DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2016.2.a07Published:
2016-07-01Issue:
Vol. 19 No. 2 (2016): July-DecemberSection:
Scientific articleEstudio de los anillos de crecimiento de Cordia alliodora (Ruiz & Pav.) en Colombia
Tree ring study of Cordia alliodora (Ruiz & Pav.) in Colombia
Keywords:
dendrocronología tropical, Cordia alliodora, cambio climático, anillos de crecimiento, fenómenos de El Niño y La Niña (es).Downloads
References
Briceño-J, A. M., & Arévalo, F. R. (2010). Potencial dendroclimático de la madera de cinco especies del bosque seco tropical de Armero–Tolima. (Trabajo de maestría, Maestría en ciencias biológicas) Ibagué: Universidad del Tolima. 56p.
CATIE (Centro agronómico tropical de investigación y enseñanza). (1994). Laurel Cordia alliodora (Ruiz &Pav.) Oken Boraginaceae un árbol de uso múltiple. Colección materiales de extensión. Turrialba, Costa Rica. 476p.
Cook, E. R. (1985). A time series analysis approach to tree-ring standardization. (Ph. D. thesis) Tucson. University of Arizona. 89p.
Del Vallle, J. I., Ramírez, J. A., & Herrera, D. A. (2012). Experiencias dendroclimáticas con árboles de ecosistemas contrastantes de Colombia. Revista colombiana de geografía cuadernos de geografía, 117-126.
Détienne, P. (1989). Appearance and periodicity of growth rings in some tropical woods. IAWA Bulletin, 10, 123-132.
Devall, M. S., Parresol, B. R., & Wright, S. J. (1995). Dendroecological analysis of Cordia alliodora, Pseudobombax septenatum and Annona spraguei in central Panama. IAWA Journal, 16 (4), 411-424.
Durán, O. J. (2012). Interpretación Dendrocronológica en Tres Especies Tropicales en una Selva Baja Caducifolia de Morelos. (Tesis en Maestro de Biología). División de Ciencias Biológicas y de la Salud de la Unidad Iztapalapa. Iztapalapa, 19-24.
Enfield, D. B. (1989). El Niño, past and present. Reviews of Geophysics, 27, 159-187.
Enquist, B. J., & Leffler, A. J. (2001). Long-term tree ring chronologies from sympatric tropical dry-forest trees: Individualistic responses to climate variation. Journal of Tropical Ecology, 17, 41-60.
Guisande, C. G., Vaamonde, A. L., & Barreiro, A. F. (2011). Tratamiento de datos con R, STATISTICA y SPSS. España, 978p.
Herrera, D. A. & Del Valle, J. I. (2011). Reconstrucción de los niveles del río Atrato con anillos de crecimiento de Prioria copaifera. Dyna, 78, 121-130.
Holmes, R. L. (1983). Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull, 43, 69-75.
IAWA COMMITTEE (Wheeler, E. A., Baas, P., & Gasson, P. Eds). (1989). IAWA list of microscopic features for hardwood identification. IAWA Journal, 10 (3), 219-233.
León, H. W. J. (2003). Anatomía de la madera y clave de identificación para especies forestales vedadas en Venezuela. Revista Forestal Venezolana, 53 (1), 51-62.
León, H. W. J. (2014). Anatomía de maderas de 108 especies de Venezuela. Pittieria, 66p.
Liegel, L. H. & Stead, J.W. (1990). Cordia alliodora (Ruiz and Pav.) Oken. En: Burns, R.M. & Honkala, B.H. (Tech. Coord.), Silvics of North America, vol. 2. Hardwoods. Agriculture Handbook 654. USDA Forest Service, Washington D.C. 270-77 p.
López, B. C., Sabaté, S., Garcia, C. A. & Rodríguez, R. (2005). Wood anatomy, description of annual rings, and responses to ENSO events of Prosopis pallida H: B. K., a wide-spread woody plant of arid and semi-arid lands of Latin America. Journal of Arid Environments, 61, 541-554.
López, L. & Villalba, R. (2011). Climate influences on the radial growth of Centrolobium microchaete, a valuable timber species from the tropical dry forest in Boliva. Biotropica, 43(1), 41-49.
Lozano, B. L. A. (2005). Patrones ecológicos de un relicto de bosque seco tropical ribereño en el CURN de la Universidad del Tolima Armero Guayabal. (Trabajo de maestría, Maestría en ciencias biológicas) Ibagué: Universidad del Tolima, 103 p.
Mariaux, A. (1967). Les cernes dans les bois tropicaux africains, nature et periodicité. Revue Bois et Forêts Des Tropiques, 113, 3-14.
Mendivelso, H. A., Camarero, J. J., Gutiérrez, E., & Zuidema, P. A. (2014). Time-dependent effects of climate and drought on tree growth in a Neotropical dry forest: Short-term tolerance vs. long-term sensitivity. Agricultural and Forest Meteorology, 188, 13-23.
Neelin, J. D. & Latif, M. (1998). El Niño Dynamics. Physics Today, 32-36.
Paredes-Villanueva, K., Sánchez-Salguero, R., Manzanedo, R. D., Quevedo-Sopepi, R., Palacios, G., & Navarro-Cerrillo, R. M. (2013). Growth rate and climatic response of Machaerium scleroxylon in a dry tropical forest in southeastern Santa Cruz, Bolivia. Tree-Ring Research, 69(2), 63-79
Pereyra, E. M. J., Guillen G. J. I., Morales M. S., & Arisméndiz, R. R. (2014). Potencialidad de Cedrela odorata (Meliaceae) para estudios dendrocronológicos en la selva central del Perú. Revista Biología Tropical, 62(2), 783-793.
Poveda, G., Vélez, J. I., Mesa, O., Hoyos, C. D., Mejía, J. F., Barco, O. J., & Correa, P. L. (2002). Influencia de fenómenos macroclimáticos sobre el ciclo anual de la hidrología colombiana: cuantificación lineal, no lineal y percentiles probabilísticos. Meteorología Colombiana, 6, 121-130.
R, Development Core Team. (2013). R. A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria.
Ramírez, J. A. & Del Valle, J. I. (2012). Local and global climate signals from tree rings of Parkinsonia praecox in La Guajira, Colombia. International Journal of Climatology, 32 (7), 1077-1088.
Ramírez, J. A., Molina E. C. & Bernal, M. (2010). Anillos anuales y clima en Rhizophora mangle L. de la Bahía de Cispatá, Colombia. Revista Facultad Nacional de Agronomía Medellín, 63, 5639-5650.
