
DOI:
https://doi.org/10.14483/2256201X.20940Publicado:
01-01-2024Número:
Vol. 27 Núm. 1 (2024): Enero-junioSección:
Artículos de investigación científica y tecnológicaConsumo de polen por sírfidos (Diptera: Syrphidae) en una cuenca urbana altoandina con influencia antrópica
Pollen Consumption by Syrphids (Diptera: Syrphidae) in a High Andean Urban Watershed with Anthropic Influence
Palabras clave:
Anidamiento, coberturas antropizadas, conectividad, redes bipartitas, resiliencia (es).Palabras clave:
Nestedness, anthropized coverage, connectivity, bipartite networks, resilience (en).Descargas
Referencias
Ángel, S. L., Bogotá-A, R. G. & Montoya, A. L. (2021). Comunidades de Sírfidos asociadas a coberturas influenciadas por actividades antrópicas en Bogotá D.C. Caldasia, 43(1), 161-171. https://dx.doi.org/10.15446/caldasia.v43n1.82464 DOI: https://doi.org/10.15446/caldasia.v43n1.82464
Bogotá, R. G., Lamprea, S. J. & Rangel-Ch. J. O. (1996). Atlas palinológico de la clase Magnoliopsida (excepto Asteridae) en el Páramo de Monserrate. En Fundación Ecosistemas Andinos ECOAN (Eds.), El páramo, ecosistema a proteger (pp. 131-191). Editorial CODICE.
Bogotá-A, R. G. (2002). El polen de la Subclase Asteridae en el Páramo de Monserrate. Centro de Investigaciones y Desarrollo Científico, Universidad Distrital Francisco José de Caldas.
Branquart, E., & Hemptinne, J. L. (2000). Selectivity in the exploitation of floral resources by hoverflies (Diptera: Syrphinae). Ecography, 23(6), 732-742. http://dx.doi.org/10.1111/j.1600-0587.2000.tb00316.x DOI: https://doi.org/10.1111/j.1600-0587.2000.tb00316.x
Carabalí, A., Pinchao, S. Y., Lamprea, I. M., Peña, J. F. & Carabalí, D. J. (2017). Insectos polinizadores del aguacate (Persea americana Mill.) cv. Hass en Colombia. CORPOICA. DOI: https://doi.org/10.21930/978-958-740-235-3
Dalsgaard, B., Schleuning, M., Maruyama, P. K., Dehling, D. M., Sonne, J., Vizentin‐Bugoni, J., Zanata, T. B., Fjeldså, J., Böhning-Gaese, K., & Rahbek, C. (2017). Opposed latitudinal patterns of network‐derived and dietary specialization in avian plant–frugivore interaction systems. Ecography, 40(12), 1395-1401. https://dpi.org/10.1111/ecog.02604 DOI: https://doi.org/10.1111/ecog.02604
Dormann, C. F., Fruend, J., Gruber, B., Dormann, M. C. F., LazyData, TRUE, & ByteCompile. (2019). ‘bipartite’ package. R. package version 2.18. https://cran.r-project.org/web/packages/bipartite/index.html
Erdtman, G. (1986). Pollen morphology and plant taxonomy: Angiosperms. Hafner. DOI: https://doi.org/10.1163/9789004612150
Gervais, A., Chagnon, M., & Fournier, V. (2018). Diversity and pollen loads of flower flies (Diptera: Syrphidae) in cranberry crops. Annals of the Entomological Society of America, 111(6), 326-334. https:/doi.org/10.1093/aesa/say027 DOI: https://doi.org/10.1093/aesa/say027
Holloway, B. A. (1976). Pollen-feeding in hover-flies (Diptera: Syrphidae). New Zealand Journal of Zoology, 3(4), 339-350. https://doi.org/10.1080/03014223.1976.9517924 DOI: https://doi.org/10.1080/03014223.1976.9517924
Hu, S., Dilcher, D. L., Jarzen, D. M., & Taylor, D. W. (2008). Early steps of angiosperm-pollinator coevolution. Proceedings of the National Academy of Sciences, 105(1), 240-245. https://doi.org/10.1073/pnas.0707989105 DOI: https://doi.org/10.1073/pnas.0707989105
Hughes, C. E. (2016). The tropical Andean plant diversity powerhouse. New Phytologist, 210(4), 1152–1154. https://doi.org/10.1111/nph.13958 DOI: https://doi.org/10.1111/nph.13958
IDEAM. (2010). Leyenda nacional de coberturas de la tierra. Metodología CORINE Land Cover adaptada para Colombia, Escala 1:100.000. Instituto de Hidrología, Meteorología y Estudios Ambientales.
Jordano, P., Vázques, D., & Bascompete, J. (2009). Redes complejas de interacciones planta-animal. En R. Medel, M. A. Aizen, & R. Zamora (Eds.), Ecología y evolución de las interacciones planta animal: conceptos y aplicaciones (pp. 17-41). Editorial Universitaria.
