
DOI:
https://doi.org/10.14483/2256201X.20968Publicado:
01-01-2024Número:
Vol. 27 Núm. 1 (2024): Enero-junioSección:
Artículos de investigación científica y tecnológicaComposición, estructura y diversidad florística del bosque Monteleón: Un relicto urbano del bosque subandino (Manizales, Colombia)
Composition, Structure, and Floristic Diversity of the Monteleón Forest: An Urban Remnant of the Sub-Andean Forest (Manizales, Colombia)
Palabras clave:
Abundancia, heterogeneidad, número efectivo de especies, perturbación, riqueza (es).Palabras clave:
Abundance, heterogeneity, effective number of species, disturbance, richness (en).Descargas
Referencias
Alcaldía de Manizales (2018). Oficio SCIA-246-218, Control y seguimiento de movimiento de tierras. Secretaría de Planeación, Alcaldía de Manizales. https://manizales.gov.co/RecursosAlcaldia/201901041238160361.pdf
Alexiades, M. N. (1996). Standard techniques for collecting and preparing herbarium specimens. Advances in Economic Botany, 10, 99–126. http://www.jstor.org/stable/43927613
Álvarez-Mejía, L. M., Sanín, D., Alzate-Quintero, N. F., Castaño-R., N., Mancera-Santa, J. C., & Gonzáles-Ocampo, G. (2007). Plantas de la región centro-sur de Caldas (1ra ed.). Universidad de Caldas.
Ángeles-García, A. Y., Suárez-Torres, J. J., Carrillo-Reyes, P., Peña-Joya, K. E., & Quijas, S. (2022). Diversidad de plantas leñosas en el bosque tropical caducifolio y subcaducifolio de Sierra de Vallejo, Nayarit, México. Botanical Sciences, 100(1), 2-27. https://doi.org/10.17129/BOTSCI.2770 DOI: https://doi.org/10.17129/botsci.2770
Ariza-Cortés, W., Toro-Murillo, J. L., & Lores-Medina, A. (2009). Análisis florístico y estructural de los bosques premontanos en el municipio de Amalfi (Antioquia, Colombia). Revista Colombia Forestal, 12, 81-102. https://doi.org/10.14483/udistrital.jour.colomb.for.2009.1.a07 DOI: https://doi.org/10.14483/udistrital.jour.colomb.for.2009.1.a07
Banhos, A., Hrbek, T., Sanaiotti, T. M., & Farias, I. P. (2016). Reduction of genetic diversity of the Harpy Eagle in Brazilian tropical forests. PloS ONE, 11(2), 0148902. https://doi.org/10.1371/journal.pone.0148902 DOI: https://doi.org/10.1371/journal.pone.0148902
Baselga, A. (2013). Separating the two components of abundance-based dissimilarity: Balanced changes in abundance vs. abundance gradients. Methods in Ecology and Evolution, 4(6), 552-557. https://doi.org/10.1111/2041-210X.12029 DOI: https://doi.org/10.1111/2041-210X.12029
Baselga, A., & Orme, C. D. L. (2012). Betapart: An R package for the study of beta diversity. Methods in Ecology and Evolution, 3, 808-812. https://doi.org/10.1111/j.2041-210X.2012.00224.x DOI: https://doi.org/10.1111/j.2041-210X.2012.00224.x
Bray, J. R., & Curtis, J. T. (1957). An ordination of the upland forest communities of Southern Wisconsin. Ecological Monographs, 27(4), 325-349. https://doi.org/10.2307/1942268 DOI: https://doi.org/10.2307/1942268
Castaño, J. H., Cardona Ramírez, D., & Botero, J. E. (2010). Ecología del mono nocturno andino (Aotus lemurinus) en fragmentos de bosque subandinos de Colombia. En V. Pereira-Bengoa, P. R. Stevenson, M. L. Bueno & Fernando Nassar-Montoya (Eds.), Primatolgía en Colombia: avances al principio del milenio (1ra ed., pp. 69-90). Fundación Universitaria San Martín.
Chao, A., & Jost, L. (2012). Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology, 93(12), 2533-2547. https://doi.org/10.1890/11-1952.1 DOI: https://doi.org/10.1890/11-1952.1
Cultid-Medina, C. A., & Escobar, F. (2019). Pautas para la estimación y comparación estadística de la diversidad biológica. En C. E. Moreno (Ed.), La biodiversidad en un mundo cambiante: fundamentos teóricos y metodológicos para su estudio (pp. 175-202). Libermex.
