturas por especie de los diferentes estratos. Se determinó para cada comunidad de vegetación la estructura horizontal y los índices de valor de importancia (IVI) de Cottam (Matteucci y Colma, 1982) y el índice de valor de importancia ampliado (IVIA). Los estratos de altura se definieron según las categorías propuestas por Rangel y Lozano (1986).
De acuerdo con la metodología desarrollada por Cantillo (2002), se definió el ordenamiento en sentido vertical y horizontal de los grupos fitosociológicos. En sentido vertical se logró por medio de la estratificación, y en sentido horizontal por medio de variables como densidad, área basal y cobertura (Rangel y Velásquez, 1997). Se determinó el índice de Diversidad Alfa, los índices de Menhinick, Margalef y coeficiente de mezcla de Holdridge. Se calcularon los índices de Shanon-Weaver, Simpsom y Berger-Parker.
RESULTADOS
CARACTERIZACIÓN FLORÍSTICA
Clasificación
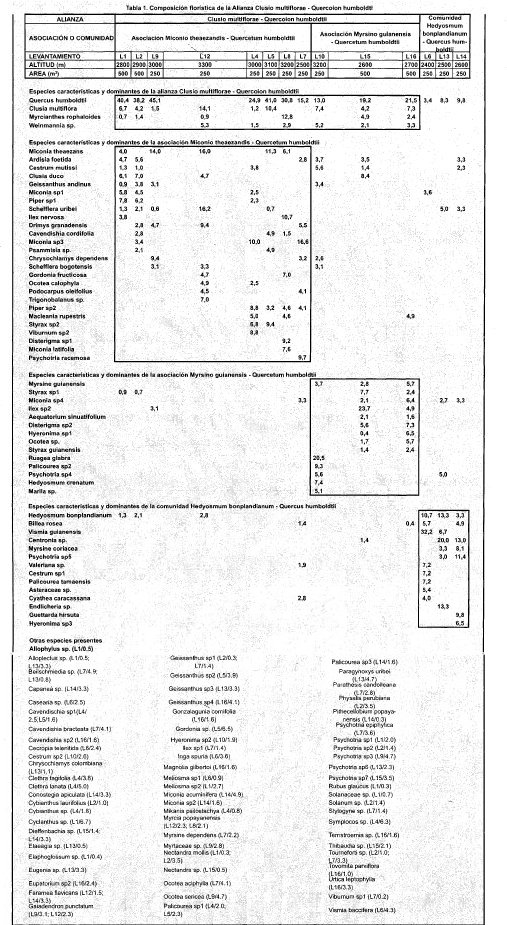
Al realizar el análisis fitosociológico propuesto por Braun -- Blanquet (1979), se clasificó la vegetación en una alianza, compuesta por dos asociaciones y una comunidad (Tabla 1). A continuación se muestran las características florísticas, fisonómicas y ecológicas de cada una de las anteriores:
Alianza Clusio multiflorae-Quercoion humboldti
Asociación tipo: Miconio theaezandis-Quercetum humboldtii
Otras Asociaciones: Myrsino guianensisQuercetum humboldtii
Composición Florística: Como especies características se encuentran: Miconia theaezans, Myrcianthes rophaloides y Weinmannia sp.
Fisionomía Vegetación de porte medio, con alturas superiores que no exceden los 25 m, abundan las especies epífitas, lianas y quiches; dosel cerrado. No se presenta estrato,arbóreo superior, el estrato arbóreo inferior tiene una cobertura de 70%. Los estratos de arbolitos y arbustivo cubren un 19 y 24% respectivamente, mientras el estrato herbáceo posee una cobertura promedio de 16%.
Ecología Localizada sobre pendientes fuertes que en promedio superan el 52%. Suelos moderadamente profundos, de textura francoarcillosa y pH's ácidos. Dominan las especies heliófitas en el estrato medio, una especie hemiesciófita en el estrato superior, y comparten dominancia las esciófitas y hemiesciófitas en el estrato inferior.
Asociación Miconio theaezandis - Quercetum humboldtii
Composición Florística Se encuentran como especies características: Ardisiafoetida, Cestrum mutissi, Clusia duco, Geissanthus andinus, Miconia sp., Schefflera uribei e Ilex nervosa.
Fisionomía Vegetación arbórea medianamente densa, individuos que no superan los 23 m de altura. Abundante presencia de epífitas. El estrato arbóreo inferior cubre cerca del 75%, los estratos arbustivo y de arbolitos un 19 y 16%, respectivamente, y el estrato herbáceo tiene una cobertura cercana al 9%.
Ecología Se localiza sobre suelos medianamente profundos, de pH ácidos y textura fina en su mayoría francoarcillolimosa, la pendiente promedio es del 60%. Una especie hemiesciófita domina el estrato superior, en el estrato medio la dominancia es de las especies de carácter heliófito, y en el estrato inferior es relevante la presencia de las especies esciófitas y hemiesciófitas.