Klecka, J., Hadrava, J., Biella, P., & Akter, A. (2018). Flower visitation by hoverflies (Diptera: Syrphidae) in a temperate plant-pollinator network. PeerJ Preprints, 6, e26516v2. https://doi.org/10.7287/peerj.preprints.26516v2 DOI: https://doi.org/10.7717/peerj.6025
Lever, J. J., van Nes, E. H., Scheffer, M., & Bascompte, J. (2014). The sudden collapse of pollinator communities. Ecology Letters, 17, 350-359. https://doi.org/10.1111/ele.12236. DOI: https://doi.org/10.1111/ele.12236
Lucas, A., Bodger, O., Brosi, B. J., Ford, C. R., Forman, D. W., Greig, C., Hegarty M., Neyland P. J., & de Vere, N. (2018). Generalisation and specialisation in hoverfly (Syrphidae) grassland pollen transport networks revealed by DNA metabarcoding. Journal of Animal Ecology, 87(4), 1008-1021. https://doi.org/10.1111/1365-2656.12828 DOI: https://doi.org/10.1111/1365-2656.12828
Memmott, J., Waser, N. M., & Price, M. V. (2004). Tolerance of pollination networks to species extinctions. Proceedings of the Royal Society B, 271, 2605-2611. DOI: https://doi.org/10.1098/rspb.2004.2909
Menke, S., Böhning-Gaese, K., & Schleuning, M. (2012). Plant-frugivore networks are less specialized and more robust at forest–farmland edges than in the interior of a tropical forest. Oikos, 121(10), 1553-1566. https://doi.org/10.1111/j.1600-0706.2011.20210.x DOI: https://doi.org/10.1111/j.1600-0706.2011.20210.x
Orford, K. A., Vaughan, I. P. & Memmott, J. (2015). The forgotten flies: The importance of non-syrphid Diptera as pollinators. Proceedings of the Royal Society B: Biological Sciences, 282(1805), 20142934. https://doi:10.1098/rspb.2014.2934. DOI: https://doi.org/10.1098/rspb.2014.2934
Petanidou, T., Kallimanis, A. S., Tzanopoulos, J., Sgardelis, S. P., & Pantis, J. D. (2008). Long‐term observation of a pollination network: Fluctuation in species and interactions, relative invariance of network structure and implications for estimates of specialization. Ecology Letters, 11, 564-575. https://doi.org/10.1111/j.1461-0248.2008.01170.x DOI: https://doi.org/10.1111/j.1461-0248.2008.01170.x
R Development Core Team (2015). R: A language and environment for statistical computing. R Foundation for Statistical Computing.
Radenkovid, S., Schweiger, O., Milid, D., Harpke, A., & Vujid, A. (2017). Living on the edge: Forecasting the trends in abundance and distribution of the largest hoverfly genus (Diptera: Syrphidae) on the Balkan Peninsula under future climate change. Biological Conservation, 212(1), 216-229. https://doi.org/10.1016/j.biocon.2017.06.026. DOI: https://doi.org/10.1016/j.biocon.2017.06.026
Reina-Ávila, D., Riaño-Jiménez, D., Aguilar, L. & Cure, JR. (2013). Visitantes florales (Arthropoda: Insecta) en zona de sub-páramo en los cerros orientales de la sabana de Bogotá, Colombia. Revista de Entomología Mexicana, 12, 599-604.
Sánchez-Bayo, F., & Wyckhuys, K. A. (2019). Worldwide decline of the entomofauna: A review of its drivers. Biological Conservation, 232, 8-27. https://doi.org/10.1016/j.biocon.2019.01.020 DOI: https://doi.org/10.1016/j.biocon.2019.01.020
Sánchez-N., D., & Amat-García, G. (2005). Diversidad de la fauna de artrópodos terrestres en el Humedal Jaboque, Bogotá-Colombia. Caldasia, 27(2), 311-329.
Saunders, M. E. (2018). Insect pollinators collect pollen from wind‐pollinated plants: Implications for pollination ecology and sustainable agriculture. Insect Conservation and Diversity, 11(1), 13-31. https://doi.org/10.1111/icad.12243 DOI: https://doi.org/10.1111/icad.12243
Schleuning, M., Fründ, J., Klein, A. M., Abrahamczyk, S., Alarcón, R., Albrecht, M., Andersson, G., Bazarian, S., Dalsgaard, B., Böhning-Gaese, K., Bommarco, R., Dalsgaard, B., Dehling, D. M., Gotlieb, A., Hagen, M., Hickler, T., Holzschuh, A., Kaiser-Bunbury, C. N., Kreft, H ... Blüthgen, N. (2012). Specialization of mutualistic interaction networks decreases toward tropical latitudes. Current Biology, 22(20), 1925-1931. https://doi.org/10.1016/j.cub.2012.08.015 DOI: https://doi.org/10.1016/j.cub.2012.08.015
Stroo, A. (2000). Pollen morphological evolution in bat pollinated plants. Plant Systematics and Evolution, 222, 225-242. DOI: https://doi.org/10.1007/978-3-7091-6306-1_12
Thébault, E. & Fontaine, C. (2010). Stability of ecological communities and the architecture of mutualistic and trophic networks. Science, 3291, 853–856. https://doi.org/10.1126/ science.1188321. DOI: https://doi.org/10.1126/science.1188321
Velásquez, C.A. (1999). Atlas palinológico de la flora vascular paramuna colombiana: Angiospermae. Universidad Nacional.
Welti, E. A., & Joern, A. (2015). Structure of trophic and mutualistic networks across broad environmental gradients. Ecology and Evolution, 5(2), 326-334. https://doi.org/10.1002/ece3.1371 DOI: https://doi.org/10.1002/ece3.1371
Zamora, M., Amat G. D., & Fernández J. L. (2011). Estudio de las visitas de las moscas de las flores (Diptera: Syrphidae) en Salvia bogotensis (Lamiaceae) en el Jardín Botánico José Celestino Mutis (Bogotá D.C., Colombia). Caldasia, 33(2), 453-470.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Recibido: 4 de junio de 2023; Aceptado: 15 de noviembre de 2023
Resumen
Las moscas de las flores son polinizadores fundamentales en hábitats de alta montaña como el bosque altoandino. Evaluamos la relación planta-sírfido asociada al consumo de polen en tres coberturas vegetales con diferentes niveles de influencia antrópica en una microcuenca cercana a Bogotá. Posterior a la extracción y acetólisis de los tractos digestivos de las 22 especies de sírfidos (n=358 especímenes), el polen encontrado se caracterizó en función de sus dimensiones y ornamentos, mientras que los espectros polínicos se analizaron empleando redes de interacción bipartitas para determinar los grados de especialización, vulnerabilidad, conectividad y anidamiento de cada red-cobertura. Las relaciones de los sírfidos en las coberturas estudiadas se mostraron generalistas. Se encontró una mayor vulnerabilidad en la vegetación secundaria alta y menor estabilidad de la red bipartita en el tejido urbano discontinúo, en contraste con la red del bosque ripario. Esto sugiere una menor resiliencia de coberturas antropizadas y sucesiones intermedias ante eventos de extinción.
