
DOI:
https://doi.org/10.14483/2256201X.20122Publicado:
01-01-2024Número:
Vol. 27 Núm. 1 (2024): Enero-junioSección:
Artículos de investigación científica y tecnológicaLa fracción no renovable de la biomasa como indicador de degradación forestal: Colombia como estudio de caso
The Fraction of Non – Renewable Biomass as an Indicator of Forest Degradation: Colombia as a Study Case
Palabras clave:
Biomass, emissions, fuelwood, REDD+, renewability (en).Palabras clave:
Biomasa, emisiones, leña, REDD+, renovabilidad (es).Descargas
Referencias
Ahrends, A., Bulling, M., Platts, P., Swetnam, R., Ryan, C., Doggart, N., Hollingsworth, P. M, Marchant, R., Balmford, A., Harris, D. J., Gross-Camp, N., Sumbi, P., Munishi, P., Madoffe, S., Mhoro, B., Leonard, C., Bracebridge, C., Doody, K., … Burgess, N. D. (2021). Detecting and predicting forest degradation: A comparison of ground surveys and remote sensing in Tanzanian forests. Plants People Planet, 3, 268-281. https://doi.org/10.1002/ppp3.10189 DOI: https://doi.org/10.1002/ppp3.10189
Architecture for REDD+ Transactions (2021). The REDD+ environmental excellence standard (TREES). ART Secretariat, Winrock International.
Aristizábal, J. (2015). Determinación de la fracción no renovable de la biomasa (fNRB) como instrumento de selección de áreas potenciales para la diseminación de estufas mejoradas. En R. Gómez, J. Aristizábal & L. M. Cardenas (Eds.), Estufas eficientes como contribución a la calidad de vida, al uso eficiente de la energía y la reducción de emisiones de GEI en áreas rurales de Antioquia y Santander, Colombia (pp. 51-72). Fundación Natura, Instituto Interamericano de Cooperación para la Agricultura (IICA).
Armenteras, D., González, T., Meza, M., & Vélez, J. (2018). Causas de la degradación de bosques en Colombia. En D. Armenteras, T. Gonzalez, M. Meza, J. Ramírez, E. Cabrera, G. Galindo & A. Yepes (Eds.), Causas de degradación forestal en Colombia: una primera aproximación (pp. 7-20). Universidad Nacional de Colombia, IDEAM, Programa ONU-REDD.
Bailis, R., Drigo, R., Ghilardi, A., & Masera, O. (2015). The carbon footprint of traditional woodfuels. Nature Climate Change, 5, 266-272. https://doi.org/10.1038/NCLIMATE2491 DOI: https://doi.org/10.1038/nclimate2491
Bailis, R., Wang, Y., Drigo, R., Ghilardi, A., & Masera, O. (2017). Getting the numbers right: revisiting woodfuel sustainability in the developing world. Environmental Research Letters, 12, 115002. https://doi.org/10.1088/1748-9326/aa83ed DOI: https://doi.org/10.1088/1748-9326/aa83ed
Bonilla, N., Cuesta, H., & Valois, H. (2011). Efectos de la extracción forestal sobre la estructura y composición de un bosque pluvial del Pacífico colombiano. Revista Biodiversidad Neotropical, 1(1), 48-54. http://dx.doi.org/10.18636/bioneotropical.v1i1.23 DOI: https://doi.org/10.18636/bioneotropical.v1i1.23
Bwalya, J. (2012). Estimation of net carbon sequestration potential of citrus under different management systems using the life cycle approach [Tesis de maestría, University of Zambia]. http://dspace.unza.zm/handle/123456789/2202
CDM Executive Board (2022a). AMS-II.G: Energy efficiency measures in thermal applications of non-renewable biomass (version 13.0). CDM, UNFCCC.
CDM Executive Board (2022b). Tool 30: Calculation of the fraction non-renewable biomass (version 04.0). CDM, UNFCCC.
CDM Meth Panel (2008). Annex 14 – Guidance on addressing uncertainty in the estimation of emissions reductions for CDM projects activities. CDM, UNFCCC.
Departamento Administrativo Nacional de Estadística & IDEAM (2021). Cuenta económica y ambiental del bosque (CAE-B). DANE, IDEAM.
Departamento Administrativo Nacional de Estadística (2019). Encuesta nacional de calidad de vida (ECV) 2018 – Anexos. https://www.dane.gov.co/index.php/estadisticas-por-tema/salud/calidad-de-vida-ecv/encuesta-nacional-de-calidad-de-vida-ecv-2018
Departmento Administrativo Nacional de Estadística (2020). Serie departamental de población por area para el periodo 2018-2050. https://www.dane.gov.co/index.php/estadisticas-por-tema/demografia-y-poblacion/proyecciones-de-poblacion
FAO & PNUMA (2020). El estado de los bosques del mundo 2020. Los bosques, la biodiversidad y las personas. FAO, PNUMA. https://doi.org/10.4060/ca8642es DOI: https://doi.org/10.4060/ca8642es
FAO (2021). Evaluación de los recursos forestal mundiales 2020 – Informe principal. https://doi.org/10.4060/ca8753es DOI: https://doi.org/10.4060/ca8753es
Farfán, F., & Rendón, J. (2014). Producción de madera por las variedades Castillo y Tabi en sistemas agroforestales (Avances Técnicos CENICAFE - Boletín No. 448). CENICAFE. https://biblioteca.cenicafe.org/handle/10778/563
Forest Carbon Partnership Facility (2020). FCPF methodological framework. Forest Carbon Partnership Facility (FCPF).