Rangel-Ch, J. O. & Arellano, H. (2008). El clima en el área del transecto Sumapaz (cordillera Oriental). En: Van der Hammen, T. (ed.). Estudios de Ecosistemas Tropandinos-Ecoandes 7. La cordillera Oriental, transecto de Sumapaz, 143-184. Cramer, J (BORNTRAEGER) Berlín-Stuttgaart.
Rangel-Ch, J. O. & Carvajal–Cogollo, J. E. (2009). Clima de la Serranía del Perijá. En: Rangel-Ch, J. O. (Ed.). Colombia Diversidad Biótica VIII. Media y baja montaña de la Serranía de Perijá, 3-49. Bogotá. Universidad Nacional de Colombia, Instituto de Ciencias Naturales.
Rangel-Ch, J. O. (2009). Síntesis sobre la biodiversidad de la Serranía de Perijá. En: Rangel-Ch, J.O. (ed.). Colombia Diversidad Biótica VIII. Media y baja montaña de la Serranía de Perijá, 677-692. Bogotá. Universidad Nacional de Colombia– Instituto de Ciencias Naturales–CORPOCESAR.
Rangel-Ch, J. O., Carvajal-Cogollo, J. E., Cortés-Duque, J., & Rivera-Díaz, O. (2009). Amenazas a la biota (vegetación, fauna, flora, ecosistemas) de la Serranía del Perijá. En: Rangel-Ch, J. O. (ed.). Colombia Diversidad Biótica VIII. Media y Baja Montaña de la Serranía de Perijá, 661–676. Bogotá Universidad Nacional de Colombia– Instituto de Ciencias Naturales–CORPOCESAR.
Rangel-Ch., J. O. (2012). (ed.). La vegetación – los bosques y el hombre con especial referencia al departamento del Cesar. Colombia Diversidad Biótica. Publicación Especial 5, 96. Bogotá. Grupo de Biodiversidad y Conservación, Instituto de Ciencias Naturales, Universidad Nacional de Colombia-CORPOCESAR.
Rodríguez, R., Mabres, A., Luckman, B., Evans, M., Masiokas, M., & Ektvedt, T. M. (2005). “El Niño” events recorded in dry forest species of the lowlands of northwest Peru. Dendrochronologia, 22, 181-186.
Roig, F. A. (2000). Dendrocronología en los bosques del Neotrópico: revisión y prospección futura. En: Roig, F. A. (2000) (ed.). Dendrocronología en América Latina, 307-355. Mendoza. Editorial de la Universidad Nacional de Cuyo.
Rosero, M. J. (2009). Dendrocronologia de árvores de mogno, Swietenia macrophylla King., Meliaceae, ocorrentes na floresta tropical Amazônica do Departamento de Madre de Dios, Peru. (Tesis Mastre em Recursos Florestais). Piracicaba. Universidade de São Paulo. 15-30.
Solíz, C., Villalba, R., Argollo, J., Morales, M. S., Christie, D .A., Moya, J., & Pacajes J. (2009). Spatio-temporal variations in Polylepis tarapacana radial growth across the Bolivian Altiplano during the 20th century. Palaeogeography, Palaeoclimatology, Palaeoecology, 281, 296-308.
Speer, J. H. (2010). Fundamentals of Tree-Ring Research. The University of Arizona Press. 324p.
Stahle, D. W., Mushove, P. T., Cleaveland, M. K., Roig, F., & Hayness, G. A. (1999). Management implications of annual growth ring in Pterocarpus angolensis from Zimbabwe. Forest Ecology and Management, 124, 217-229.
Stokes, M. A. & Smiley, T. L. (1968). An Introduction to Tree-Ring Dating. University of Chicago Press, Chicago, Illinois, 11.
Vetter, R. E. & Botosso, P. C. (1989). El Niño may affect growth behavior of Amazonian trees. GeoJournal, 19, 419-421.
Wigley, T. M. L., Briffa, K. R., & Jones, P. D. (1984). On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. Journal of Applied Meteorology and Climatology, 23, 201-213.
Worbes, M. & Junk, W. J. (1999). How old are tropical trees? The persistence of a myth. IAWA Journal, 20, 255-260.
Worbes, M. (1992). Occurrence of seasonal climate and tree-ring research in the tropics. Lundqua Report, 34, 338-342.
Worbes, M. (1995). How to measure growth dynamics in tropical trees: A review. IAWA Journal, 16, 337-351.
Worbes, M. (1999). Annual growth rings, rainfall-dependent growth and long-term growth patterns of tropical trees from the Caparo Forest Reserve in Venezuela. Journal of Ecology, 87, 391-403.
How to Cite
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Download Citation
 |
Briceño-J, A., Rangel-Ch, J. & Bogino, S. (2016). Estudio de los anillos de crecimiento de Cordia alliodora (Ruiz & Pav.) en Colombia. Colombia Forestal, 19(2), 95-107. |
Artículo de investigación
ESTUDIO DE LOS ANILLOS DE CRECIMIENTO DE Cordia alliodora (Ruiz & Pav.) EN COLOMBIA
Tree ring study of Cordia alliodora (Ruiz & Pav.) in Colombia
Ana María Briceño-J1, Jesús Orlando Rangel-Ch2 & Stella Marys Bogino3
1 Instituto de Ciencias Naturales, Universidad Nacional de Colombia. Bogotá, Colombia. ambricenoj@unal.edu.co. Autora encargada de la correspondencia.
2 Instituto de Ciencias Naturales, Universidad Nacional de Colombia. Bogotá, Colombia. jorangelc@gmail.com.
3 Departamento de Ciencias Agropecuarias, Universidad Nacional de San Luis. San Luis, Argentina. stellabogino@gmail.com.
Recepción: 12 de septiembre de 2015 / Aprobación: 14 de marzo de 2016
RESUMEN
La especie Cordia alliodora tiene un alto valor ecológico y económico en la formación de bosque seco tropical de Colombia. En este estudio se documentó la relación entre el monto de precipitación anual, temperatura media anual y el índice de oscilación del sur (SOI) con los anillos anuales de crecimiento de 28 individuos de C. alliodora con base en la aplicación de la metodología dendrocronológica estándar. C. alliodora registró anillos anuales de crecimiento demarcados por la porosidad semicircular y el parénquima terminal. La cronología construida cubrió un período de 70 años (1944-2013). El crecimiento radial de C. alliodora, representado a través de la función matemática de Gompertz, estuvo influenciado positivamente por el monto de precipitación anual y negativamente por la temperatura media anual. No se encontró influencia de los años con la presencia del fenómeno de El Niño (años secos). La cronología residual (1946-2013) mostró un ciclo de activación del crecimiento de 3.3 años vinculados al monto de precipitación anual y a la temperatura media anual. Los resultados sobre el índice de crecimiento muestran que C. alliodora alcanzaría el máximo crecimiento diamétrico a los 65 años con 205.5 cm.