Palabras clave:
anidamiento, coberturas antropizadas, conectividad, redes bipartitas, resiliencia..Abstract
Flower flies are fundamental pollinators in high mountain habitats such as high Andean forests. We evaluated the plant-syrphid relationship associated with pollen consumption in three vegetation covers with different levels of anthropic influence in a micro-watershed near Bogotá. After extracting and performing acetolysis the digestive tracts of 22 syrphid species (n = 358 specimens), the pollen found was characterized and categorized according to its size and ornamentation, while the pollen spectra were analyzed by means of bipartite interaction networks to assess the degrees of specialization, connection, vulnerability, and nestedness of each network. The relationships of the syrphids in the studied covers appeared to be generalist in nature. Greater vulnerability was observed in tall secondary vegetation, and less stability was found in the discontinuous urban fabric's bipartite network when compared to that of the riparian forest. This suggests a lower resilience for the anthropized covers and intermediate successions in the face of extinction events.
Keywords:
nestedness, anthropized coverage, connectivity, bipartite networks, resilience..INTRODUCCIÓN
Los sírfidos, también conocidos como moscas de las flores, desempeñan funciones importantes en diversos ecosistemas, tales como como el reciclaje de nutrientes, el control biológico de plagas y la polinización (Orford et al., 2015; Hughes, 2016). El servicio ecosistémico de polinización puede ser evaluado desde el enfoque de redes bipartitas (consumidores: sírfidos, productores: plantas), las cuales informan sobre el grado de especialización en escalas espaciales, temporales y de niveles tróficos (Jordano et al., 2009). Con base en este tipo de análisis, indicadores como la especialización muestran la predilección de especies entre niveles tróficos. De esta forma, los sírfidos especialistas consumen polen de una o pocas especies de plantas, lo que indica una menor tolerancia a la extinción de sus plantas nutricias y una mayor vulnerabilidad frente a perturbaciones ecosistémicas (Menke et al., 2012; Schleuning et al., 2012).
A nivel funcional, los sírfidos presentan diferencias en su generalísimo y su potencial como polinizadores. Las especies de la subfamilia Syrphinae suelen ser más pequeñas y presentan poca pilosidad y un acicalamiento más frecuente, por lo que el polen acumulado en sus cuerpos es menor y, por tanto, no suelen ser percibidas como polinizadoras. Por otra parte, las especies de la subfamilia Eristalinae tienden a ser más grandes, desarrollan mayor pilosidad y presentan una menor frecuencia en su acicalamiento, por lo que se asocian más con la polinización (Holloway, 1976). Respecto al tamaño de los sírfidos, se ha identificado que las especies grandes tienden a desplazar a las más pequeñas y a establecerse en inflorescencias de mayor tamaño, mientras que las pequeñas pueden al acceder al polen y el néctar de flores con corolas más profundas (Branquart & Hemptinne, 2000).
Si bien los sírfidos son considerados buenos polinizadores en los bosques altoandinos colombianos, son escasos los estudios sustentados en el análisis del polen adherido a sus cuerpos (Sánchez-N & Amat-García, 2005). Se han confirmado visitas florales por parte de estos a algunas especies nativas del bosque andino, tales como ‘mora silvestre’ y ‘mano de oso’ (Rubus robustus C. Presl. y Oreopanax sp.) (Reina-Ávila et al., 2013). Se destacan las relaciones con Salvia bogotensis Benth (Zamora et al., 2011) y Persea americana Mill (Carabalí et al., 2017), las cuales son visitadas por 9 y 20 morfoespecies de moscas de las flores respectivamente.
Aunque se ha señalado que las moscas de las flores son mayoritariamente generalistas (Klecka et al., 2018), lo que las hace estratégicas en procesos de restauración (Radenkovid et al., 2017), en los Andes aún no se comprende cómo la antropización de sus coberturas puede afectar las relaciones entre ellas y las plantas desde la perspectiva del consumo de polen.
En los Cerros Orientales de Bogotá DC, el cambio de uso de suelo ha causado el deterioro ecológico de la cobertura vegetal natural por cuenta de la urbanización, la potrerización y la introducción de especies invasoras, motores de pérdida de biodiversidad (Sánchez-Bayo & Wyckhuys, 2019). Por lo tanto, es pertinente saber cómo los cambios antropogénicos en las coberturas vegetales influyen en las asociaciones entre sírfidos y plantas, con el fin de tomar decisiones informadas sobre la gestión y conservación de estas moscas y sus funciones ecosistémicas. Con esto en mente, este estudio evaluó la relación funcional a partir del consumo de polen por parte de los sírfidos y las características morfológicas tanto de las moscas como del polen. Esto, en tres coberturas vegetales con diferentes niveles de influencia antrópica en la cuenca de la Quebrada Las Delicias de Bogotá.
MATERIALES Y MÉTODOS
Área de estudio y muestreo
Para este estudio, se emplearon datos publicados por Ángel et al. (2021) en conjunto con información inédita relacionada con muestreos activos de sírfidos en las tres coberturas vegetales identificadas en la Quebrada Las Delicias, Bogotá, Colombia (coordenadas 4°38’23.36” Norte, 74°3’16.84” Oeste y 4°38’14.20” Norte, 74°3’7.25” Oeste; World Geodetic System 1984, WGS 84). Los autores realizaron una fotointerpretación de las coberturas sustentada en la historia ambiental del área y la leyenda de coberturas de uso de suelo para Colombia del Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM (2010). Las coberturas estudiadas fueron: tejido urbano discontinuo (TUD) con una alta incidencia antrópica; vegetación secundaria alta (VSA), producto de la regeneración vegetal no gestionada, posterior a la tala de eucaliptos; y bosque ripario (BR), un área de restauración gestionada en la cual se han sembrado especies vegetales de importancia ecológica en la Quebrada.
Los muestreos activos se realizaron entre agosto y octubre de 2018, durante los periodos de máxima actividad de estos dípteros. Todos los insectos fueron recolectados utilizando una red entomológica, realizando recorridos aleatorios en cada una de las coberturas. Los sírfidos capturados fueron preservados individualmente en tubos Eppendorf con etanol, a una temperatura de 4 °C. En total se recolectaron 381 individuos adultos. Sin embargo, solo a 371 individuos se les realizó un análisis del espectro polínico, y a 358 se les realizó un análisis morfométrico. Producto de este último análisis, se diferenciaron tres grupos de tamaño para los sírfidos: grandes, medianos y pequeños (Anexo 1).