Asociación Myrsino guianensis-Quercetum humboldtii
Composición florística Se encuentran como especies características las siguientes: Ruagea glabra, Palicourea sp., Psychotria sp., Hedyosmum crenatum, Marila sp.
Fisionomía Árboles que no exceden los 16 m de altura, vegetación medianamente densa, abundantes epífitas. El estrato arbóreo inferior tiene una cobertura promedio del 64%, el estrato de arbolitos cubre un 13%, y los estratos arbustivo y herbáceo tienen una cobertura del 9 y 6 %, respectivamente.
Ecología Se localiza sobre pendientes que superan el 35%, suelos moderadamente profundos, de textura francoarcillosa y pH ácidos. Una especie de carácter hemiesciófito posee mayor presencia en el estrato superior, mientras las especies heliófitas y hemiesciófitas dominan en los estratos medio e inferior.
Comunidad Hedyosmum bonplandianumQuercus humboldtii
Composición florística Las especies características son: Cestrum sp., Palicourea tamaensis, Asteraceae sp., Cyathea caracassana.
Fisionomía Vegetación arbórea medianamente densa, individuos que alcanzan alturas inferiores a los 16 m. En esta comunidad el estrato arbóreo superior cubre un 4%, mientras el estrato arbóreo inferior tiene una cobertura del 50%, los estratos de arbolitos, arbustivo y herbáceo cubren un 16, 15 y 5%, respectivamente.
Ecología Se ubica sobre pendientes fuertes superiores al 60%, que se suavizan al llegar a las zonas más altas (3350 msnm), suelos poco profundos, de textura francoarcillosa y pH ácidos. Las especies heliófitas y hemiesciófitas dominan en los estratos medio y bajo, y una especie hemiesciófita resalta su presencia en el estrato superior.
ANÁLISIS DE LA ESTRUCTURA
Altura
La distribución de alturas para los individuos con DAP > 10 cm en la alianza Clusio multiflorae-Quercoion humboldti (Figura 1), identifica nueve clases según el método de distribución de Sturges (1926). Para este caso, el mayor número de individuos se concentra en la categoría III (8-10 m) con un porcentaje de 21.1% de un total de 232 individuos (Figura 1), le sigue la clase IV (10-12 m) con un total de 42 individuos correspondientes a un 18.1%. La asociación Miconio theaezandis-Quercetum humboldtii (Figura 2), identifica también, nueve clases de alturas, donde el mayor número de individuos (24) se agrupan en la clase VI (1416 m) representando un porcentaje de 16.4% del total, un 15.7% corresponde a la clase IV (10-12m). El estrato inferior (clase uno, 4-6 m) es el que menor número de individuos posee (6), le sigue el estrato superior o dominante (clase IX >20 m) con nueve individuos.
En la asociación Myrsino guianensis-Quercetum humboldtii (Figura 3), se distinguen únicamente seis clases de alturas, los mayores porcentajes de individuos corresponden a las clases III (8-10 m) y IV (10-12 m), con un 34.9 y 22.1%, respectivamente. El menor porcentaje (4.6%) corresponde a la clase dominante (VI 14-16 m), representada tan solo por 4 individuos. La comunidad Hedyosmum bonplandianum-Quercus humboldtii (Figura 4), identifica también seis clases de alturas, de las cuales las tres primeras poseen el mayor número de individuos, con un porcentaje superior de 39.5% para la clase III (12-16 m), seguida de la clase II (8-12 m) con un 18.9%. La clase V (20-24 m) no posee individuos, y la clase VI (24-28 m) de estrato superior, tan solo posee un individuo.
Cobertura
El análisis de cobertura se realizó a partir de la distribución de coberturas propuesta por Rangel y Velásquez (1997). En la alianza Clusio multiflorae- Quercoion humboldti, los valores de cobertura se distribuyen en nueve clases, 164 individuos pertenecen a la clase uno, mientras las últimas cinco categorías están representadas solo por un individuo, que corresponde únicamente al 2.2% del total; las primeras cuatro categorías constituyen el 97.8%. El rango de amplitud es 17 m2, y la diktribución muestra un comportamiento muy regular (Figura 5). La asociación Miconio theaezandis Quercetum humboldtii presenta ocho clases con una amplitud de 19 m2 (Figura 6), al igual que en el anterior caso, la mayor frecuencia de individuos se presenta en la clase número uno que cuenta con 100 individuos, las primeras cuatro clases representan el 97.2%, mientras que las últimas corresponden al 2.8%, de un total de 143 individuos.
En la asociación Myrsino guianensis--Quercetum humboldtii, la cobertura se distribuye en siete clases (Figura 7), el rango de amplitud es de 5 m2 . Las primeras dos clases corresponden a un 32.5 y 27.3%, respectivamente, las clases tres y cuatro representan cada una 14.4 %, y los restantes porcentajes de 8.4% y 2.4% son los dé las clases cinco y siete, respectivamente. La clase número seis no tiene representantes.