Gao, Y., Skutsch, M., Paneque, J., & Ghilardi, A. (2020). Remote sensing of forest degradation: a review. Environmental Research Letters, 15, 103001. https://doi.org/10.1088/1748-9326/abaad7 DOI: https://doi.org/10.1088/1748-9326/abaad7
Ghilardi, A., Bailis, R., Mas, J., Skutsch, M., Elvir, J., Quevedo, A., Masera, O., Dwivedi, P., Drigo, R., & Vega, E. (2016). Spatiotemporal modelling of fuelwood environmental impacts: Towards improved accounting for non-renewable biomass. Environmental Modelling & Software, 82, 241-254. https://doi.org/10.1016/j.envsoft.2016.04.023 DOI: https://doi.org/10.1016/j.envsoft.2016.04.023
Gill-Wiehl, A., Kammen, D., & Haya, B. (2023). Cooking the books: Pervasive over-crediting from cookstoves offsets methodology. Research Square. https://doi.org/10.21203/rs.3.rs-2606020/v1 DOI: https://doi.org/10.21203/rs.3.rs-2606020/v1
Hassanpoor Tichi, A., Khademieslam, H., & Rezanezhad Divkolae, M. (2020). Evaluation of physical, anatomical and biometric characteristics of Citrus sinensis trees in longitudinal and transverse axes of stem. BioResources, 15(3), 5696-5709. https://doi.org/10.15376/biores.15.3.5696-5709 DOI: https://doi.org/10.15376/biores.15.3.5696-5709
IPCC (2003). Definitions and methodological options to inventory emissions from direct human-induced degradation of forest and devegetation of others vegetation types. IGES, IPCC.
IPCC (2006). Volume 4: Agriculture, forestry and other land use. En H. Eggleston, L. Buendía, K. Miwa, T. Ngara & K. Tanabe (Eds.), 2006 IPCC guidelines for national greenhouse gas inventories (pp. 1-93). IGES, IPCC.
Lozano, L., & González, J. (2011). Bajo Calima: riqueza biológica y cultural afectada por la extracción de maderas tropicales. Revista Lebret, 3, 205-220. https://doi.org/10.15332/rl.v0i3.52 DOI: https://doi.org/10.15332/rl.v0i3.52
Meza, M., González, T., & Armenteras, D. (2018). Causas directas de degradación de bosques en Colombia: Sintesis de los patrones regionales. En D. Armenteras, M. Meza, J. Ramírez, E. Cabrera, G. Galindo & A. Yepes (Eds.), Causas de degradación forestal en Colombia: una primera aproximación (pp. 20-57). Universidad Nacional de Colombia, IDEAM, Programa ONU-REDD.
Ministerio de Agricultura y Desarrollo Rural (2019). Boletín estadístico forestal (boletín no. 1). https://vuf.minagricultura.gov.co/Documents/5.%20Estadisticas%20Sector%20Forestal/01_BOLETIN_FORESTAL_DICIEMBRE_2019.pdf
Ministerio de Agricultura y Desarrollo Rural (2021). Evaluaciones agropecuarias municipales 2006-2018. http://www.agronet.gov.co/estadistica/Paginas/home.aspx?cod=1
Mitchell, A., Rosenqvist, A., & Mora, B. (2017). Current remote sensing approaches to monitoring forest degradation in support of countries measurement, reporting and verification (MRV) systems for REDD+. Carbon Balance and Management, 12, 9. https://doi.org/10.1186/s13021-017-0078-9 DOI: https://doi.org/10.1186/s13021-017-0078-9
Murdiyarso, D., Skutsch, M., Guariguata, M., Kanninen, M., Luttrell, C., Verweij, P., & Martins, O. (2008). How do we measure and monitor forest degradation. En A. Angelsen (Ed.), Moving ahead with REDD: Issues, Options and Implications (pp. 99-106). CIFOR.
ep43zb
Parques Nacionales Naturales de Colombia - IDEAM (2021). Cifra de monitoreo de bosques IDEAM para las áreas protegidas del SPNN 1990-2020. Parques Nacionales Naturales de Colombia, IDEAM, Ministerio de Ambiente y Desarrollo Sostenible
Pearson, T., Brown, S., Murray, L., & Sidman, G. (2017). Greenhouse gas emissions from tropical forest degradation: an underestimated source. Carbon Balance and Management, 12, 3. https://doi.org/10.1186/s13021-017-0072-2 DOI: https://doi.org/10.1186/s13021-017-0072-2
Pulido, A., Chaparro, N., Granados, S., Ortiz, E., Rojas, A., Torres, C., & Turriago, J. (2018). Inventario nacional de emisiones de gases de efecto invernadero (2014). En IDEAM, PNUD, DNP, Ministerio de Ambiente y Desarrollo Sostenible & Cancillería de Colombia (Eds.), Segundo reporte bienal de actualización de Colombia a la Convención Marco de las Naciones Unidas para el Cambio Climático (pp. 68-177). IDEAM, PNUD, DNP, Ministerio de Ambiente y Desarrollo Sostenible, Cancillería de Colombia.
Rojas, Y., Loguercio, G., Nieto, V., & Bahamondez, C. (s.f.). Análisis de la degradación forestal en el marco de REDD+. INFOR, CIEFAP, CONIF, Instituto Forestal.
Romero, M., & Ponce, R. (2017). Assessing and monitoring forest degradation in a deciduos tropical forest in Mexico via remote sensing indicators. Forests, 8(9), 302. https://doi.org/10.3390/f8090302 DOI: https://doi.org/10.3390/f8090302
Samaniego, J., & Sánchez, E. (1999). Crecimiento y producción de cuatro cultivares de aguacate (Persea americana Mill.), en el sur de Sonora, México. Revista Chapingo - Serie Horticultura, 5, 61-66. https://doi.org/10.5154/r.rchsh.1999.05.037 DOI: https://doi.org/10.5154/r.rchsh.1999.05.037
Sharma, S., Rana, V. S., Prasad, H., Lakra, J., & Sharma, U. (2021). Appraisal of carbon capture, storage and utilization through fruits crops. Frontiers in Environmental Science, 9, 700768. https://doi.org/10.3389/fenvs.2021.700768 DOI: https://doi.org/10.3389/fenvs.2021.700768
The Gold Standard Foundation (2016). Fraction of non-renewable biomass (fNRB): Assessment for Bolivia, Colombia, Guatermala y Honduras. The Gold Standard Foundation. https://www.goldstandard.org/our-work/innovations-consultations/fraction-non-renewable-biomass-fnrb-assessment-bolivia-colombia
The Gold Standard Foundation (2022). Simplified methodology for clean and efficient cookstoves (version 3). The Gold Standard Foundation.