Palabras clave: Cambio climático, Cordia alliodora, dendrocronología tropical, fenómenos de El Niño y La Niña, madera.
ABSTRACT
Cordia alliodora has a high ecological and economical value in the dry forest of Colombia. We documented the relationship between tree growth and climate variables (total annual precipitation, mean annual temperature) and the southern oscillation index (SOI) in this study. Standard dendrochronological methods were applied on 28 individuals. Presence of annual growth rings weas defined by semicircular porosity and terminal parenchyma. The tree-ring chronology spanned over the period 1944-2013 (70 years). Gompertz model was the mathematical function that best represented the growth. Radial growth was positively influenced by total annual precipitation and negatively related to mean annual temperature. There was no relation between tree growth and the SOI index (indicator of El Niño/La Niña events). A spectral density analysis of the residual tree-ring chronology showed a periodicity of 3.3 years cycle, similar to those cycles recorded on the precipitation and temperature series. According to the results, C. alliodora diameter growth would peak 205.5 cm at 65 years old.
Key words: Cordia alliodora, climate change, tropical dendrochronology, El Niño and La Niña phenomena, woods.
INTRODUCCIÓN
En la mayor parte del territorio de Colombia el fenómeno de El Niño se manifiesta con la disminución marcada de la precipitación, que en ocasiones puede significar la ausencia de lluvias en varios meses del año incluyendo enero, febrero, marzo, junio, julio y agosto. El fenómeno de La Niña es la condición contraria, con aumentos en la precipitación y la prevalencia de lluvia en casi todos los meses del año (Rangel-Ch. et al., 2009). La variabilidad del clima como las condiciones de sequía, inundaciones y eventos atípicos (fenómenos de El Niño y La Niña) pueden marcarse en los anillos de crecimiento de las especies arbóreas, confiriéndoles características especiales en el leño como disminución o ampliación del ancho del anillo de crecimiento, cambio en el tamaño de los vasos, presencia de canales traumáticos y tílides, entre otras características en la madera (Détienne, 1989; Vetter & Botosso, 1989; Worbes, 1992, 1995, 1999; Worbes & Junk, 1999; Stahle et al., 1999; Ramírez et al., 2010). Esta condición sugiere que los anillos son buenos indicadores de la variabilidad climática (Speer, 2010), como se ha documentado en diferentes especies arbóreas de las regiones tropicales (Worbes & Junk, 1999; Roig, 2000; Enquist & Leffler, 2001; Solíz et al., 2009; Briceño & Arévalo, 2010; Herrera & del Valle, 2011; López & Villalba, 2011; Ramírez & del Valle, 2012; Pereyra et al., 2014).
En los bosques secos tropicales los estudios de los anillos de crecimiento han documentado los eventos de sequía e inundaciones (Rodríguez et al., 2005). El bosque seco tropical ha sido transformado para constituir asentamientos humanos y su uso excesivo ha fragmentado buena parte de la superficie boscosa original (Rangel-Ch., 2012). Los remanentes actuales se encuentran muy amenazados por la ampliación de la frontera agrícola, la minería de alto impacto y otros efectos asociados con el cambio climático, como El Niño (Rangel-Ch., 2012) y la disponibilidad del agua (Poveda et al., 2002; Rangel-Ch. et al., 2009). Por lo tanto, como consecuencia de esta intervención humana los servicios ambientales que estos bosques brindan son afectados.
Cordia alliodora, familia Boraginaceae, comúnmente conocido como laurel, es una especie de hoja caduca y de rápido crecimiento con amplia distribución geográfica en las islas del Caribe y desde el norte de México a través de América Central hasta Paraguay, el sur de Brasil y el norte de Argentina (CATIE, 1994). Es una especie dominante en los bosques de tierras bajas y es muy utilizada en la industria forestal (Liegel & Stead, 1990). En los bosques tropicales secos de Panamá se demostró la anualidad en la formación de los anillos de crecimiento de C. alliodora y su relación con las variables climáticas (Devall et al., 1995).
Considerando: a) la importancia del impacto de fenómenos climáticos globales como El Niño y La Niña sobre los bosques secos tropicales; b) la factibilidad de la demarcación de los anillos de crecimiento de las especies leñosas que crecen en estos ambientes y cuyo ancho fluctúa con la variabilidad climática; c) el valor de estos bosques tropicales secos y la amenaza que se cierne sobre ellos por alteraciones climáticas y antrópicas; d) la importancia de C. alliodora como especie leñosa estructural de estos bosques, tanto fuera como dentro de Colombia; y e) la falta de información sobre la respuesta de esta especie a las variables climáticas en Colombia. Se propuso el siguiente objetivo al presente trabajo: determinar la relación entre las variaciones anuales de precipitación, temperatura e índice SOI con el crecimiento radial de C. alliodora en el bosque tropical seco de Colombia.
MATERIALES Y MÉTODOS
Sitio de muestreo
El muestreo se realizó en relictos de bosque seco tropical, en el Centro Universitario Granja de Armero de la Universidad del Tolima, ubicado en el norte del departamento del Tolima(Colombia), sobre el valle del río Magdalena (5° 0’ 0.6’’N, 74° 54’ 27.77’’W). En la composición florística de estos bosques secos C. alliodora es una especie arbórea dominante. Las áreas con bosque original fueron transformadas casi en su totalidad para implantar cultivos de arroz, algodón y maíz (Lozano, 2005). Además, la ganadería extensiva ha reducido su extensión a pequeños fragmentos. La textura del suelo es franco-arenosa, el pH es ácido, la fertilidad es baja y presenta gran compactación (Lozano, 2005).
El promedio histórico de la precipitación total anual en la zona de estudio es de 1793.17 mm. El régimen de distribución de las lluvias es de tipo bimodal-tetraestacional. Los períodos lluviosos se presentan entre abril y mayo y entre septiembre y noviembre, siendo octubre el mes más lluvioso. Los períodos con menor precipitación se presentan desde diciembre hasta marzo y desde junio hasta agosto. Enero es el mes más seco, con un promedio histórico de 81.02 mm (Rangel-Ch. et al., 2009). El promedio histórico de las temperaturas medias anuales es de 28.23º C. Los períodos cálidos van desde enero hasta febrero y desde junio hasta septiembre, siendo agosto el mes con mayor temperatura media (28.80ºC). Los períodos con temperaturas menores al promedio histórico van desde marzo hasta junio y desde octubre hasta diciembre (figura 1).