Extracción y caracterización del polen consumido
Antes de extraer y concentrar el polen, se diseccionó el tracto digestivo de cada sírfido utilizando un estereomicroscopio. Posteriormente, los tractos fueron acetolizados siguiendo la técnica desarrollada por Erdtman (1986). Finalmente, una alícuota del polen concentrado por insecto fue montada en una lámina portaobjetos junto con gelatina glicerinada y sellada con parafina. Para la identificación de los granos de polen, se utilizó un microscopio Zeiss Primo Star. Siempre que fue posible, el análisis del espectro polínico se sustentó en el conteo de al menos 300 granos de polen por muestra-individuo. Este número ha sido tradicionalmente usado en palinología para caracterizar la riqueza de polen presente en las muestras. El polen encontrado fue identificado por contraste con el polen de referencia de la zona y con atlas palinológicos de alta montaña andina (Bogotá et al., 1996; Velásquez, 1999; Bogotá-A, 2002). El polen consumido se agrupó según su ornamentación y tamaño, de acuerdo con las categorías propuestas en Bogotá et al. (1996) (Anexo 2).
Análisis de las redes mutualistas planta-animal con base en el consumo de polen
Para evaluar el grado de especialización en la interacción planta-sírfido polinizador por tipo de cobertura vegetal, se utilizaron todos los insectos a lo que se les pudo extraer polen. Se analizaron los datos de abundancia del polen consumido por las especies de sírfidos para elaborar redes de interacción bipartitas cuantitativas, generadas con el paquete bipartite en Rstudio versión 1.0.153 (R Development Core Team, 2015). Se calcularon varios índices para estas redes, incluyendo la conectividad (la proporción de enlaces establecidos en relación a los posibles), la asimetría de la red (el indicador de dependencia hacia los consumidores o productores), el anidamiento ponderado (que representa la tendencia de los especialistas a interactuar con especies de plantas generalistas, donde los valores cercanos a 0 indican una red no anidada y los cercanos a 100 indican una red altamente anidada), la especialización de la red H 2 y la especialización a nivel de especie d' (que refleja la afinidad de una especie hacia un recurso/consumidor específico). Por último, se realizaron simulaciones de extinción en escenarios de pérdida de especies tanto de plantas como de sírfidos (Dormann et al., 2019). Se generaron diagramas boxplot y se realizó un análisis ANOVA mediante el paquete The R Stats Package (R Development Core Team, 2015) para entender el grado de especialización de los sírfidos y del polen. En estos análisis se emplearon las variables de tamaño y subfamilia para los primeros, y tamaño y ornamentación para los segundos.
RESULTADOS
Las relaciones planta-sírfido identificadas incluyen 50 especies de plantas y 22 especies de sírfidos distribuidas en las tres coberturas estudiadas (Figura 1, Anexo 1). De los 358 especímenes de sírfidos disecados, el espectro polínico está sustentado en más de 300 granos para 285. En 66 especímenes se encontraron entre 1 y 299 granos, y solo en 20 individuos fue imposible extraer polen. En total se contaron 90 731 granos de polen, de los cuales, en función de su ornamentación, 38 088 fueron clasificados como espinados, 16 255 como reticulados, 16 195 como fenestrados, 12 331 como microreticulados, 5989 como psilados y 1873 como gemados. En relación con el tamaño, 87 229 granos fueron medianos, 2261 grandes y 1241 pequeños (Anexo 2).
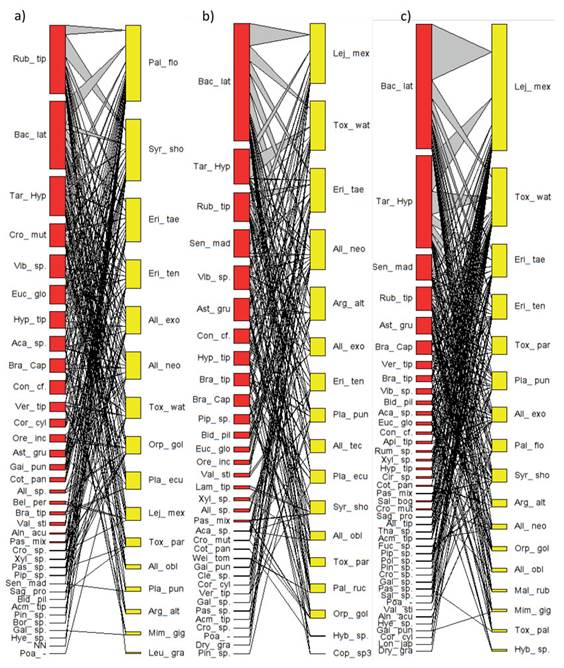
Figura 1: Redes mutualistas de consumo de polen por sírfidos.