Thompson, I., Guariguata, M., Okabe, K., Bahamondez, C., Nasi, R., Heymell, V., & Sabogal, C. (2013). A operational framework for defining and monitoring forest degradation. Ecology and Society, 18(2), 1-20.. https://doi.org/10.5751/ES-05443-180220 DOI: https://doi.org/10.5751/ES-05443-180220
Unidad de Planeación Minero Energética (2019). Demanda energética. https://sig.upme.gov.co/SIPERS/Files
Wheeler, C., Mitchard, E., Nolasco, H., Iñíguez, G., Marquez, J., Carstairs, H., & Williams, M. (2021). A new field protocol for monitoring forest degradation. Frontiers in Forests and Global Change, 4, 655280. https://doi.org/10.3389/ffgc.2021.655280 DOI: https://doi.org/10.3389/ffgc.2021.655280
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Recibido: 24 de noviembre de 2022; Aceptado: 29 de noviembre de 2023
Resumen
El monitoreo de la degradación forestal y su efecto en el perfil de emisiones del sector AFOLU constituye un reto significativo para países cuyos bosques tienen un papel crucial en el cumplimiento de sus objetivos climáticos. Recientemente, se han propuesto diversos métodos para la medición de esta actividad REDD+, entre las que se considera el uso de indicadores de apoyo como complemento a los enfoques tradicionales. Se propone el uso de la fracción no renovable de la biomasa como un potencial indicador del estado de la degradación de los bosques derivado del aprovechamiento insostenible de la biomasa leñosa. Se estableció que los bosques de las regiones del Caribe y el Pacífico presentaban degradación forestal, con efecto sobre las emisiones por deforestación y degradación forestal de Colombia en 2018. Se recomienda incorporar este indicador como herramienta de apoyo a los sistemas MRV del sector forestal.
Palabras clave:
biomasa, emisiones, leña, REDD+, renovabilidad.Abstract
Forest degradation monitoring and its impact on the emissions profile of the AFOLU sector constitutes a significant challenge for countries whose forests play a crucial role in fulfilling climate goals. Recently, a diversity of methods for measuring this REDD+ activity have been proposed, which consider the use of supporting indicators as a complement to traditional approaches. The use of the fraction of non-renewable biomass is proposed as a potential indicator of the state of forest degradation derived from the unsustainable utilization of woody biomass. It was determined that the forests in the Caribbean and Pacific regions exhibit forest degradation, with an impact on the deforestation and forest degradation emissions of Colombia in 2018. We recommend incorporating this indicator as a supporting tool for MRV systems in the forestry sector.
Keywords:
biomass, emissions, fuelwood, REDD+, renewability.INTRODUCCIÓN
Con una superficie que abarca más de 4 000 millones de hectáreas, los bosques mundiales representan el único recurso de subsistencia para una tercera parte de la población mundial (FAO & PNUMA, 2020). La fuerte dependencia que existe de los bienes y servicios forestales ha generado una importante presión sobre estos ecosistemas, evidenciada principalmente en la pérdida de la cobertura forestal, que ha sido cuantificada en 420 millones de hectáreas desde 1990 (FAO, 2021). Sin embargo, los tensores a los cuales se ven sometidos los bosques son de distinta naturaleza y magnitud y no son necesariamente visibles y rastreables a través de los medios que usualmente se utilizan para el monitoreo de la cobertura forestal (Murdiyarso et al., 2008; FAO, 2021; Wheeler et al., 2021).
De particular interés para el ámbito de la mitigación del cambio climático es la degradación forestal, que, como concepto, puede tener varias acepciones dependiendo del campo especifico de análisis (Rojas et al., s.f.; Thompson et al., 2013). Sin embargo, para efectos del mecanismo de reducción de emisiones por deforestación y degradación forestal (REDD+), la degradación forestal se define como “la pérdida a largo plazo e inducida por el hombre (persistiendo por X años o más) de al menos Y % de los reservorios de carbono forestales (y otros beneficios del bosque) desde el tiempo T y que no califica como deforestación” (IPCC, 2003). Bajo esta definición, ciertas actividades generan un impacto paulatino sobre la estructura del bosque, propiciando la pérdida de algunos depósitos de carbono y, por tanto, generando emisiones de gases de efecto invernadero (GEI).
En Colombia, el consumo residencial de leña en el medio rural ha sido identificado como un agente causante de degradación forestal, principalmente en las regiones Andina y Caribe (Armenteras et al., 2018; Meza et al., 2018). Según el inventario de gases de efecto invernadero (INGEI) para 2014, las emisiones asociadas a la degradación forestal por consumo de leña fueron del orden de 14 Tg, lo que representaba el 17.6 % de las emisiones del sector forestal, el 10.6 % de las emisiones del sector de agricultura, silvicultura y otros usos de la tierra (AFOLU), y el 5.9 % de las emisiones totales nacionales (Pulido et al., 2018). Debe señalarse que, para efectos de una contabilidad de emisiones de GEI, la degradación forestal está dentro de la categoría de emisiones de bosque que permanecen como tales, en tanto que dicha actividad no constituye una conversión del uso del suelo (IPCC, 2006).
La magnitud de las emisiones de GEI procedentes de la degradación forestal cobra especial importancia para las diferentes iniciativas que le apuestan al financiamiento de REDD+, dada la necesidad de garantizar sistemas de monitoreo, reporte y verificación (MRV) cada vez más transparentes y exhaustivos con la contabilidad de GEI. Tales iniciativas exigen el reporte de las emisiones por degradación forestal en función de su significancia frente a las emisiones generadas por el sector forestal (Pearson et al., 2017; Architecture for REDD+ Transactions, 2021; Forest Carbon Partnership Facility, 2021).
Sin embargo, a diferencia de la deforestación, establecer datos de actividad para la degradación forestal ha resultado ser un asunto lleno de complejidades, dado que las técnicas actuales de monitoreo resultan insuficientes o inadecuadas para rastrear la pérdida de los depósitos de carbono (Romero & Ponce, 2017; Ahrends et al., 2021; Wheeler et al., 2021). Lo anterior aplica particularmente a la recolección de leña, pues, a diferencia de los otros motores usualmente asociados a la degradación forestal (i.e., extracción selectiva de madera o incendios forestales), los rastros de tal actividad son menos evidentes en la estructura del bosque y solo pueden ser inferidos a partir de variables exógenas que suponen su extracción de dichos ecosistemas.