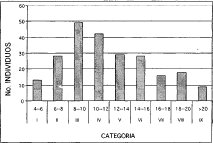
Figura 1. Distribución de Altura Alianza
Clusio multiflorae - Quercoion humboldti
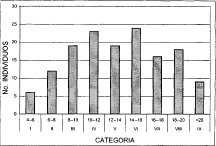
Figura 2. Distribución de Altura Asociación
Miconio theaezandis - Quercetum humboldtii
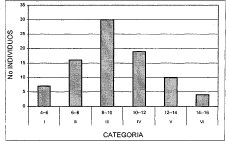
Figura 3. Distribución de Altura Asociación
Myrsino guianensis --Quercetum humboldtii
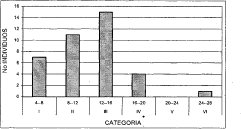
Figura 4. Distribución de Altura Comunidad
Hedyosmum bonplandianum- Quercus humbol
La comunidad Hedyosmum bonplandianum Quercus humboldtii presenta seis clases con una amplitud de 4 m2 (Figura 8). Las primeras dos clases corresponden a un 32.4 y 24.3%, respectivamente. Las clases tres y cuatro representan cada una un 13.5% y 8.2%, al igual que las clases cinco y seis, respectivamente.
Diámetro
En la medición de la estructura diarnétrica se muestrearon todos los individuos con DAP > 10 cm. En general se obtuvo que el 44.6% de los individuos hacen parte del rango de 10-20 cm DAP, la distribución diamétrica general para la unidad de bosque de roble (Figura 9). La distribución diamétrica general describe un comportamiento típico de bosque disetáneo, teniendo en cuenta una distribución normal en forma de "J" invertida.

Figura 5. Distribución de Coberturas Alianza
Clusio multiflorae - Quercoion humboldti

Figura 6. Distribución de Coberturas Asociación
Miconio theaezandis - Quercetum humboldtii

Figura 7. Distribución de Coberturas Asociación
Myrsino guianensis Quercetum humboldtii

Figura 8. Distribución de Cobertura Comunidad
Hedyosmum bonplandianum - Quercus humboldtii
Al correr un modelo de correlación bajo una distribución exponencial, en la figura anterior se observa un alto nivel de correspondencia (R2 = 0.9439) entre el número de individuos y las clases diamétricas, después de trabajar de forma agregada toda la unidad de bosque de roble. La alianza Clusio multiflorae Quercoion humboldti en la (Figura 10), presenta nueve clases diamétricas, de las cuales, la de mayor número de individuos (103) es la clase uno (10-17 cm DAP), las clases VII y IX (52-59 cm y 66-73 cm DAP, respectivamente) solo cuentan con tres representantes. En la asociación Miconio theaezandis-Quercetum humboldtii se tienen ocho clases diamétricas (Figura 11), domina la clase uno (10-18 cm) con el mayor número de individuos (72), y la clase VIII (66-74cm) posee tan solo tres representantes. La asociación Myrsino guianensisQuercetum humboldtii está distribuida en seis categorías diamétricas, como se ilustra en la (Figura 12), la clase uno (10-15 cm) posee 25 individuos, mientras la clase VI con tan solo dos individuos, posee diámetros mayores a los 40 cm. La distribución diamétrica de la comunidad Hedyosmum bonplandianumQuercus humboldtii (Figura 13), se manifiesta con seis categorías, siendo considerable el número superior de individuos (26) de la clase uno (10-21cm), mientras la clase VI (>65 cm) solo posee un representante.
ÍNDICES ESTRUCTURALES
Índice de Predominio Fisionómico (IPF)
En la alianza Clusio multiflorae-Quercoion humboldti puede observarse la total dominancia de la especie Quercus humboldtii, tan solo dos especies superan el 3% como valor de IPF (Clusia multiflora, Weinnmania sp.). Las cinco especies con mayor valor de IPF (Figura 14), representan el 86.7% del valor total de este índice, mientras que el 13.3% restante está constituido por especies que en su mayoría no superan el 1% en el valor del índice de predominio fisionómico. En la asociación Miconio theaezandis-Quercetum humboldtii la situación es muy similar; la especie Quercus humboldtii posee un 'valor de IPF superior al 70%; le siguen en orden las especies Schefflera uribeii (4%), Clusia multiflora (3.3%), Weinnamnia sp. (2.5%) y Beils chmiedia sp. (2.4%) que, junto con Quercus humboldtii, representan el 86.5% del valor total de este índice (Figura 15).