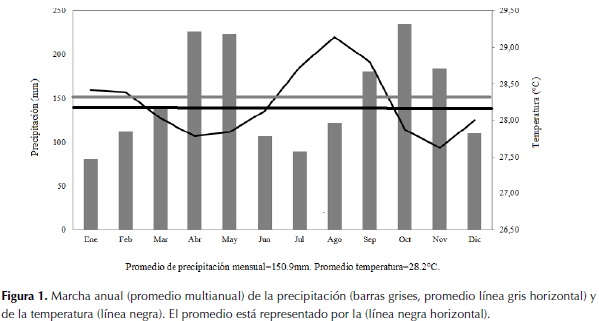
El tipo de clima según Thornthwaite (adaptación de Rangel-Ch & Arellano, 2008) se clasifica como C1RD semiseco, con poca o ninguna deficiencia de agua en la época seca y poco o nada de superávit de agua. En años húmedos, como por ejemplo 1999, el valor de la precipitación estuvo por encima de la evapotranspiración potencial (ETP) en los meses de febrero, abril, junio, agosto, septiembre, octubre, noviembre y diciembre, es decir, hay superávit de agua. En años secos, como por ejemplo 1995, los valores de evapotranspiración potencial (ETP) se encontraron por encima de los valores de precipitación exceptuando julio, agosto y octubre, e indicaron una deficiencia de agua.
Toma de muestras en campo
Se colectaron 28 rodajas de C. alliodora, 18 de árboles que crecían en áreas naturales y 10 de rodales de plantación de ocho años de edad, ubicados en el Centro Universitario Granja de Armero de la Universidad del Tolima. Las rodajas fueron secadas durante ocho días a temperatura ambiente, posteriormente, para la correcta demarcación de los anillos de crecimiento, estas se lijaron con una pulidora manual (granulometrías desde 36, 50, 80, 120, 1000 hasta 1500) hasta visualizar completamente los elementos celulares del leño (Stokes & Smiley, 1968).
Datación de los anillos de crecimiento
Para comprobar la anualidad de los anillos de crecimiento se siguió el método de Mariaux (1967), que consiste en hacer una herida en el cambium de las especies y verificar después de un tiempo la formación de los anillos de crecimiento (Rosero, 2009; Durán, 2012). En el fuste de los árboles de C. alliodora se hizo una herida con el barreno de incrementos de Pressler en seis individuos en el año 2008, que se revisaron en el año 2009. Para la descripción anatómica se cortaron 10 secciones transversales entre 20 y 25mm de espesor usando un micrótomo Marca Leitz modelo 1208 y guiados por el protocolo de la Asociación Internacional de Anatomistas de la Madera (IAWA, 1989).
Construcción de la cronología
Para la medición de los anillos se utilizó una lupa estereoscópica Olympus SZ61 (0.9 a 4 X) y una mesa de medición Velmex TA Unislide modelo TA4021 H–S6 con una precisión de 0.001mm. Para verificar errores en la datación de los anillos de crecimiento se utilizó el programa COFECHA (Holmes, 1983). Para la estandarización de los anillos de crecimiento se empleó el programa ARSTAN (Cook, 1985), que transforma las mediciones de los anillos en índices de crecimiento. Este programa reduce la variación interanual causada por la tendencia biológica y los disturbios endógenos, además maximiza el porcentaje de varianza común en el ancho de los anillos entre las series. Las auto-correlaciones miden la dependencia estadística entre series y árboles diferentes. A este respecto se evaluó el intervalo común óptimo que se refiere al mayor período de tiempo con el máximo número de series de índices de crecimiento; la correlación media entre radios, entre árboles, dentro de árboles y de cada radio con la cronología media, la relación señal-ruido que es la robustez de la señal común observada entre los árboles que componen la cronología y la señal expresada de la población (EPS) que es la concordancia con la cronología poblacional, que cuantifica el grado de representación de la cronología obtenida (Speer, 2010). El presente estudio utilizó un segmento de estandarización de 50 años.
Modelos de crecimiento
Para obtener la función matemática que representa el crecimiento de la especie se utilizaron los modelos de regresión para variables dependientes cuantitativas, funciones no lineales (Clench, Exponencial negativa, Saturación, Racional, Logístico y Gompertz) con el paquete estadístico R (R Development Core Team, 2013). Para seleccionar el mejor modelo se tuvieron en cuenta la normalidad de los residuos, el mayor valor del coeficiente de determinación, el menor número de error estándar residual y el menor número de iteraciones (Guisande et al., 2011).
Relación clima-crecimiento
Los registros climáticos de la precipitación total mensual desde 1964 hasta 2013 y la temperatura media mensual desde 1987 hasta 2013 fueron suministrados por el Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia (IDEAM) para la estación Granja-Armero (05°01’N y 74°54´W). El índice de oscilación del sur (SOI) desde 1964 hasta 2013 fue descargado de la página web de la National Oceanic and Atmospheric Administration, USA del National Weather Service. Los valores positivos de SOI indican eventos de La Niña y los valores negativos indican eventos de El Niño (Enfield, 1989; Neelin & Latif, 1998).
Para el análisis de la relación del clima con el crecimiento de la especie se utilizó un contraste de independencia y la asociación de las variables cuantitativas mediante la correlación múltiple de Pearson. Este coeficiente indica el grado de asociación entre varias variables independientes. Los análisis anteriores se realizaron con el programa R comparando cada variable ambiental (precipitación total mensual, temperatura media y el índice de SOI) con el índice de crecimiento residual de los anillos por presentar los mejores resultados de correlación (R Development Core Team, 2013).
Se amplió el registro de la precipitación anual entre 1946 y 1963 con base en los modelos de regresión simple (lineal, logarítmico, curva-S, potencial, exponencial, cuadrática, cúbica e inversa). Por lo tanto, se utilizó el monto de precipitación anual y el índice de crecimiento residual. De los anteriores modelos de regresión simple se escogió el mejor con base en el mayor valor del coeficiente de determinación y el menor error estándar residual. Los principios para aplicar la regresión fueron: a) la distribución normal de los residuos del modelo; b) la homocedasticidad en los residuos, esto indica que la varianza de los mismos debe ser constante; y c) la ausencia de autocorrelación en la serie de los residuos.
Para ampliar el registro de la temperatura media anual entre 1946 y 1986 se utilizó un modelo de regresión múltiple, empleado el método paso a paso (stepwise) que emplea el criterio de información de Akaike para definir las variables que se excluyen del modelo de regresión, ya que no contribuyen a la explicación del índice de crecimiento. Las variables incluidas fueron: el monto de precipitación anual y la temperatura media anual. En este modelo se incluyó la precipitación anual porque confiere aumentar el coeficiente de determinación y mejorar el modelo de regresión múltiple. Este análisis debe cumplir los mismos requisitos que la regresión simple y no debe existir multicolinealidad entre las variables (Guisande et al., 2011).