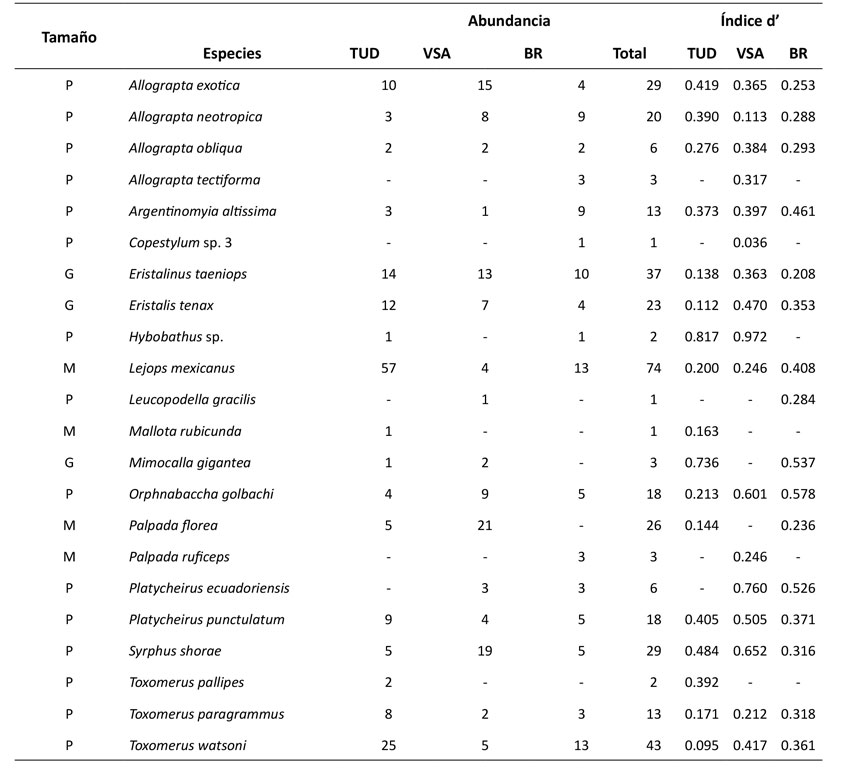
En las redes pertenecientes al TUD y VSA, ocurren 17 especies de sírfidos, mientras que en BR hay 16 (Figura 1). En las redes VSA y BR, cinco y tres especies, respectivamente, presentaron una mayor tendencia a la especialización (d’>0.5) (Tabla 1). En cuanto al componente vegetal, la red de TUD involucra 42 palinomorfos-plantas diferentes, mientras que las de VSA y BR incluyen 34 y 36 palinomorfos. De estas siete, nueve y cinco plantas-palinomorfos presentaron, respectivamente, una mayor tendencia a la especialización (Figura 1, Tabla 1).
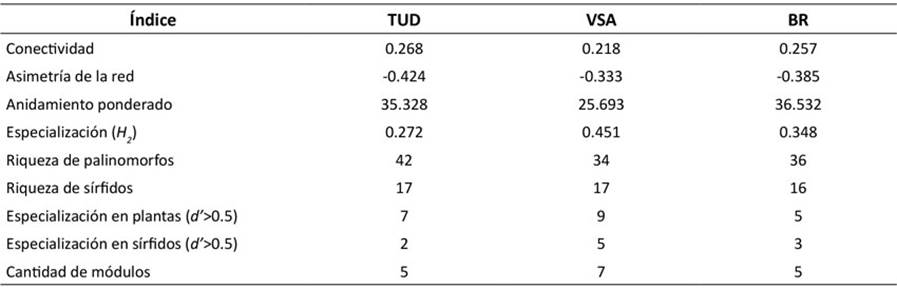
Tabla 1: Índices de interacción para las redes planta-sírfido asociadas a las tres coberturas analizadas en la Quebrada Las Delicias. Coberturas TUD: tejido urbano discontinuo, VSA: vegetación secundaria alta, BR: bosque ripario
Una comparación basada en los sírfidos entre las redes tróficas de las tres coberturas (Figura 1) señala una mayor semejanza entre TUD y VSA. En estas coberturas vegetales, las especies de sírfidos que más interacciones acumulan son Lejops mexicanus (Macquart, 1842), especialmente en TUD, y Toxomerus watsoni (Curran, 1930). En BR, estas especies tienen una participación menos relevante. En esta última cobertura, Palpada florea (Hull, 1925) y Syrphus shorae (Fluke, 1950) son los sírfidos que más interacciones acumularon. Respecto a los palinomorfos, las Asteraceas Baccharis latifolia (Ruiz & Pav.) Pers. y Taraxacum-Hipochaeris son las plantas que más interacciones acumularon en las tres coberturas vegetales. Se resalta el papel de Rubus en BR y VSA (Figura 1). Particularmente en BR, la acumulación de red se ve acompañada por dos especies arbóreas: Croton mutisianus Kunth y Eucalyptus globulus Labill.
En cuanto a los atributos de interacción de las tres coberturas, TUD exhibió la mayor conectividad, y riqueza de palinomorfos, pero la menor asimetría. El mayor anidamiento ponderado lo registró el BR, mientras que la cobertura con mayores valores de especialización (H 2 y d’) fue VSA, en contraste con BR (Tabla 1). En cuanto a los sírfidos, la riqueza y abundancia de especies fue similar en las tres coberturas vegetales estudiadas.
La ornamentación de polen más común fue la espinada (41.9; Asteraceae) seguida de la fenestrada (17.9%; Asteraceae) y el reticulado (17.8%; Brassicaceae, Adoxaceae). Por su parte, el tamaño más común fue el mediano (96.1%), seguido por el pequeño y el grande (1.3%). Respecto a los análisis de especialización d’, se encontró que la ornamentación del polen tiende a ser superior en los microreticulados, psilados y reticulados (i.e., Rubus, Alnus, Passiflora), en contraste con los clavados (Croton) (Figura 2; Pr(>F) 0.011). En particular, la subfamilia Syrphinae presentó una mayor tendencia a la especialización frente a Eristalinae (Pr(>F) 0.014). Respecto al tamaño de los sírfidos, se encontró que las especies más pequeñas tienen una mayor tendencia a la especialización (Figura 2; Pr(>F) 0.0024). Esta condición también fue identificada en el tamaño de los granos de polen Pr (>F) 0.006). Estos resultados fueron reafirmados con un ANOVA, que mostró diferencias significativas para todas las variables (Anexos 4 y 5).