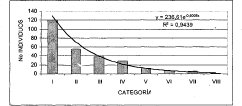
Figura 9. Distribución Diamétrica general bosque de Quercus humboldtii Individuos con DAP > 10 cm
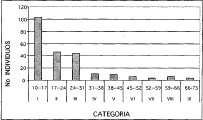
Figura 10. Distribución Diamétrica Alianza
Clusio multiflorae Quercoion humboldti
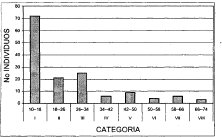
Figura 11. Distribución Diamétrica Asociación
Miconio theaezandis - Quercetum humboldtii
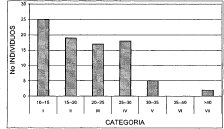
Figura 12. Distribución Diamétrica Asociación Myrsino guianensis -- Quercetum humboldtii
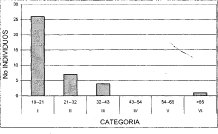
Figura 13. Distribución Diamétrica Comunidad
Hedyosmum bonplandianum - Quercus humboldtii
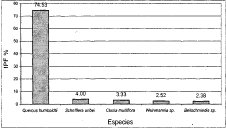
Figura 14. IPF Alianza
Clusio multiflorae - Quercoion humboldtii
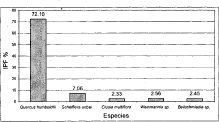
Figura 15. IPF Asociación
Miconio theaezandis - Quercetum humboldtii
En la asociación Myrsino guianensis-Quercetum humboldtii las cuatro especies con mayor porcentaje de IPF después de la dominante Quercus humboldtii (70.9%) son (Figura 16): Clusia multiflora (10%), Weinmmania sp. (6.7%), Schefflera bogotensis (3.4%) y Geissanthus andinus (3%). En la comunidad Hedyosmum bonplandianum-Quercus humboldtii la especie Quercus humboldtii reduce su dominancia a un valor de IPF de 39.5%, le siguen en orden (Figura 17): Billea rosea (9.6%), Cecropia telenitida (9.5%), Parathesis candolleana (9.3%) y Vismia baccifera (5.8%).
Índice de Valor de Importancia (IVI)
El cálculo del. IVI se realizó para todos las especies con individuos con DAP ? 10 cm en cada una de las comunidades. En la alianza Clusio multiflorae-Quercoion humboldtii, la especie que mayor valor de IVI registra es Quercus humboldtii con un porcentaje de 60.3%, el 39.7% restante corresponde a 30 especies cuyo valor de IVI es poco considerable. Las cuatro especies con mayor valor de IVI (Figura 18) después del roble son: Clusia multiflora (5.6%) Weinmannia sp. (5.1%), Schefera uribei (3.8%) y Beilschmiedia sp. (2,3%).
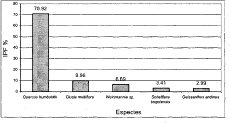
Figura 16 IPF Asociación
Myrsino guianensis -- Quercetum humboldtii

Figura 17. IPF Comunidad
Hedyosmum bonplandianum - Quercus humboldtii
De la misma forma en la asociación Miconio theaezandis-Quercetum humboldtii, la especie dominante es Quercus humboltii con un valor de IVI cercano al 60%, le siguen en orden las especies (Figura 19) Schefflera uribei (6.1%), Clusia multiflora (4%), Weinmannia sp. (3,4%) y Beilschmiedia sp. (3,4%). En la asociación Myrsino guianensis-Quercetum humboldtii el roble es la especie dominante con un valor de IVI relativo de 59.8%. Las cuatro especies con mayor porcentaje de IVI (Figura 20) después del roble son: Weinmannia sp. (10.3%), Clusia multiflora (9.6%), Schefflera bogotensis (5%) y Geissanthus andinus (4,5%). En la comunidad Hedyosmum bonplandianum-Quercus humboldtii, la especie Quercus humboldtii tiene el mayor valor de IVI (34.4%), le siguen en orden (Figura 21) Cecropia telenitida (9.3%), Billea rosea (9%), Parathesis candolleana (8.2%) y Vismia baccifera (5.4%). El IVI relativo de otras especies presentes indica que la dominancia se distribuye de manera más uniforme que para los casos anteriores.
Índice de Valor de Importancia Ampliado (IVIA)
Al tener en cuenta la posición sociológica y el porcentaje de regeneración de las especies con DAP≥10 cm, se reafirma la dominancia de la especie Quercus humboldtii, en la alianza Clusio multiflorae--Quercoion humboldti esta especie obtuvo un valor de 50.9%, aunque éste es ligeramente menor que el de IVI, es significativamente mayor en relación con las otras especies, de las cuales las cuatro que mayor valor de IVIA obtuvieron son: Clusia multiflora (6,2%), Weinmannia sp. (4,7%), Schefflera uribei (4%) y Drimys granadensis (2,8%).