Para analizar el comportamiento de ciclos en la serie entre los años 1946 y 2013 se utilizó el análisis espectral, el cual es un procedimiento utilizado para identificar comportamientos periódicos en una serie temporal y para estudiar la existencia de ciclos o variaciones regulares cuyo período es generalmente superior a un año. El procedimiento descompone el conjunto de todas las observaciones de la serie en componentes periódicos de diferentes frecuencias y permite identificar las más relevantes (Guisande et al., 2011). En este análisis se utilizó el programa R. Las variables utilizadas fueron el índice de crecimiento residual, el monto de precipitación anual y la temperatura media mensual. No se incluyó en el análisis el índice SOI, porque se pretende averiguar la ciclicidad de períodos húmedos y secos, temperaturas máximas en la serie.
RESULTADOS
Anatomía de Cordia alliodora
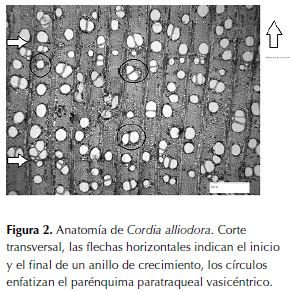
En el corte transversal se observa una porosidad semicircular, poros solitarios con agrupaciones racemiformes de dos, tres a cuatro poros. Posee bandas de parénquima axial marginal, finas, hasta de tres células de ancho, parénquima paratraqueal vasicéntrico, aliforme de ala corta (figura 2). Los anillos de crecimiento están definidos por la porosidad semicircular y el parénquima marginal y la disminución del tamaño de las fibras.
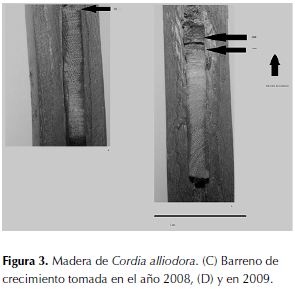
Presenta anillos de crecimiento de frecuencia anual, de acuerdo con el resultado de las heridas anuales en el fuste de C. alliodora en los años 2008 y 2009 (figuras 3a y 3b).
Datación de los anillos de crecimiento
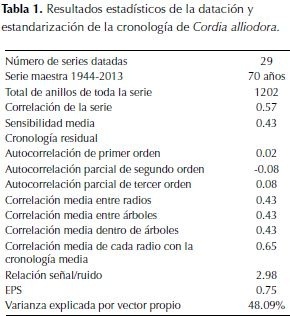
En varias secciones de las rodajas se presentaron dificultades para la datación debido al crecimiento irregular, especialmente la anchura de los anillos alrededor de la sección transversal. Los resultados de la verificación de la datación de 70 años (1944-2013) con el programa COFECHA mostraron una correlación media de 0.57 para 29 series, la sensibilidad media fue de 0.43. Los resultados de la estandarización (tabla 1) según la señal expresada de la población (EPS) oscila entre 0.44 y 0.94 el valor medio es 0.757. Los valores bajos corresponden a pocos individuos en el período analizado (figura 4).
Crecimiento radial acumulado
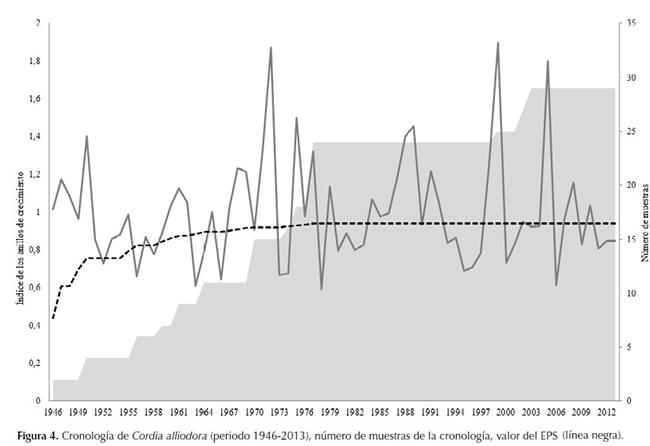
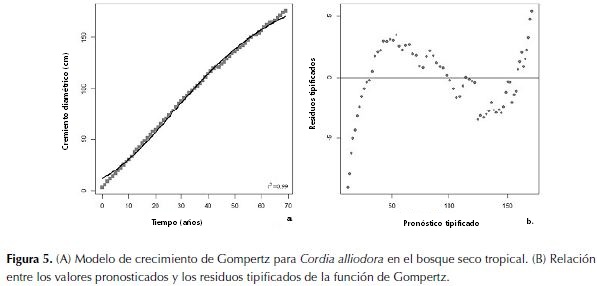
La función que mejor representó el crecimiento de C. alliodora fue la curva de Gompertz (figura 5a). Los residuos del modelo fueron normales (p=0.46) (figura 5b). El coeficiente de determinación fue 0.99, el error estándar residual fue de 2.85 y el número de iteraciones fue 7. La ecuación (1) representa el crecimiento de la especie:

Relación clima-crecimiento
Correlación entre las variables ambientales y el índice de crecimiento
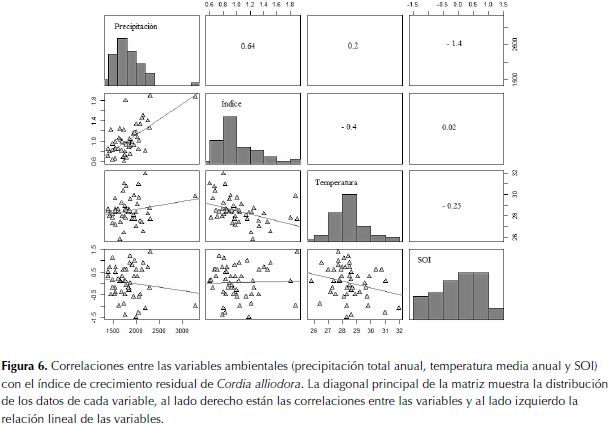
La correlación entre el monto de precipitación anual, la temperatura media anual y el SOI con el índice de crecimiento residual (figura 6) mostró los siguientes resultados: a)entre el índice de crecimiento residual y el monto de precipitación anual fue 0.64 (p= <0.001) (valor más alto del análisis) (período de la correlación 1964-2013); b) entre el índice de crecimiento residual y la temperatura media anual, el valor fue negativo -0.4 (p= 0.003) (período de la correlación 1987-2013); c) entre el índice de crecimiento residual y el SOI fue de 0.02 (p= 0.872) (período de la correlación 1964-2013); d) entre el monto de precipitación anual y la temperatura media anual el valor fue de 0.2 (p= 0.172) (período de la correlación 1987-2013); e) entre el monto de precipitación anual y el SOI fue de -1.4 (p= 0.34) (período de la correlación 1964-2013); f) entre la temperatura media anual y el SOI hubo una correlación negativa de -0.25 (p= 0.074) (período de la correlación 1987-2013).