No obstante, la leña no tiene a los ecosistemas forestales como único nicho de recolección, en tanto que otros usos del suelo también pueden proveer biomasa leñosa. Lo anterior desdibuja el impacto real que esta práctica tiene sobre la degradación forestal. Luego, establecer su verdadera magnitud constituye un paso esencial para comprender la incidencia de la recolección de leña como un factor que contribuye a las emisiones de GEI del sector forestal. Recientemente, el mercado de carbono ha catalizado el desarrollo de metodologías que apuntan a medir el impacto del consumo de leña en las emisiones de GEI. Estas metodologías han sido generadas desde una perspectiva energética y pocas veces han sido pensadas para determinar sus efectos sobre los ecosistemas forestales y en articularse con el sector AFOLU (CDM Executive Board, 2022a; The Gold Standard Foundation, 2022). Esta falta de articulación entre enfoques metodológicos podría redundar en contradicciones frente a la forma en que se contabilizan las emisiones, derivando incluso en problemas de doble contabilidad.
Una variable de interés para el sector AFOLU es la fracción no renovable de la biomasa (fNRB), la cual puede definirse como la proporción de la biomasa leñosa extraída dentro de un área geográfica especifica en una magnitud que excede la capacidad regenerativa de los ecosistemas forestales y, por extensión, de los ecosistemas vegetales presentes en dicha área, generando una pérdida paulatina de los stocks de biomasa aérea. Su uso ha tenido efectos prácticos para contabilizar las reducciones de emisiones de proyectos que disminuyen el consumo de leña (i.e, estufas eficientes), pero de poca aplicación en proyectos de mitigación relacionados con la gestión de los bosques (i.e, REDD+).
Este artículo de investigación propone el uso de esta variable como un indicador de la degradación forestal a partir de la estimación efectuada para cada región político-administrativa de Colombia, de tal manera que pueda ser armonizada y considerada en futuros niveles de referencia de emisiones forestales (NREF) que se desarrollen para el país.
MATERIALES Y MÉTODOS
Para calcular la fNRB de cada región de Colombia, se utilizó la herramienta metodológica no. 30 definida por la Convención Marco de las Naciones Unidas sobre Cambio Climático (CMNUCC), en la cual se establece el procedimiento para calcular dicha variable (CDM Executive Board, 2022b). La fracción no renovable de la biomasa se estima a partir de la Ecuación (1):
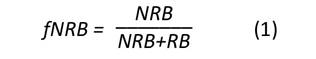
Esta ecuación expresa fNRB como el cociente resultante de dividir la cantidad de biomasa no renovable (NRB) para un área geográfica especifica entre la cantidad de biomasa renovable (RB) sumada a la no renovable que ha sido definida para esa misma área. Estos parámetros pueden ser expresados en unidades de masa o de volumen en función de un periodo de tiempo (usualmente Mg.año-1 o m3.año-1). A su vez, el cálculo de NRB corresponde a la diferencia entre el consumo total anual de biomasa leñosa (H) (madera y leña principalmente) y la cantidad de RB, tal como se indica en la Ecuación (2):
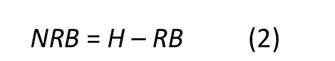
Por su parte, la RB se obtiene como la suma de las existencias de biomasa en crecimiento de las diferentes coberturas vegetales que se encuentren dentro del área geográfica de análisis. Estas coberturas no se limitan a ecosistemas forestales, sino que deben incluir todos los usos del suelo con potencial para generar biomasa leñosa (i.e., áreas de cultivo, sistemas agroforestales, plantaciones forestales, etc.). Este parámetro se obtiene a través de la siguiente expresión Ecuación (3):
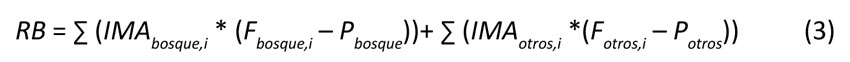
Cada variable de esta expresión puede ser consultada en la herramienta metodológica mencionada anteriormente. Para hallar cada variable, fue necesario recurrir a información secundaria, tomando 2018 como año de referencia. Se empleó la cifra de incremento medio anual (IMA) para bosques utilizada en la Cuenta Ambiental y Económica del Bosque (CAE-B), que establece una productividad en términos de volumen de 0.41 m3.ha-1.año-1 (Departamento Administrativo Nacional de Estadística & IDEAM, 2021). Asimismo, se utilizó un incremento medio anual para plantaciones forestales de 12 m3.ha-1.año-1, como se sugiere en dicha publicación.
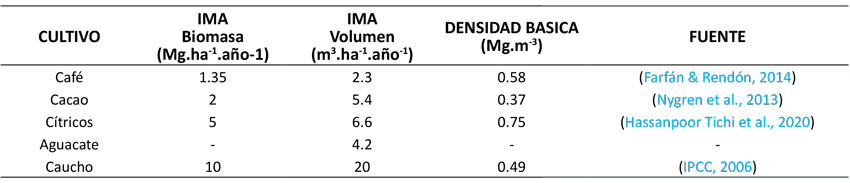
Para asegurar un cálculo lo más robusto posible de fNRB, también se incluyó la biomasa leñosa producida en áreas que no están dentro de la definición de bosque o de plantación forestal. Para tal efecto, se escogieron los cinco cultivos leñosos permanentes con mayor superficie a nivel nacional en 2018, tal como se presenta en la Tabla 1 (Ministerio de Agricultura y Desarrollo Rural, 2021).