Figura 18
IVI Alianza
Clusio multiflorae-Quercoion humboldtii
Individuos con DAP > 10 cm
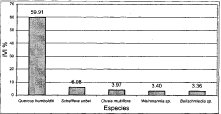
Figura 19 IVI Asociación
Miconio theaezandis-Quercetum humboldtii
Individuos con DAP > 10 cm
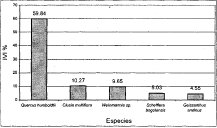
Figura 20 IVI Asociación
Myrsino guianensis- Quercetum humboldtii
Individuos con DAP > 10 cm
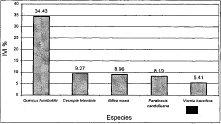
Figura 21 IVI Comunidad
Hedyosmum bonplandianum-Quercus humboldtii
Individuos con DAP > 10 cm
En la asociación Miconio theaezandis-Quercetum humboldtii, la especie Quercus humboldtii obtuvo un valor de IVIA mayor al 50%, le siguen en orden las especies Schefflera uribei (6,3%), Clusia multiflora (4,6%), Drimys gnanadensis (4,2%) y Miconia sp 1 (3,8%). En la asociación Myrsino guianensis-Quercetum humboldtii, el valor de IVIA que registra la especie Quercus humboldtii es de 55.9%, siendo las cuatro especies que le prosiguen Clusia multiflora (10,3%), Weinmannia sp. (10%), Mynsine coniacea (6,8%) y Schefflera bogotensis (4%). En la comunidad Hedyosmum bonplandianum-Quercus humboldtii, el mayor valor de IVIA (26.7%) corresponde igualmente a la especie Quercus humboldtii; no obstante, y al igual que para el valor de IVI, la dominancia de esta especie disminuye considerablemente, permitiendo que otras especies se destaquen, como lo son: Vismia guianensis (11,2%), Billea vosea (10,6%), Cecnopia telenitida (8,5%), Panathesis candolleana (6,5%).
DIVERSIDAD
El total de especies encontradas es de 128, pertenecientes a 78 géneros, distribuidos en 41 familias (Tabla 2). La familia con mayor número de especies representantes es Rubiaceae (18) que corresponde a un 14.1%, le siguen en orden las familias Myrsinaceae (13 especies, 10.2%), Ericaceae y Melastomataceae (9 especies, 7%), Clusiaceae y Lauraceae (8 especies, 6.2%). El restante 49.2% corresponde a familias que poseen entre una y cinco especies.
Tabla 2. Número de especies, géneros y familias por comunidad.

Diversidad por estratos
En el estrato arbóreo se encontraron 18 especies correspondientes al 14% del total. Las familias Clusiaceae y Myrsinaceae con 3 y 2 especies, representan el 16.7 y 11.1% del estrato, respectivamente. El restante 72.2% corresponde a 13 familias que solo poseen una especie. En el estrato de arbolitos se identificaron 30 especies equivalentes al 23.4% del total. De 19 familias encontradas la familia Clusiaceae es la más representativa con siete especies equivalentes a un 23.3% en el estrato. Le siguen en orden las familias: Myrsinaceae (4 especies, 13.3%), Araliaceae y Rubiaceae (2 especies, 6.7%). En el estrato arbustivo se encontraron 81 especies que corresponden al 63.3% del total. La familia Myrsinaceae domina en el estrato con 10 especies representantes equivalentes al 12.4%. Las familias Clusiaceae y Rubiaceae tienen cada ocho especies que corresponden al 9.9% del total en el estrato, y le siguen las familias Ericaceae y Melastomataceae cada una con seis especies equivalentes al 7.4%. En el estrato herbáceo se identificaron 75 especies que corresponden al 58.6% del total de especies muestreadas. La familia Rubiaceae es la más representativa con 10 especies que equivalen al 13.3% del total en el estrato. Le siguen las familias: Ericaceae (8 especies, 10.7%), Melastomataceae (7 especies, 9.3%), Myrsinaceae (6 especies, 8%), Asteraceae y Clusiaceae (5 especies, 6.7%).
Índices de riqueza
El análisis de riqueza se realizó a partir del cálculo de los índices de Margalef, Menhinick y el cociente de mezcla de Holdridge (Tabla 3). El mayor valor en el índice de Margalef fue para la alianza Clusio multiflorae-Quercoion humboldti con 5.49. Se encontró un muy bajo grado de riqueza ya que el cociente de mezcla de Holdridge no supera la unidad en ninguno de los casos. La relación más cercana al 1:1 entre el número de especies y el de individuos, se presenta en la comunidad Hedyosmum bonplandianum-Quercus humboldtii donde se obtuvo un cociente de 0.42.
Tabla 3. Índices de riqueza

Índices de diversidad
El análisis de diversidad se realizó a partir del cálculo de los índices de Shannon-Wiener y Simpson. Se determinó, además, el índice de uniformidad de Pielou para cada una de las comunidades. Los resultados pueden apreciarse en la Tabla 4.
Tabla 4. Índices de diversidad

DINÁMICA
En el estrato Arbóreo, además de la especie dominante Quercus humboldtii, que concentra su mayor número de individuos en este estrato, se destacan las especies Beilschmiedia sp., Ilex nervosa, Cecropia telenitida y Clusia multiflora, de hábito arbóreo y que poseen de cuatro a cinco individuos en el dosel superior. En el estrato de Arbolitos, la especie Quercus humboldtii es la que posee mayor número de representantes; sin embargo, el número de individuos es significativamente menor que para el caso anterior. En el rango de altura de 5 a 12 m resalta la presencia de otras especies como Weinmannia sp., Clusia multiflora y Schefflera uribei. En el estrato arbustivo el núm'éro de especies se incrementa, y adicionalmente a la especie dominante Quercus humboldtii, se destacan las especies Miconia spp., Myrsine guianensis, Miconia theaezans, Ilex sp., Schefflera uribei, Clusia duco y Clusia multiflora.