Modelo de regresión lineal entre la precipitación y el índice de crecimiento
El modelo que expresa de manera más significativa la regresión lineal entre el monto de precipitación anual (1964-2013) y el índice de crecimiento es la ecuación (2):

El error estándar residual=0.25. Este modelo cumple la premisa de la normalidad en los residuos probado con Kolmogorov-Smirnov-Lilliefors (p=0.36), se cumple el principio de homocedasticidad según el test de Breusch-Pagan (p=0.73) y no hay auto-correlación probada con el test de Durbin-Watson (p=0.36).
Modelo de regresión múltiple entre las variables ambientales y el índice de crecimiento
El mejor modelo de regresión múltiple entre la temperatura media anual (1964-2013), el monto de precipitación anual (1964-2013) y el índice de crecimiento se obtuvo por el método stepwise, ecuación 3:
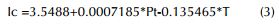
Dónde: Ic es el índice de crecimiento residual de C. alliodora, Pt el monto de precipitación anual y T la temperatura media anual, los coeficientes son significativos para el modelo (p>0.05).
Este modelo explica un 57.3% de la varianza observada en el índice de crecimiento de C. alliodora (r2=0.57), cumple con normalidad en los residuos p>0.05 (p=0.05), se cumple el principio de homocedasticidad (test de Breusch-Pagan, p=0.56), no hay auto-correlación probada con el test de Durbin-Watson (p=0.41) y la multi-colinealidad en el modelo del monto de precipitación anual y la temperatura media anual fue (1.03 y 1.02) respectivamente.
Ciclos en la serie
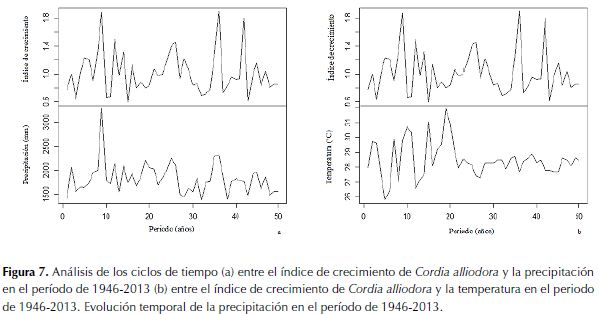
En la serie de tiempo de 50 años (período de datos instrumentales 1964-2013) hay una variación cíclica entre índice de crecimiento de C. alliodora y la precipitación de 3.3 años, se muestran los valores más altos en el índice de crecimiento y los años más lluviosos (figura 7a). La coherencia entre estas dos variables es de 2.52, (valores cercanos a 1 indican relaciones lineales entre las variables). Por lo tanto, existe coherencia entre las dos variables. El valor del estadígrafo de retardo entre las dos variables fue de 0.2. El crecimiento de C. alliodora después de la manifestación de un período húmedo se activa a los dos meses. Con la temperatura (figura 7b) se detectó el mismo tiempo en el ciclo (3.3 años) entre la manifestación de períodos muy cálidos, el valor de coherencia entre las dos variables fue de 0.88. Entre las dos variables hay un retraso en la coincidencia de 2.9 años.
DISCUSIÓN
Las características anatómicas de las muestras de C. alliodora son similares a las registradas por León (2003; 2014) en la familia Boraginaceae en Venezuela. Las agrupaciones de los vasos que se encontraron en los cortes transversales son una de las estrategias utilizadas por las plantas que crecen en el bosque seco para asegurar la conducción de agua. Igualmente, la formación del parénquima axial, encontrado en los cortes transversales le permite a la especie reservar agua y almidones para la época seca (López et al., 2005). La formación anual de los anillos de crecimiento, detectado por el método de Mariaux (1967), confirma los hallazgos de Devall et al., (1995), quienes habían documentado el carácter anual de los anillos de crecimiento para esta especie en los bosques secos de Panamá.
La datación de los anillos de crecimiento de C. alliodora demuestra que es posible fechar los anillos y que existe sincronía en el crecimiento según los resultados de la verificación con los programas COFECHA y ARSTAN. Los valores de la inter-correlación de la serie (0.57), están dentro de los límites de variación conocidos para estudios dendrocronológicos. A este respecto, Rodríguez et al. (2005), en una serie de 38 años (1964–2001) encontraron en diferentes bosques secos del Perú para Bursera graveolens (Kunth) Triana y Planchon, valores de correlación entre 0.53–0.71. Los estadísticos de la señal expresada de la población (EPS) ≥ 0.85 se consideran como aceptables para especies tropicales (Wigley et al., 1984). Valores de EPS >0.85 fueron mostrados en dos cronologías de Centrolobium microchaete (Benth.) H.C. Lima, en dos bosques secos de Bolivia con un número similar de muestras al del nuestro estudio (29 y 38) (López & Villalba, 2011).
De acuerdo con el modelo de Gompertz, el crecimiento diamétrico máximo de la especie se alcanzaría a los 65 años con 205.5 cm. Otros modelos de crecimiento para las especies tropicales en el bosque seco tropical se han ajustado a curvas sigmoidales como es el caso de Machaerium scleroxylon Tul., en Bolivia, que según su modelo de crecimiento alcanzan el máximo diámetro (40cm) a los 140 años (Paredes-Villanueva et al., 2013).
Las variables más relacionadas con el índice de crecimiento anual de la cronología residual, fueron la precipitación total anual (relación positiva) y la temperatura media anual (relación negativa). Estos resultados coinciden con estudios previos donde la precipitación estuvo asociada positivamente con el crecimiento de diferentes especies tropicales como Capparis indica (L.) Druce y Genipa americana L., en Costa Rica (Enquist & Leffler, 2001), Cordia apurensis en Venezuela (Worbes, 1999), Prosopis pallida (Willd.) Kunth en Perú (López et al., 2005), y seis especies de árboles del bosque seco tropical de Bolivia (Centrolobium microchaete, Acosmium cardenasii, Caesalpinia pluviosa, Aspidosperma tomentosum, Zeyheria tuberculosa y Anadenanthera macrocarpa) (Mendivelso et al., 2014).
Los resultados muestran que el crecimiento C. alliodora se relacionó con el monto de precipitación anual y con la temperatura y no con el SOI, estas relaciones son probables porque el crecimiento de la especie está influenciado por las condiciones locales de precipitación y temperatura. Resultados similares en un bosque con precipitación media anual de 1700 mm en Venezuela mostraron que el crecimiento de Terminalia guyanensis Eichler no tuvo una relación estadísticamente significativa con el fenómeno de El Niño (Worbes, 1999). De la misma forma, comparaciones de la precipitación y el índice de crecimiento muestran una relación estadística positiva como en la costa noreste de Perú (5°S, 80°W) donde se encontró mayor crecimiento radial en Bursera graveolens (Kunth) Triana y Planch (Rodríguez et al., 2005). Resultados similares mostró Prosopis pallida (Willd.) Kunth., donde en los años de mayor precipitación hubo mayor crecimiento radial (López et al., 2005).