Fuente: Elaboración propia con base en datos del Ministerio de Agricultura y Desarrollo Rural (2018)
Tabla 1: Cultivos leñosos permanentes con mayor superficie nacional de siembra en 2018
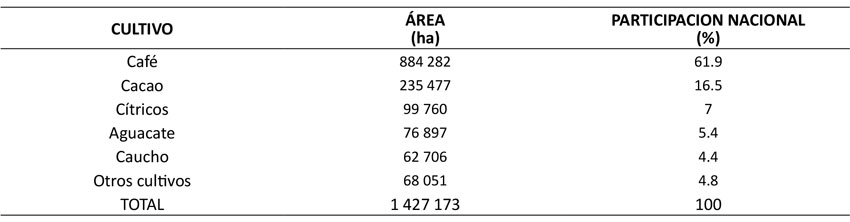
Con excepción del aguacate, los incrementos medios anuales (IMA) para los cuatro cultivos leñosos restantes fueron obtenidos a partir de fuentes secundarias. Para café y cacao, se reportaron incrementos en biomasa de 1.35 y 2 Mg.ha-1.año-1 respectivamente (Aristizábal, 2015); para los cítricos, se consideró un incremento de 5 Mg.ha-1.año-1 (Bwalya, 2012; Sharma et al., 2021); mientras que, para el caucho, se asumió un valor mínimo por defecto de 10 Mg.ha-1.año-1, según lo sugerido por el Panel Intergubernamental sobre Cambio Climático (IPCC, 2006). En el caso del aguacate, el IMA en biomasa fue proyectado a partir de una tabla volumétrica convencional construida con cifras de crecimiento diamétrico y de altura para esta especie. Esto, de acuerdo con datos reportados por Samaniego y Sánchez (1999) para plantaciones de este cultivo en México.
Para garantizar la homogeneidad de los datos utilizados, los valores de biomasa fueron transformados a unidades de volumen mediante las densidades básicas reportadas para los cinco cultivos referidos. La Tabla 2 muestra la transformación de biomasa a volumen aplicando la densidad básica correspondiente.
Tabla 2: Densidades básicas de los cultivos leñosas utilizadas en la conversión de biomasa a volumen
Para cada región político-administrativa, se calculó la superficie de bosque en áreas protegidas para el año 2018, representada principalmente por las áreas del sistema nacional de Parques Nacionales que presentan este tipo de cobertura según el Sistema de Monitoreo de Bosques y Carbono (SMByC) del Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia (IDEAM) (Parques Nacionales Naturales de Colombia - IDEAM, 2021). Este parámetro es necesario, pues la RB se calcula solo a partir de áreas de bosque que no tengan ningún tipo de restricción respecto a su accesibilidad física y legal. El primer caso implica eliminar del análisis las áreas de bosque que, por distancia o condiciones topográficas, son de difícil acceso y, por lo tanto, se presume no serán consideradas como una fuente de extracción de productos maderables. Por otro lado, la accesibilidad legal supone restricciones en el aprovechamiento de recursos forestales que procedan de áreas geográficas que, jurídicamente hablando, deben cumplir un papel en la protección y conservación de los bosques y, por consiguiente, no admiten ninguna actividad que implique la extracción de productos maderables.
Debido a la escala del análisis, solo se consideró la restricción por accesibilidad legal, dada la limitada disponibilidad de información con la que cuenta el país en términos de accesibilidad física. Sustraer áreas de bosque por accesibilidad física requiere análisis a escalas de mayor detalle que están fuera del alcance de la presente investigación, pues esto requiere el uso de información espacialmente explicita que relacione dinámicas poblacionales, infraestructura vial y tiempo de recorrido hasta las áreas boscosas.
El área de bosque accesible para cada región político-administrativa se calculó como la diferencia entre la superficie total de bosque para dicha región y la superficie de bosque presente en las áreas protegidas que hacen parte de la misma.
El consumo total de biomasa leñosa (H) solo consideró el consumo residencial rural de leña y la extracción total de madera para el año 2018. Para el primer caso, se utilizaron los consumos per capita obtenidos de los Planes de Energización Rural Sostenible (PERS) de los departamentos de Cesar, Chocó, Cundinamarca, La Guajira, Nariño, Norte de Santander y Tolima. De allí se infirió un valor promedio nacional que fue aplicado a los demás departamentos del país. Se utilizaron los datos poblacionales rurales del año 2018 arrojados por el último censo demográfico (Departamento Administrativo Nacional de Estadística, 2020), los cuales fueron ajustados con base en el porcentaje de la población rural que solo utiliza leña como combustible de cocción (Departamento Administrativo Nacional de Estadística, 2019; Unidad de Planeación Minero Energética, 2019). Con base en lo anterior, se determinó el consumo total de leña por departamento y, posteriormente, se consolidó la información para cada una de las regiones político-administrativas.
Por otro lado, el consumo de madera para usos no energéticos se estimó a partir de las cifras de volumen de madera movilizada de plantaciones comerciales en 2018 y reportadas por el Ministerio de Agricultura y Desarrollo Rural (2019), considerando su participación dentro del volumen total nacional, que correspondería al 60.9 %, mientras que el 39.1 % restante procedería del bosque natural (Departamento Administrativo Nacional de Estadística & IDEAM, 2021). Debe indicarse que, para efectos de este análisis, no fue posible incluir la leña usada como combustible en aplicaciones no residenciales (i.e., los sectores industrial, comercial e institucional), pues no existen estadísticas confiables sobre dichos consumos desagregados a nivel departamental.
RESULTADOS
Biomasa renovable (RB)
Se obtuvieron datos de RB tanto para el componente forestal como para el agrícola. El componente forestal fue dividido en biomasa renovable procedente de bosques naturales (RBnf) y biomasa renovable de plantaciones forestales (RBpf). Dado que en las regiones de la Amazonía, la Orinoquía y el Pacífico se concentra la mayor superficie de bosque del país, en estas regiones se obtuvieron las cifras más altas de biomasa renovable (RBnf), del orden de 10.92, 3.07 y 3 Mm3.año-1. La región Caribe presentó la menor cantidad de RBnf, con solo 0.48 Mm3.año-1 (Tabla 3).
Nota: RBnf: biomasa renovable de bosque natural; RBnf+pf+ag: la biomasa renovable conjunta de bosque natural, plantaciones forestales y cultivos agrícolas; H: tasa de extracción (equivalente a consumo de recursos leñosos); NRBnf: la biomasa no renovable que solo contempla el bosque natural como fuente del recurso; NRBnf+pf+ag: la biomasa no renovable que contempla tanto bosque natural como las plantaciones forestales y los cultivos agrícolas; fNRBnf: la fracción no renovable de la biomasa calculada con NRBnf,; fNRBnf+pf+ag: la fracción no renovable de la biomasa calculada con NRBnf+pf+ag.