En el estrato inferior, la especie con mayor número de individuos sigue siendo Quercus humboldtii, pero en este rango de altura su número de representantes no es altamente superior en comparación con los casos anteriores. Se destacan además muchas más especies como Piper spp., Psychotria spp., Miconia spp., Disterigma sp., Geissanthus andinus, Styrax spp., Drimys granadensis, Ardisia foetida y Ocotea sp. Las especies comunes a todos los estratos, además de la dominante Quercus humboldtii, que posee representantes en todos los estratos, son Clusia multiflora, Miconia spp., Schefflera uribei, Vismia guianensis y Weinmannia sp.
DISCUSIÓN
La composición florística general de las unidades de bosque altoandino de las Veredas Minas y Patios Altos del municipio de Encino muestra una clara disminución en el número de especies en la medida que se asciende altitudinalmente, hecho que está en concordancia con lo encontrado por Galindo y Cárdenas, (1999) quienes mencionan en su estudio, que la riqueza de familias y especies está de acuerdo con la disminución en altitud.
Los grupos fitosociológicos encontrados no están reportados en la literatura; sin embargo, Cleef et al., (1984) y Rangel y Garzón (1994), identifican a la asociación Clusio multiflorae - Weinmannietum pinnatae, en la Sierra Nevada de Santa Marta, en alturas entre los 2475 y 3000 msnm. Por su parte Kuhry (1988), encontró a la especie Quercus humboldtii asociada a especies de los géneros Inga, Cecropia, Hedyosmum y Billea (1000-2300 msnm). En el transecto Valle del Magdalena - Volcán Puracé (Rangel y Lozano, 1986; 1989), se identifica a la asociación Hedyosmo - Quercetum humboldtii, con dominancia absoluta del roble y en alturas entre los 2200 y 2700 msnm. En la parte alta del transecto Parque de los Nevados, Lozano y Torres (1974) reportan a la especie Quercus humboldtii (2500-3100 msnm) asociada a algunas especies como Weinmannia sp., Clusia sp., dentro del estrato arbóreo; a las especies Drymis granadensis, Schefflera sp., en el estrato de arbolitos y Psychotria sp., en el estrato herbáceo.
El análisis de la estructura vertical demuestra que el bosque se distribuye en tres estratos principales. De acuerdo con Andrade (1993), en los bosques tropicales de montaña se observa, aunque no tan pronunciado, el efecto de estratificación vertical. En el estudio realizado por Torres y Torres (1997), el bosque también se estratifica en tres grupos de individuos, aunque para este caso el rango de alturas es menor y menos amplio. También se menciona que la mayoría de individuos se concentra en los estratos intermedios, tal como resultó en el presente estudio; así como también es similar la situación en los estratos inferiores donde se registra un número muy reducido de individuos, lo cual vislumbra un futuro muy preocupante para el bosque.
La distribución de cobertura y diámetro de los grupos fitosociológicos encontrados, muestra, de manera aislada, baja correspondencia con la distribución exponencial (típica de bosques heterogéneos disetáneos, Vásquez (1987); lo anterior genera un indicio sobre un alto nivel de perturbación y conllevó a un nuevo análisis en el que se tomaron de forma simultánea todos los levantamientos, en éste último caso, se encontró una alta correspondencia entre las variables mencionadas, lo cual señala que en las diferentes áreas muestreadas debe desarrollarse una estrategia de manejo de manera conjun ta y no desagregada.
En cuanto a los índices estructurales dado el carácter gregario y dominante del roble Quercus humboldtii, es natural que en todas las comunidades vegetales identificadas haya obtenido un valor de IVI, IPF e IVIA, notoriamente superior al de otras especies. El roble tiende a formar bosques monoespecíficos, donde su espesa copa no permite el desarrollo total de las especies establecidas bajo su cobertura (Vargas, 2002). Según Gentry (1993), es muy frecuente que una especie sea dominante de manera local en un bosque nublado.
En la medición de la regeneración natural y posición sociológica, se observó como Quercus humboldtii incrementaba el número de renuevos, brinzales y latizales, a medida que se pasaba de una franja altitudinal a otra superior, lo anterior se explica porque, con el aumento en altura, la especie disminuye en dominancia en los estratos superiores, ya que al parecer las condiciones ambientales que permiten su pleno desarrollo, se han modificado; de esta manera, los individuos que hacen parte de los estratos inferiores pueden instaurarse, al verse disminuidos los limitantes de competencia como la luz y los nutrientes. Por otro lado, los porcentajes de regeneración natural son alarmantes, ya que no superan el 20% en la mayoría de las comunidades, estos valores no solo corresponden a la especie dominante, sino a aquellas propias del interior del bosque como Clusia multiflora, Myrsine guianensis y Drymis granadensis. Lo anterior está en concordancia con los resultados obtenidos por Torres y Torres (1997), quienes citan a Devia et al., (1997) compartiendo una apreciación de preocupación frente a la presencia del roble en etapas sucesionales futuras.