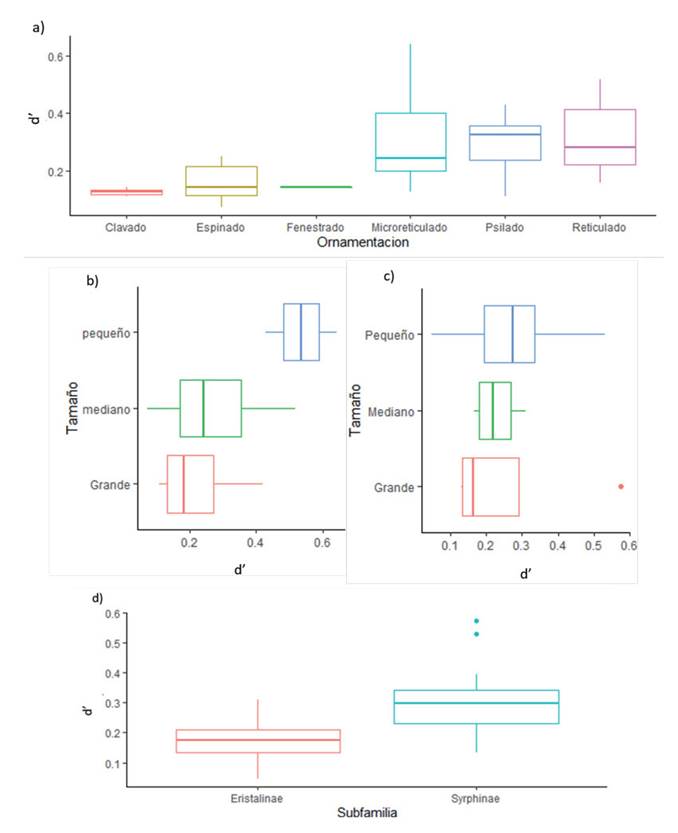
Figura 2: Boxplots de las variables morfométricas del polen consumido y de los sírfidos en las tres coberturas estudiadas en función del grado de especialización d’.
Se presenta el grado de especialización a) según la ornamentacion del polen, b) según el tamaño del grano de polen, c) según el tamaño de los insectos y d) según la subfamilia de Sirfidae.
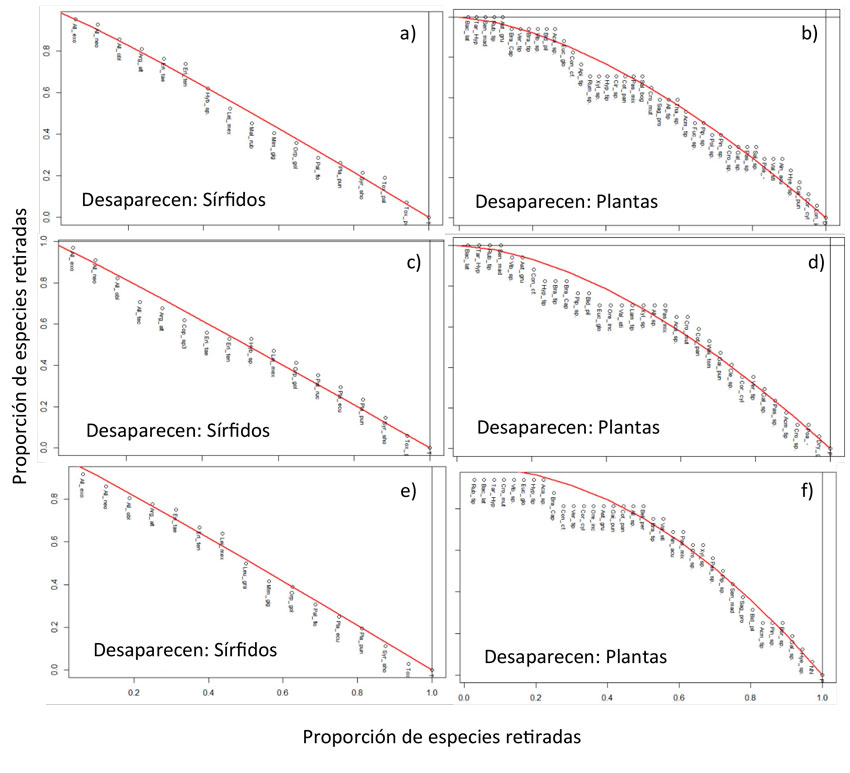
En lo que respecta a la tolerancia a la extinción de las tres redes estudiadas, las simulaciones de extinciones locales revelaron que, al desaparecer las especies de las comunidades de sírfidos en las tres redes, las plantas se extinguirían al mismo ritmo. En cambio, se evidencia que, con la desaparición de las especies vegetales en las tres redes, habría una reducción en la proporción de pérdida de especies de sírfidos. Con ello, sería TUD la cobertura que presentaría la mayor proporción de pérdida, seguida de VSA y BR (Anexo 6).
DISCUSIÓN
De las tres coberturas vegetales diferenciadas en el sector de la Quebrada Las Delicias (Ángel et al., 2021), es la vegetación secundaria alta (VSA: H 2 = 0,45) la que mayor susceptibilidad a la extinción presenta frente al bosque ripario (BR) y al tejido urbano discontinuo (TUD), que reflejan mayor cercanía a condiciones naturales y a afectación antropogénica respectivamente. La especialización en VSA está relacionada con el mayor número de especies de plantas y sírfidos que se muestran especialistas y con el respectivo número de módulos de red encontrados (Tabla 1, Anexo 3), además de concordar con lo planteado por Menke et al. (2012), quienes sugirieron que las vegetaciones en transición tienden a generar estrategias especializadas en la regulación planta-animal. Más aún, al presentar los niveles más bajos de interconexión y de especies anidadas, esta red tiene una baja capacidad para resistir extinciones (Memmot et al., 2004; Thébault & Fontaine, 2010; Schleuning et al., 2012; Lever et al., 2014). Por su parte, si bien BR y TUD presentaron una conectividad similar, BR se mostró más especialista y con mayor anidamiento ponderado. Esto plantearía una mayor estabilidad en su comunidad, pues está más conservada, una condición que coincide con lo propuesto por Thébault & Fontaine (2010) y Dormann (2011).