Tabla 3: Cálculo comparativo de fNRB por regiones (bosque natural vs. todos los componentes juntos)
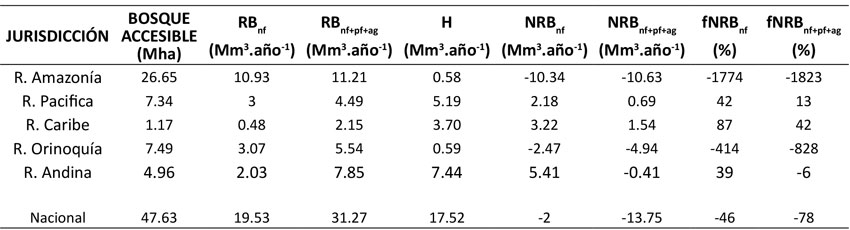
En lo que respecta a la biomasa renovable de plantaciones forestales (RBpf), los mayores valores se observaron en las regiones Andina, de la Orinoquía y del Caribe, con 2.45, 1.71 y 1.26 Mm3.año-1 respectivamente. Esto se debe a que en estas regiones se encontraba la mayor cantidad de plantaciones establecidas en 2018, con 39.7, 12 y 20.4 % de la superficie plantada respectivamente (Tabla 3).
Al igual que la RBpf, la biomasa renovable de cultivos agrícolas (RBag) se generó de manera proporcional a la superficie dedicada a este uso del suelo. Por lo tanto, la región Andina fue, de lejos, la que produjo mayor cantidad de biomasa renovable, con 3.36 Mm3, seguida de la Orinoquía con 0.75 Mm3 y la región del Pacífico con 0.74 Mm3 (Tabla 3). Lo anterior resulta de las extensas plantaciones de café, cacao y aguacate que existen en los departamentos de esta región y que, en conjunto con los cítricos y el caucho, representaron el 67 % del área sembrada con respecto al área total sembrada de estos cinco cultivos a nivel nacional en 2018.
Al totalizar los componentes de la biomasa renovable (i.e., RBnf , RBpf y RBag), se obtuvo que la región Amazónica generó 11.2 Mm3.año-1, mientras que la región Andina produjo 7.85 Mm3.año-1 y la Orinoquía 5.54 Mm3.año-1. Las regiones del Pacífico y el Caribe solo generaron 4.49 y 2.16 Mm3.año-1 respectivamente.
Extracción de biomasa leñosa (H)
El consumo de biomasa leñosa (madera y leña) reportó valores altos en las regiones Andina y del Pacífico, con 7.90 y 5.56 Mm3 respectivamente. Por su parte, la Orinoquía y la región Amazónica presentaron las cifras más bajas de extracción total de biomasa leñosa (Tabla 3).
Biomasa no renovable (NRB)
Se calcularon los valores de NRB considerando dos escenarios: 1) analizando solo la biomasa renovable procedente de bosque natural y 2) considerando todos los tres componentes juntos (i.e., biomasa renovable de bosque natural, plantaciones forestales y cultivos agrícolas). Los valores negativos indican que la tasa de incremento del recurso (los crecimientos de la biomasa) es superior a la tasa de extracción del mismo, mientras que los valores positivos denotan la situación opuesta para el área de análisis (Tabla 3).
Estos dos escenarios revelan diferencias notables. En el primer caso, donde la biomasa leñosa solo se extrae de bosques naturales, los valores aumentan dramáticamente si se comparan con el escenario en que los tres componentes se constituyen como la fuente del recurso. Se evidencia que, en las regiones Amazónica y de la Orinoquía, no existe una condición deficitaria del recurso leñoso, pues en ambos escenarios los resultados son negativos; mientras que los valores positivos de las regiones del Pacífico y el Caribe para ambos escenarios indican insostenibilidad en la extracción de recursos leñosos, pues los ecosistemas forestales no recuperan la biomasa a la misma velocidad en que esta es extraída como productos forestales maderables o como leña para uso energético. La región Andina es un caso excepcional dado que la tasa de incremento de la biomasa se encuentra más o menos balanceada con la tasa de extracción sobre una base anual. No obstante, se presenta no renovabilidad de la biomasa bajo el escenario en el que los recursos maderables se extraen exclusivamente del bosque. Dicha condición desaparece cuando el cálculo integra la biomasa procedente de las plantaciones forestales y los cultivos agrícolas, dado que, solo al sumar los aportes de los incrementos de biomasa de estas coberturas, la biomasa general de la región supera la tasa de extracción (equivalente al consumo de biomasa leñosa).
Fracción no renovable de la biomasa (fNRB)
Como puede observarse en la Figura 1, la región del Caribe es la que enfrenta una mayor presión sobre sus bosques naturales, dado que, al considerar las tres coberturas vegetales, más del 42 % del recurso forestal extraído en forma de leña o madera no es compensado por la biomasa en crecimiento, situación que se agudiza aún más si se tiene en cuenta que los bosques naturales son la única fuente de extracción. En dicha circunstancia, esta cifra prácticamente se duplica.
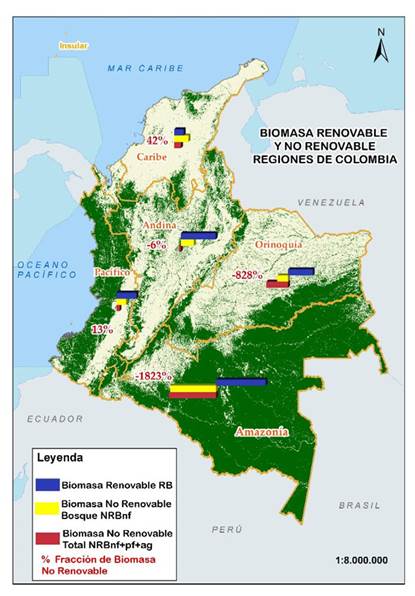
Figura 1: Fracción no renovable de la biomasa (fNRB) por regiones político-administrativas para el año 2018
Una situación muy similar puede observarse en la región del Pacífico, donde el primer escenario indica una fNRB del 42 %, que se reduce al 13 % cuando se incluyen los otros dos usos del suelo (plantaciones forestales y cultivos agrícolas). La región Andina no presenta una condición crítica para el año de referencia (2018), en virtud de que la extracción total de biomasa leñosa es más o menos equiparable con la biomasa renovable total en el escenario que incluye los tres usos del suelo, generando una fNRB de básicamente 0 %, lo que indica que las pérdidas apenas logran ser compensadas con las ganancias de biomasa. Si solo se consideran los incrementos de los bosques naturales de la región Andina, existiría una situación deficitaria, pues la extracción de recursos leñosos prácticamente cuadruplicaría el potencial regenerativo de estos ecosistemas forestales, lo que supondría una mayor presión sobre los mismos.