En lo referente al análisis de diversidad y uniformidad, los índices de Shannon-Wiener, Simpson, Berger Parker y Pielou señalan que la mayoría de grupos fitosociológicos tienen una diversidad medianamente baja, donde se ratifica el carácter monoespecífico y dominante de la especie Quercus humboldtii, y la homogeneidad del bosque en general. Sin embargo, la comunidad Hedyosmum bonplandianum-Quercus humboldtii es la excepción ya que registró altos valores en estos índices, comprobando lo antes mencionado acerca de la relación inversamente proporcional entre la altura y el número de especies, ya que esta comunidad se desarrolla en el menor piso altitudinal muestreado (2400-2600 msnm). No obstante, la mayor riqueza y diversidad de la comunidad Hedyosmum bonplandianurn-Quercus humboldtii también puede señalar un alto grado de intervención, teniendo en cuenta la presencia de especies como Cecropia telenitida que tiene entre sus características más importantes colonizar claros y crecer en terrenos altamente degradados (Vargas, 2002).
Los grupos fitosociólogicos identificados en el presente estudio se asemejan a las comunidades mencionadas al inicio de esta discusión (Rangel et al., 1997), en el número de especies encontradas en el estrato arbóreo, sin embargo, difieren considerablemente en cuanto al número de especies pertenecientes a los estratos arbustivo y herbáceo, siendo este significativamente mayor para el caso de las comunidades de las veredas Minas y Patios Altos (Encino-Santander). Lo anterior indica un estado sucesional secundario, donde el desarrollo de las especies pioneras como Cecropia telenitida está permitiendo la gradual modificación de las características ambientales hacia el establecimiento de especies de los gremios ecológicos esciófito y hemisciófito, como Ocotea sp. y Quercus humboldtii, respectivamente.
CONCLUSIONES
El análisis fitosociológico de los datos recolectados en campo permitió la identificación de cuatro unidades diferentes de vegetación en los bosques alto andinos de las Veredas Minas y Patios Altos, Encino-Santander. Las unidades de vegetación encontradas corresponden a la alianza Clusio multiflorae--Quercóion humboldti, a las asociaciones Miconio theá-ezandisQuercetum humboldtii y Myrsino guianensisQuercetum humboldtii y a la comunidad Hedyosmum bonplandianum-Quercus humboldtii. Los anteriores ,s'on grupos de vegetación que se diferencia entre sí, por el comportamiento de variables como la distribución de altura y cobertura, que constituyen manifestaciones fieles de la dinámica de la vegetación al interior de cada una de las unidades identificadas. En forma general, las diferentes unidades cuentan con la presencia significativa de especies características de este tipo de bolque como lo son, además de la especie dominante Quercus humboldtii, las especies Ciusia multiflora, Weinmannia sp., Myrsine guianensis, Drymis granadensis, Miconia spp. y Billea rosea, principalmente; sin embargo, se destaca tarnbi,ép, la presencia de especies típicas de bosques secundarios como Cecropia talenitida, hecho que da una pauta acerca del estado de conservación del bosque, y las posibles consecuencias de las alteraciones antrópicas.
Se estableció a partir del análisis de la distrubución de altura para cada una de las comunidades vegetales, que los individuos se agrupan en los diferentes estratos y muestran un comportamiento regular, en el cual las clases de alturas intermediasconstituye el mayor aporte. En cuanto a la estructura horizontal, la distribución diamétrica manifiesta un comportamiento típico de bosque disetáneo, con el mayor número de individuos agrupados en las categorías diamétricas inferiores, y pese a que existen pocos individuos en las clases diamétricas superiores, éstas representan el estrato dominante y el que constituye el mayor aporte en términos de área basal.
Los índices de predominio fisionómico (IPF), valor de importancia (IVI) y valor de importancia ampliado (IVIA), registraron los mayores valores para la especie Quercus humboldtii, que por sus características naturales predomina sobre las demás especies, limitando su desarrollo. Esta especie está presente en todos los estratos encontrados y domina totalmente tanto en abundancia como en frecuencia y cobertura; además obtuvo también los mayores porcentajes de regeneración y posición sociológica.
En el transcurso de la presente investigación, tanto en la toma de datos en campo como en su posterior análisis, se pudo comprobar cómo la vegetación varía su estructura a medida que se asciende altitudinalmente, ya que en el análisis preliminar para el cálculo del tamaño de la muestra se encontró que el número de especies disminuía a medida que aumentaba la altitud; sumado además el hecho de que los grupos fitosociológicos encontrados agrupan en su mayoría a los levantamientos establecidos en limites altitudinales comunes.