Las comunidades de sírfidos en la Quebrada Las Delicias son, en su mayoría, generalistas (d’ <0.5; Anexo 1). Sin embargo, al analizar de manera detallada el índice de especialización a nivel de especie en cuanto a subfamilia y tamaño, se identificó que la especialización podría estar más asociada a las características propias de la subfamilia que al tamaño. De manera general, se encontró que los sírfidos de tamaño pequeño presentan un mayor nivel de especialización, una característica propuesta por Branquart & Hemptinne (2000) al plantear que los sírfidos pequeños son desplazados por los grandes a flores con nectarios profundos, pues estos últimos más grandes no pueden acceder a este recurso floral. No obstante, Zamora et al. (2012) identificaron que algunos sírfidos pequeños como P. punctulatum poseen aparatos bucales proporcionalmente más grandes que aquellos de tamaño corporal más grande como S. shorae (3.66 y 1.89 mm respectivamente). Respecto a las subfamilias de sírfidos en el área estudiada, Syrphinae fue menos generalista que Eristalinae, una característica también registrada en pastizales de Gales, Reino Unido, donde las moscas de Eristalinae tienden a ser polinizadoras eficientes y generalistas (Lucas et al., 2018). Esta tendencia hacia la especialización estaría asociada a los comportamientos de las subfamilias. Holloway (1976) identificó que las moscas de Syrphinae presentan mayor acicalamiento y, a su vez, una mayor recolección del polen en sus cuerpos, por lo que tienen que buscar asiduamente más recursos florales, contrario a las moscas de Eristalinae, que son visitantes florales mucho más activos y, por tanto, generalistas.
Para explicar la mayor especialización de la subfamilia Syrphinae respecto a la de Eristalinae, sería conveniente incluir dos factores importantes como la latitud y la representatividad del muestreo por subfamilia. Esto, ya que existen indicios que muestran que la especialización de comunidades de sírfidos tiende a ser mayor en países de mayores latitudes frente a los tropicales (Schleuning et al., 2012; Welti & Joern, 2015; Dalsgaard et al., 2017). En la Republica Checa se ha registrado un comportamiento especialista en los miembros de Eristalinae en comparación con los de Syprhinae (Klecka et al., 2018). Por otra parte, las especies M. gigantea, O. golbachi, P. ecuadoriensis, P. punctulatum y S. shorae e Hybobathus sp., todas de la familia Syrphinae, presentan tendencias especialistas, lo que podría ser reflejo de su baja representación (Petanidou et al., 2008; Dormann, 2011).
En general, los sírfidos presentaron predilección por el consumo de polen de plantas con abundante recurso floral (número de flores, presencia de nectarios, producción de polen), por lo que estos resultados respaldarían la teoría de búsqueda óptima simple señalada por Holloway (1976). En las tres coberturas, los sírfidos consumieron ante todo polen proveniente de flores de las familias botánicas Asteraceae, Rosaceae y Brassicaceae, cuya morfología es espinada, fenestrada y microrreticulada respectivamente, así como de tamaño mayoritariamente mediano. Sin embargo, se encontró consumo de polen de Alnus, Thalictrum, Vallea, Piper, Poacea y Pinnacea, entre otros, cuyo síndrome de dispersión puede ser considerado como anemófilo. Con lo anterior, los resultados obtenidos resultados contrastarían parcialmente con lo planteado por Stroo (2000) y Hu et al. (2008), quienes afirman que el polen evolucionó con estrategias hacia un mayor tamaño y ornamentos para adherirse a los insectos y así garantizar la polinización. En este sentido, estos insectos parecerían tener predilección por plantas con polen ambófilo, que pueden ser polinizadas tanto por el viento como por insectos (Saunders, 2018). Sin embargo, la diversidad en el consumo de polen depende de la composición, la oferta del recurso y, así, del grado de conservación de las coberturas, de modo que las coberturas alteradas con baja riqueza de estos recursos tienden a hacer que las comunidades de sírfidos presenten menor diversidad y riqueza (Gervais et al., 2018). Este planteamiento es congruente con lo encontrado por Ángel et al. (2021), donde la mayor diversidad en términos de especies comunes (q=1) y dominantes (q=2) de sírfidos se reportó en BR. Esto, en contraste con TUD y VSA.
CONCLUSIÓN
Este es el primer estudio en el que se analizan las interacciones entre los sírfidos y las comunidades vegetales (polen consumido) en áreas altoandinas con intervención humana en Colombia. En las tres coberturas diferenciadas, las relaciones asociadas a la dieta de los sírfidos muestran una tendencia generalista a nivel de red, lo que hace que estas redes puedan ser menos vulnerables ante futuros eventos de extinción. En general los sírfidos de la subfamilia Syrphinae presentan una mayor afinidad a hacia la especialización a nivel de especie, una característica que refleja su comportamiento de acicalamiento.
Se destaca una menor vulnerabilidad en los sírfidos ante eventos de extinción por cuenta de la desaparición de plantas en la red del BR, lo que sería producto de un mayor anidamiento. En contraste, la red con mayor vulnerabilidad sería la VSA, que, al estar en estados intermedios de conservación, puede presentar relaciones planta-animal más especializadas con mayor frecuencia.
Por último, estos insectos obtienen polen de diversos tamaños y ornamentaciones pertenecientes a plantas de diversas especies -incluso de plantas potencialmente ambófilas-, lo que hace que puedan tolerar extinciones locales de plantas, al poder reemplazar con mayor facilidad sus recursos alimenticios.
Acknowledgements
AGRADECIMIENTOS
Los autores agradecen a los docentes de la Universidad Distrital Francisco José de caldas Olga Patricia Pinzón Florián y Diego Tomás Corradine Mora por facilitar los equipos y espacios necesarios en el marco de la presente investigación. Así mismo, a Ángela Parrado Roselli y Marcia Carolina Muñoz Neyra por sus comentarios críticos y sus aportes a las versiones preliminares de esta investigación.