DISCUSIÓN
La degradación forestal constituye una de las actividades REDD+, cuyo monitoreo presenta los mayores retos. Hasta la fecha, solo se han identificado sus principales causas, pero con avances muy limitados en la identificación de su ocurrencia dentro de determinada área geográfica. Lo anterior se debe a la naturaleza particular de este fenómeno, que tiende a variar en marcos tanto espaciales como temporales (Mitchell et al., 2017; Thompson et al., 2013). En años recientes, han prevalecido los enfoques de monitoreo basados en la percepción remota de las coberturas forestales (Mitchell et al., 2017; Romero & Ponce, 2017; Gao et al., 2020; Ahrends et al., 2021). Sin embargo, estos enfoques presentan limitaciones cuando se trata de hacer seguimiento a los componentes estructurales que se encuentran por debajo del dosel del bosque (Gao et al, 2020; Ahrends et al, 2021; Wheeler et al., 2021). Algunos autores sugieren el uso de indicadores como un complemento (y en algunos casos como alternativa) al enfoque de monitoreo basado en sistemas de teledetección, entre ellos el almacenamiento de carbono (Thompson et al., 2013). En este artículo se propone una variable de uso común para cuantificar los resultados de mitigación obtenidos por las iniciativas que reducen el consumo de biomasa leñosa mediante la implementación de tecnologías de combustión que desplazan el uso de combustibles dendroenergéticos y que, por tanto, reducen las existencias de biomasa dentro de una determinada área geográfica.
Los valores de fNRB regionalizados demuestran una extracción insostenible de la biomasa en las regiones del Caribe y el Pacífico, dado que el 42 y 13 % (respectivamente) de la biomasa extraída de estas regiones no está siendo ‘compensada’ por las existencias en crecimiento, i.e., la tasa de extracción es superior a los incrementos que allí se observan. Esto puede ser explicado en buena medida por el tamaño de las poblaciones rurales de estas regiones, las cuales demandan importantes cantidades de leña para cocción doméstica, generando una importante presión sobre los ecosistemas forestales.
Para la región del Caribe, la situación se torna crítica, pues la superficie de bosque accesible solo representa una sexta parte en comparación con la región del Pacífico (1.17 y 7.34 Mha respectivamente). Además, a pesar de que la región cuenta con una mayor superficie de plantaciones forestales, los crecimientos de las existencias de biomasa resultan insuficientes para equiparar el nivel de extracción, lo cual se agudiza teniendo en cuenta que existe un área relativamente pequeña dedicada a cultivos agrícolas leñosos (predominantemente café y cacao).
Si bien cuenta con la tercera extensión de bosque más grande del país (después de la Amazonía y la Orinoquía), el 40 % del aprovechamiento de madera tanto de bosque natural como de plantaciones forestales ocurre en la región del Pacífico, lo cual tiene un efecto notable en la configuración del parámetro NRB regional. La extracción de madera comercial no solo representa cerca del 30 % de la extracción total de biomasa leñosa, sino que incluso constituye casi el 50 % del volumen de biomasa leñosa que anualmente se regenera en la región. En otras palabras, se extrae 1 m3 de biomasa leñosa por cada 2.5 m3 que se regeneran en las coberturas forestales del Pacífico colombiano. En contraste, en el caso de la región Caribe, esta misma relación fue de 1:9. Lo anterior indica que el aprovechamiento forestal (tanto legal como ilegal) sí es un motor de degradación forestal relevante en la región Pacífica, situación previamente advertida por distintos estudios en aspectos tanto cuantitativos como estructurales del bosque (Bonilla et al., 2011; Lozano & González, 2011; Meza et al., 2018).
Por otro lado, la región Andina se enfrenta a una situación peculiar: su fNRB es básicamente del 0 %, i.e., las existencias en crecimiento se encuentran más o menos en equilibrio con la tasa de extracción (7.85 y 7.44 Mm3 respectivamente). Lo anterior se debe al volumen de RB que genera el componente agrícola, que representa casi el 43 % de la RB total para dicha región, en comparación con el 31 % que aportan las plantaciones forestales y el 26 % que generan los bosques naturales allí presentes. De esta forma, se hace evidente el papel crucial de los cultivos agrícolas leñosos en la provisión de leña como recurso energético y de otros productos maderables. Estos cultivos contribuyen a reducir la presión sobre los ecosistemas forestales andinos, que tradicionalmente han servido de fuente de extracción de este tipo productos, lo cual ha conducido a su paulatina desaparición. Se calcula que, al sustraer del análisis la biomasa leñosa agrícola, la región Andina incrementaría notablemente su fNRB (39 %). Esto indicaría que dos de cada cinco unidades de biomasa leñosa extraídas no podrían ser compensadas por la vegetación en crecimiento en la región.
Un dato contrastante hallado en el presente análisis con respecto al valor calculado en la CAE-B tiene que ver con la variable denominada extracción sostenible, que se define como “la cantidad de madera que puede ser cosechada sin poner en riesgo la resiliencia y capacidad productiva de los bosques y otras coberturas boscosas” y que fue estimada en 6.66 Mm3 para todo el país (Departamento Administrativo Nacional de Estadística & IDEAM, 2021). Este parámetro sería equiparable a la RBnf, y en este estudio se calculó en 19.53 Mm3 para todo el país. Las discrepancias entre ambas cifras pueden obedecer principalmente a la superficie total de bosque considerada para la estimación, pues ambas variables emplearon el mismo IMA. En esta investigación se aplicó el incremento medio anual sobre la superficie de bosque accesible (47.6 Mha), pero se desconoce la cifra utilizada por la CAE-B para determinar dicha variable.