Los índices de riqueza y diversidad muestran en su mayoría un bosque de carácter homogéneo, con diversidad media a baja, características que concuerdan con el carácter monoespecífico y gregario de la especie dominante. Sin. embargo, esta situación no es del todo alentadora si se tienen en cuenta factores adicionales como el bajo porcentaje de regeneración natural y la poca presencia de individuos en los estratos in feriores. Lo anterior pone de manifiesto un preocupante futuro para la conservación y desarrollo normal del bosque de roble; no obstante, esta apreciación puede ser un tanto apresurada, teniendo en cuenta que el área muestreada es muy reducida y que no se tienen registros adicionales acerca de la estructura y composición florística del área.
REFERENCIAS BIBLIOGRÁFICAS
- ANDRADE, G. 1993. Paisaje y biodiversidad en las selvas de los Andes. Proyecto Reserva Biológica Carpanta. Fundación Natura, EAAB & INDERENA. Bogotá, D.C.
- BRAUN BLANQUET, J. 1979. Fitosociología: Bases para el estudio de las comunidades vegetales. Ed. Blume, Madrid.
- CANTILLO, H. 2002 Diversidad y caracterización florística y estructural de la vegetación en la zona de captación de aguas de la microcuenca El Tigre -- municipio de Norcasia, Caldas. Tesis de grado - Maestría en Ecología. UNAL. Bogotá D.C.
- CLEEF et al., 1984. En J.O. Rangel Ch, Lowy. P y Aguilar. M. 1997. Colombia Diversidad Biótica II. Instituto de Ciencias Naturales Universidad nacional de Colombia. Bogotá D.C.
- CUATRECASAS, J. 1958 Aspectos de la vegetación natural de Colombia. Rev. Acad. Col. Cs. Ex. Fis. Nat. 10 (40): 221-268.
- GALINDO & CARDENAS. 1999. Estudio de Vegetación en el Santuario de Fauna y Flora Guanentá-Alto río Fonce, departamentos de Santander y Boyacá. Tesis de Grado. Universidad Distrital Francisco José de Caldas. Bogotá D.C.
- GENTRY, · A. 1993. Vistazo General a los Bosques Nublados Andinos y a la Flora de Carpanta. En: Andrade, G. Carpanta, Selva Nublada y Páramo. Colombia. Fundación Natura.
- HILL, M. 1979. Twinspan -- A Fortran program for arranging multivariate data in an ordered two-way table by classification of the individuals and attributes. Cornell University, Ithaca. New York.
- HOLDRIDGE, L. 1978. Ecología basada en zonas de vida. Traducción de H. Jiménez IICA. Serie de libros y materiales educativos No 34.
- IGAC. 1996. Diccionario Geográfico de Colombia. Tomo II. Tercera Edición Colombia.
- KUHRY, 1988. En J.O. Rangel Ch, Lowy. P y Aguilar. M. 1997. Colombia Diversidad Biótica II. Instituto de Ciencias Naturales UNAL. Bogotá D.C.
- MATTEUCI, S. & COLMA, A. 1982. Metodología para el estudio de la vegetación. OEA. Estado de Falcón, Venezuela.
- RANGEL, J.O. & GARZÓN, A. 1994. Aspectos de la estructura, de la diversidad y de la dinámica de la vegetación del parque regional Ucumari. Instituto de Ciencias Naturales Universidad nacional de Colombia. Bogotá, D.C.
- RANGEL, J.O. & LOZANO, C. 1986. Un Perfil de Vegetación entre La Plata Huila y el Volcán Puracé. Caldasia 14 (68-70). Bogotá D.C.
- RANGEL, J.O. & VELÁZQUEZ, A. 1997. Métodos de estudio de la vegetación. En: J.O. Rangel Ch, Lowy. P. y Aguilar. M. 1997. Colombia Diversidad Biótica II.
- ROMERO, J. 1998. Comparación de los sistemas de producción (agrosistemas) en un gradiente altitudinal en el municipio de EncinoSantander. Universidad Javeriana. Bogotá, D.C.
- STURGES, H.G, 1926. The Choice of class interval. Journal of the American Statistical Association.
- TORRES, D. & TORRES L. 1997. Estructura, diversidad florística y crecimiento inicial de un bosque secundario y un bosque de roble en el santuario de fauna y flora de Iguaque. Universidad Distrital Francisco José de Caldas. Bogotá D.C.
- VARGAS, W. 2002. Guía ilustrada de las plantas de las montañas del Quindío y Los Andes Centrales. Universidad de Caldas. Manizales. 803p.
- VÁSQUEZ, G. 1987. Crecimiento de un bosque de Guandal explotado en el Litoral Pacífico colombiano. Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias. Medellín.
Creation date:




License
Colombia Forestal retains the patrimonial rights (copyright) of the published works, and favors and allows the reuse of the same under the Creative Commons Attribution-ShareAlike 4.0 International license, so they can be copied, used, disseminated, transmitted and exhibited publicly, provided that:
You acknowledge the credits of the work in the manner specified by the author or licensor (but not in a way that suggests that you have their support or that they endorse your use of their work).