REFERENCIA
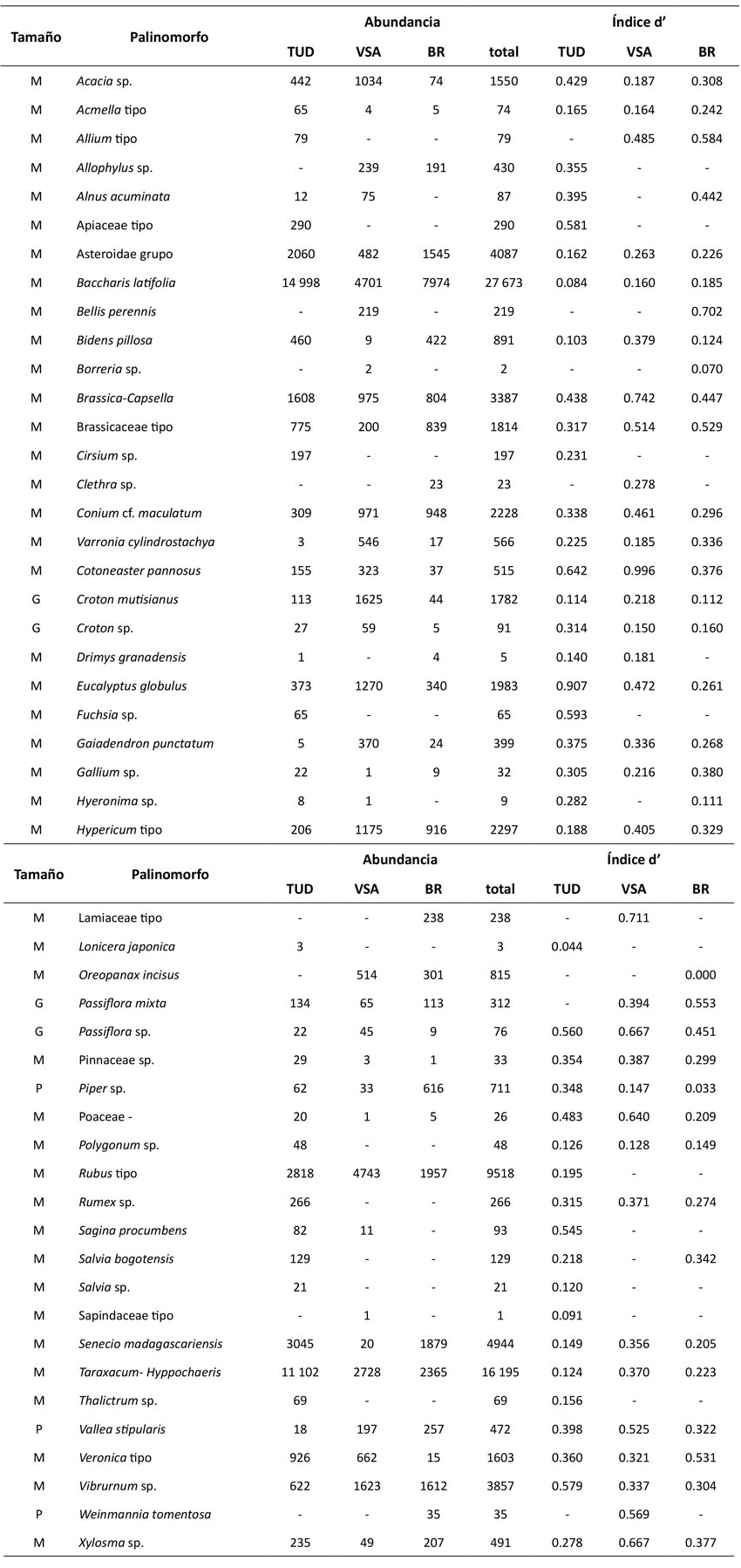
Anexo 1: Grado de especialización (d’) de los insectos del conteo de 300 granos por coberturas. Los valores cercanos a 0 representan insectos generalistas y los cercanos a 1 representan insectos especialistas. Tamaño de los insectos: P = pequeño, M = mediano y G = grande. TUD: tejido urbano discontinuo; VSA: vegetación secundaria alta y BR: bosque ripario.
Anexo 2: Grado de especialización (d’) de las plantas del conteo de 300 granos. Los valores cercanos a 0 representan insectos generalistas y los cercanos a 1 representan plantas especialistas. Tamaño de los insectos: P = pequeño, M = mediano y G = grande. TUD: tejido urbano discontinuo; VSA: vegetación secundaria alta y BR: bosque ripario. Tamaño de los granos de polen: P = pequeño, M = mediano y G = grande.
Anexo 3: Graficas de modularidad generadas para cada una de las redes de cada cobertura. a) tejido urbano discontinuo; b) vegetación secundaria alta; c) bosque ripario. Abreviaturas en la Figura 1.
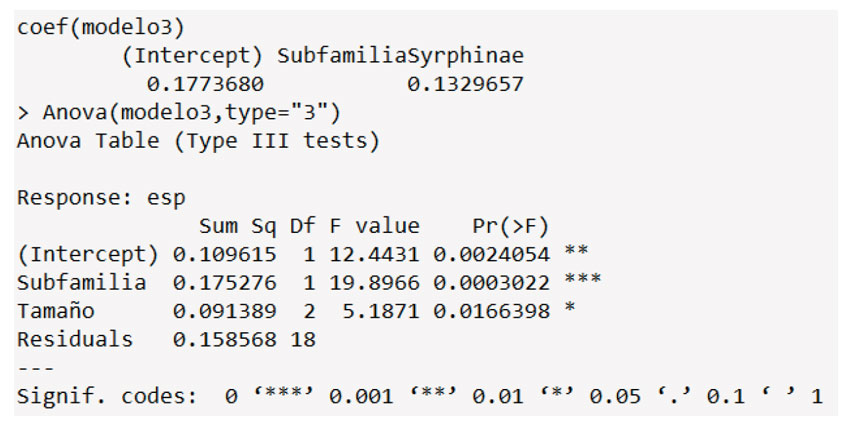
Anexo 4: Captura de pantalla de la salida del ANOVA realizado para la especialización respecto la subfamilia de sírfidos.
Anexo 5: Captura de pantalla de la salida del ANOVA realizado para la especialización respecto a la ornamentación del polen y el tamaño.

Anexo 6: Gráficas de proporción de extinción a raíz de la extinción local de especies de plantas y sírfidos. a, c y e muestran el grado de extinción de las plantas si desaparecen estos dípteros; b, d y f muestran el grado de extinción de los sírfidos si desaparecen las plantas. a y b presentan las gráficas del TUD; c y d corresponden al VSA, y e y f corresponden a BR.




Licencia
Derechos de autor 2024 Colombia Forestal

Esta obra está bajo una licencia internacional Creative Commons Atribución-CompartirIgual 4.0.
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