Según los resultados, hay un ciclo de 3.3 años en la activación del crecimiento de C. alliodora, que se manifiesta igualmente con el monto de precipitación anual y la temperatura. La activación del crecimiento fue mayor en los años con mayores valores de precipitación. Aunque son muy escasas las contribuciones sobre la ciclicidad en la distribución de las variables climáticas en nuestro país, cabe mencionar el aporte de Rangel-Ch & Carvajal-Cogollo (2009), quienes para varias localidades de la serranía del Perijá (Cesar), mencionaron una ciclicidad de 10 años de duración para la variación entre períodos secos y húmedos, periodo más largo que el encontrado en esta investigación.
CONCLUSIONES
El estudio dendrocronológico de C. alliodora (anualidad de los anillos, datación, evaluación de las mediciones) permitió establecer una cronología de 70 años. A partir de la medición de los anillos se realizaron análisis de regresión lineal para ampliar los registros del monto de precipitación anual en 18 años entre 1946 y 1964 y de regresión múltiple para ampliar los registros de temperatura promedio anual en 41 años entre 1946-1986.
El índice de crecimiento de los anillos de C. alliodora mostró una relación positiva con el monto de precipitación anual (período de la correlación 1964-2013) y negativa con la temperatura media anual) (período de la correlación 1987-2013). Y poca relación estadística con el índice SOI (período de la correlación 1964-2013).
De acuerdo con los resultados sobre el índice de crecimiento, C. alliodora alcanzaría el máximo crecimiento diamétrico a los 65 años con 205.5 cm. Ante la deforestación incontrolada esta cifra debería ser tomada en cuenta para impulsar programas de conservación de los parches de bosque seco tropical que aún persisten.
Con este estudio se amplían los registros dendroclimaticos para C. alliodora y se consolida su potencial dendrocronológico y la utilidad del análisis del crecimiento de sus anillos en reconstrucciones climáticas.
AGRADECIMIENTOS
A la Universidad Nacional de Colombia, especialmente a la Facultad de Ciencias, Instituto de Ciencias Naturales–Departamento de Biología. A Colciencias por el apoyo en la investigación. Al laboratorio de maderas de la Universidad del Tolima por permitir realizar los cortes anatómicos.
REFERENCIAS BIBLIOGRÁFICAS
Briceño-J, A. M., & Arévalo, F. R. (2010). Potencial dendroclimático de la madera de cinco especies del bosque seco tropical de Armero–Tolima. (Trabajo de maestría, Maestría en ciencias biológicas) Ibagué: Universidad del Tolima. 56p.
CATIE (Centro agronómico tropical de investigación y enseñanza). (1994). Laurel Cordia alliodora (Ruiz &Pav.) Oken Boraginaceae un árbol de uso múltiple. Colección materiales de extensión. Turrialba, Costa Rica. 476p.
Cook, E. R. (1985). A time series analysis approach to tree-ring standardization. (Ph. D. thesis) Tucson. University of Arizona. 89p.
Détienne, P. (1989). Appearance and periodicity of growth rings in some tropical woods. IAWA Bulletin, 10, 123-132.
Devall, M. S., Parresol, B. R., & Wright, S. J. (1995). Dendroecological analysis of Cordia alliodora, Pseudobombax septenatum and Annona spraguei in central Panama. IAWA Journal, 16 (4), 411-424.
Durán, O. J. (2012). Interpretación Dendrocronológica en Tres Especies Tropicales en una Selva Baja Caducifolia de Morelos. (Tesis en Maestro de Biología). División de Ciencias Biológicas y de la Salud de la Unidad Iztapalapa. Iztapalapa, 19-24.
Enfield, D. B. (1989). El Niño, past and present. Reviews of Geophysics, 27, 159-187.
Enquist, B. J., & Leffler, A. J. (2001). Long-term tree ring chronologies from sympatric tropical dry-forest trees: Individualistic responses to climate variation. Journal of Tropical Ecology, 17, 41-60.
Guisande, C. G., Vaamonde, A. L., & Barreiro, A. F. (2011). Tratamiento de datos con R, STATISTICA y SPSS. España, 978p.
Herrera, D. A. & Del Valle, J. I. (2011). Reconstrucción de los niveles del río Atrato con anillos de crecimiento de Prioria copaifera. Dyna, 78, 121-130.
Holmes, R. L. (1983). Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull, 43, 69-75.
IAWA COMMITTEE (Wheeler, E. A., Baas, P., & Gasson, P. Eds). (1989). IAWA list of microscopic features for hardwood identification. IAWA Journal, 10 (3), 219-233.
León, H. W. J. (2003). Anatomía de la madera y clave de identificación para especies forestales vedadas en Venezuela. Revista Forestal Venezolana, 53 (1), 51-62.
León, H. W. J. (2014). Anatomía de maderas de 108 especies de Venezuela. Pittieria, 66p.
Liegel, L. H. & Stead, J.W. (1990). Cordia alliodora (Ruiz and Pav.) Oken. En: Burns, R.M. & Honkala, B.H. (Tech. Coord.), Silvics of North America, vol. 2. Hardwoods. Agriculture Handbook 654. USDA Forest Service, Washington D.C. 270-77 p.
López, B. C., Sabaté, S., Garcia, C. A. & Rodríguez, R. (2005). Wood anatomy, description of annual rings, and responses to ENSO events of Prosopis pallida H: B. K., a wide-spread woody plant of arid and semi-arid lands of Latin America. Journal of Arid Environments, 61, 541-554.
López, L. & Villalba, R. (2011). Climate influences on the radial growth of Centrolobium microchaete, a valuable timber species from the tropical dry forest in Boliva. Biotropica, 43(1), 41-49.
Lozano, B. L. A. (2005). Patrones ecológicos de un relicto de bosque seco tropical ribereño en el CURN de la Universidad del Tolima Armero Guayabal. (Trabajo de maestría, Maestría en ciencias biológicas) Ibagué: Universidad del Tolima, 103 p.
Mariaux, A. (1967). Les cernes dans les bois tropicaux africains, nature et periodicité. Revue Bois et Forêts Des Tropiques, 113, 3-14.
Mendivelso, H. A., Camarero, J. J., Gutiérrez, E., & Zuidema, P. A. (2014). Time-dependent effects of climate and drought on tree growth in a Neotropical dry forest: Short-term tolerance vs. long-term sensitivity. Agricultural and Forest Meteorology, 188, 13-23.