Los resultados derivados de esta investigación controvierten los análisis de fNRB realizados por estudios previos, en los cuales se estimaron valores para Colombia que superaban el 70 % a nivel regional y nacional (Aristizábal, 2015; The Gold Standard Foundation, 2016). Esto indicaba un nivel crítico de regeneración de los ecosistemas forestales frente a la tasa de extracción de productos forestales para las áreas analizadas.
En contraste, la presente evaluación arroja un valor negativo de fNRB (-78 %) si se analiza el país como una sola unidad geográfica, lo cual demuestra que, considerando las tres coberturas analizadas, la tasa de extracción nacional se encuentra muy por debajo del incremento anual de la biomasa leñosa. Sin embargo, este resultado debe tomarse con cautela, puesto que, si el análisis se realiza a escalas geográficas más pequeñas, los valores de fNRB pueden variar sustancialmente, como lo demuestran cada uno de los resultados regionales. Lo anterior indica que la configuración geográfica del área de análisis juega un papel crucial en la determinación de la fNRB, pues los diferentes parámetros pueden variar notablemente.
Cabe señalar que las fNRB calculadas para las cinco regiones ratifican los hallazgos previos de otros autores que utilizaron modelos basados en análisis espaciales, estableciendo variaciones regionales de fNRB que fluctuaban entre el 0 y el 40 % para Colombia (Bailis et al., 2015).
En términos de la contabilidad de GEI, los resultados obtenidos sugieren que solo el 13 % de las emisiones generadas por la extracción de biomasa leñosa de los bosques de la región del Pacífico pueden ser consideradas como emisiones netas a la atmósfera, en tanto que en la región Caribe solo podría contabilizarse el 42 % de las emisiones generadas por el mismo concepto. Esto implica una emisión total por degradación forestal de 3.4 TgCO2eq para el año 2018, lo que solo representa una cuarta parte de las emisiones que fueron reportadas originalmente por el INGEI 2014.
Debe señalarse que el enfoque metodológico utilizado, si bien es poco demandante en cuanto a la cantidad de parámetros requeridos para hallar la fNRB, arroja una estimación razonablemente coherente en términos de lo que se espera que sea el comportamiento de la condición de no renovabilidad de la biomasa para un área geográfica en particular. En años recientes han surgido criticas frente a la utilización de este procedimiento metodológico. Esto, debido a su ‘estaticidad’ y su potencial sobreestimación de esta condición para los ecosistemas forestales presentes en una determinada área geográfica (Ghilardi et al, 2016; Bailis et al., 2017; Gill-Wiehl et al., 2023). Sin embargo, los datos hallados en este estudio indican que, si la aplicación de la metodología planteada en la herramienta no. 30 incorpora la mayor cantidad de parámetros requeridos, esta puede resultar útil para determinar la fNRB dentro de rangos que no sobreestimen la condición de no renovabilidad. Se observa, por ejemplo, que las cifras de fNRB tienden a ser más conservadoras y cercanas a la realidad cuando se incluyen otros usos del suelo que proveen biomasa leñosa adicionales a las tierras forestales (i.e., tierras agrícolas y/o pastizales arbolados).
No obstante, la herramienta metodológica tiene limitaciones en su aplicabilidad, que deberían ser subsanadas en aras de incrementar su nivel de precisión en el resultado final. Por un lado, el resultado es altamente sensible a la configuración geográfica del área de análisis. Los resultados de fNRB tienden a ser más coherentes y relativamente precisos cuando la evaluación se efectúa a escalas finas en comparación con las escalas gruesas, lo que es una consecuencia de la variación en los parámetros utilizados (i.e., extensión y tipo de bosque, incrementos medios anuales, datos poblacionales, usos del suelo considerados, etc.). Por otra parte, debería considerarse el nivel de incertidumbre asociado a variables como el IMA aplicado a las coberturas vegetales evaluadas, el consumo promedio de leña por hogar para las diferentes escalas de análisis, el consumo de leña para uso energético no residencial y el volumen de madera extraído de las coberturas forestales. Si bien la herramienta metodológica per se no provee indicaciones para el manejo de la incertidumbre, es claro que, al tratarse de una herramienta de apoyo a metodologías MDL aprobadas por la CMNUCC, el tratamiento de los parámetros considerados debería adherirse a las directrices establecidas para la adecuada gestión de este atributo (CDM Meth Panel, 2008).
A pesar de las limitaciones anteriormente mencionadas, una de sus ventajas tiene que ver con la inclusión de los consumos de biomasa leñosa para usos no energéticos, lo que le confiere versatilidad para incorporar el aprovechamiento selectivo de madera como motor de degradación forestal dentro de un análisis que no solo se circunscribe a la recolección de leña, sino que integra otros elementos cuya espacialización enfrenta dificultades. No obstante, la inclusión de este driver dependerá, en buena medida, de información estadística robusta sobre volúmenes de madera que puedan ser rastreables hasta el área de extracción, pues resulta crítico asignar un valor correspondiente a este parámetro a cada área definida para el análisis.
CONCLUSIONES
Este artículo propone el uso de la fracción no renovable de la biomasa (fNRB) como un indicador de degradación forestal aplicable a escalas nacionales o subnacionales. Este indicador señala que, en 2018, los bosques existentes en las regiones del Pacífico y el Caribe presentaban importantes niveles de degradación, pues la tasa de extracción de biomasa excedió la tasa de regeneración en un 13 y un 42% respectivamente. La aplicación de esta metodología está supeditada a las realidades y circunstancias propias de cada jurisdicción en cuanto a disponibilidad de información y necesidades de reporte en el marco de sus propios sistemas MRV.
REFERENCIA




Licencia
Derechos de autor 2024 Colombia forestal

Esta obra está bajo una licencia internacional Creative Commons Atribución-CompartirIgual 4.0.
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