Neelin, J. D. & Latif, M. (1998). El Niño Dynamics. Physics Today, 32-36.
Paredes-Villanueva, K., Sánchez-Salguero, R., Manzanedo, R. D., Quevedo-Sopepi, R., Palacios, G., & Navarro-Cerrillo, R. M. (2013). Growth rate and climatic response of Machaerium scleroxylon in a dry tropical forest in southeastern Santa Cruz, Bolivia. Tree-Ring Research, 69(2), 63-79
Pereyra, E. M. J., Guillen G. J. I., Morales M. S., & Arisméndiz, R. R. (2014). Potencialidad de Cedrela odorata (Meliaceae) para estudios dendrocronológicos en la selva central del Perú. Revista Biología Tropical, 62(2), 783-793.
Poveda, G., Vélez, J. I., Mesa, O., Hoyos, C. D., Mejía, J. F., Barco, O. J., & Correa, P. L. (2002). Influencia de fenómenos macroclimáticos sobre el ciclo anual de la hidrología colombiana: cuantificación lineal, no lineal y percentiles probabilísticos. Meteorología Colombiana, 6, 121-130.
R, Development Core Team. (2013). R. A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria.
Ramírez, J. A. & Del Valle, J. I. (2012). Local and global climate signals from tree rings of Parkinsonia praecox in La Guajira, Colombia. International Journal of Climatology, 32 (7), 1077-1088.
Ramírez, J. A., Molina E. C. & Bernal, M. (2010). Anillos anuales y clima en Rhizophora mangle L. de la Bahía de Cispatá, Colombia. Revista Facultad Nacional de Agronomía Medellín, 63, 5639-5650.
Rangel-Ch, J. O. & Arellano, H. (2008). El clima en el área del transecto Sumapaz (cordillera Oriental). En: Van der Hammen, T. (ed.). Estudios de Ecosistemas Tropandinos-Ecoandes 7. La cordillera Oriental, transecto de Sumapaz, 143-184. Cramer, J (BORNTRAEGER) Berlín-Stuttgaart.
Rangel-Ch, J. O. & Carvajal–Cogollo, J. E. (2009). Clima de la Serranía del Perijá. En: Rangel-Ch, J. O. (Ed.). Colombia Diversidad Biótica VIII. Media y baja montaña de la Serranía de Perijá, 3-49. Bogotá. Universidad Nacional de Colombia, Instituto de Ciencias Naturales.
Rangel-Ch, J. O. (2009). Síntesis sobre la biodiversidad de la Serranía de Perijá. En: Rangel-Ch, J.O. (ed.). Colombia Diversidad Biótica VIII. Media y baja montaña de la Serranía de Perijá, 677-692. Bogotá. Universidad Nacional de Colombia– Instituto de Ciencias Naturales–CORPOCESAR.
Rangel-Ch, J. O., Carvajal-Cogollo, J. E., Cortés-Duque, J., & Rivera-Díaz, O. (2009). Amenazas a la biota (vegetación, fauna, flora, ecosistemas) de la Serranía del Perijá. En: Rangel-Ch, J. O. (ed.). Colombia Diversidad Biótica VIII. Media y Baja Montaña de la Serranía de Perijá, 661–676. Bogotá Universidad Nacional de Colombia– Instituto de Ciencias Naturales–CORPOCESAR.
Rangel-Ch., J. O. (2012). (ed.). La vegetación – los bosques y el hombre con especial referencia al departamento del Cesar. Colombia Diversidad Biótica. Publicación Especial 5, 96. Bogotá. Grupo de Biodiversidad y Conservación, Instituto de Ciencias Naturales, Universidad Nacional de Colombia-CORPOCESAR.
Rodríguez, R., Mabres, A., Luckman, B., Evans, M., Masiokas, M., & Ektvedt, T. M. (2005). “El Niño” events recorded in dry forest species of the lowlands of northwest Peru. Dendrochronologia, 22, 181-186.
Roig, F. A. (2000). Dendrocronología en los bosques del Neotrópico: revisión y prospección futura. En: Roig, F. A. (2000) (ed.). Dendrocronología en América Latina, 307-355. Mendoza. Editorial de la Universidad Nacional de Cuyo.
Rosero, M. J. (2009). Dendrocronologia de árvores de mogno, Swietenia macrophylla King., Meliaceae, ocorrentes na floresta tropical Amazônica do Departamento de Madre de Dios, Peru. (Tesis Mastre em Recursos Florestais). Piracicaba. Universidade de São Paulo. 15-30.
Solíz, C., Villalba, R., Argollo, J., Morales, M. S., Christie, D .A., Moya, J., & Pacajes J. (2009). Spatio-temporal variations in Polylepis tarapacana radial growth across the Bolivian Altiplano during the 20th century. Palaeogeography, Palaeoclimatology, Palaeoecology, 281, 296-308.
Speer, J. H. (2010). Fundamentals of Tree-Ring Research. The University of Arizona Press. 324p.
Stahle, D. W., Mushove, P. T., Cleaveland, M. K., Roig, F., & Hayness, G. A. (1999). Management implications of annual growth ring in Pterocarpus angolensis from Zimbabwe. Forest Ecology and Management, 124, 217-229.
Stokes, M. A. & Smiley, T. L. (1968). An Introduction to Tree-Ring Dating. University of Chicago Press, Chicago, Illinois, 11.
Vetter, R. E. & Botosso, P. C. (1989). El Niño may affect growth behavior of Amazonian trees. GeoJournal, 19, 419-421.
Wigley, T. M. L., Briffa, K. R., & Jones, P. D. (1984). On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. Journal of Applied Meteorology and Climatology, 23, 201-213.
Worbes, M. & Junk, W. J. (1999). How old are tropical trees? The persistence of a myth. IAWA Journal, 20, 255-260.
Worbes, M. (1992). Occurrence of seasonal climate and tree-ring research in the tropics. Lundqua Report, 34, 338-342.
Worbes, M. (1995). How to measure growth dynamics in tropical trees: A review. IAWA Journal, 16, 337-351.
Worbes, M. (1999). Annual growth rings, rainfall-dependent growth and long-term growth patterns of tropical trees from the Caparo Forest Reserve in Venezuela. Journal of Ecology, 87, 391-403.




License
Colombia Forestal retains the patrimonial rights (copyright) of the published works, and favors and allows the reuse of the same under the Creative Commons Attribution-ShareAlike 4.0 International license, so they can be copied, used, disseminated, transmitted and exhibited publicly, provided that:
You acknowledge the credits of the work in the manner specified by the author or licensor (but not in a way that suggests that you have their support or that they endorse your use of their work).