Cumming, G., Fidler, F., & Vaux, D. L. (2007). Error bars in experimental biology. The Journal of Cell Biology, 177(1), 7-11. https://doi.org/10.1083/jcb.200611141 DOI: https://doi.org/10.1083/jcb.200611141
Dirzo, R., & Raven, P. H. (1994). Un inventario biológico para México. Boletín de La Sociedad Botánica de México, 55, 29-34. https://doi.org/10.17129/botsci.1444 DOI: https://doi.org/10.17129/botsci.1444
Donovan, T. M., & Flather, C. H. (2002). Relationships among North American songbird trends, habitat fragmentation, and landscape occupancy. Ecological Applications, 12(2), 364-374. DOI: https://doi.org/10.1890/1051-0761(2002)012[0364:RANAST]2.0.CO;2
Dueñas-C., A., Betancur, J., & Galindo-T., R. (2007). Estructura y composición florística de un bosque húmedo tropical del Parque Nacional Natural Catatumbo Barí, Colombia. Revista Colombia Forestal, 10(20), 26-39. https://doi.org/10.14483/udistrital.jour.colomb.for.2007.1.a02 DOI: https://doi.org/10.14483/udistrital.jour.colomb.for.2007.1.a02
Duque-Castrillón, C. A., & Sierra-Giraldo, J. A. (2022). Estructura y diversidad florística de dos bosques andinos en el Resguardo Indígena Nuestra Señora Candelaria de La Montaña (Riosucio, Caldas, Colombia). Boletín Científico Centro de Museos Museo de Historia Natural de La Universidad de Caldas, 26(2), 13-34. https://doi.org/10.17151/bccm.2022.26.2.1 DOI: https://doi.org/10.17151/bccm.2022.26.2.1
Duque-Escobar, G. (2015). El desarrollo urbano y económico de Manizales. https://godues.wordpress.com/2015/11/28/
Duque-Salazar, M. C., Alzate-Marín, J. J., & Otálvaro-Trejos, C. A. N. (2000). Problemática socio ambiental en la expansión urbana de Manizales [Tesis de especialización, Universidad de Manizales].
Echeverri-González, J. (1993). Monteleón: relicto de selva andina en Manizales. Anfora, 1(2), 45-53. DOI: https://doi.org/10.30854/anf.v1.n2.1993.463
El Tiempo (1993, noviembre 30). Montaña de lodo sepultó a 10 personas en Manizales. https://www.eltiempo.com/archivo/documento/MAM-269493
Fahrig, L. (2003). Effects of habitat fragmentation on biodiversity. Annual Review of Ecology, Evolution, and Systematics, 34, 487-515. https://doi.org/10.1146/annurev.ecolsys.34.011802.132419 DOI: https://doi.org/10.1146/annurev.ecolsys.34.011802.132419
Fahrig, L., Arroyo-Rodríguez, V., Bennett, J. R., Boucher-Lalonde, V., Cazetta, E., Currie, D. J., Eigenbrod, F., Ford, A. T., Harrison, S. P., Jaeger, J. A. G., Koper, N., Martin, A. E., Martin, J.-L., Metzger, J. P., Morrison, P., Rhodes, J. R., Saunders, D. A., Simberloff, D., Smith, A. C., … Watling, J. I. (2019). Is habitat fragmentation bad for biodiversity? Biological Conservation, 230, 179-186. https://doi.org/10.1016/j.biocon.2018.12.026 DOI: https://doi.org/10.1016/j.biocon.2018.12.026
Finegan, B. (1992). The management potential of neotropical secondary lowland rain forest. Forest Ecology and Management, 47, 295-321. DOI: https://doi.org/10.1016/0378-1127(92)90281-D
Forero, E. (1977). Instrucciones para coleccionar plantas. Notas Divulgativas (Instituto de Ciencias Naturales), 7, 1-29.
Fournier O., L. A., & Herrera de Fournier, M. E. (1977). La sucesión ecológica como un método eficaz para la recuperación del bosque en Costa Rica. Agronomía Costarricense, 1(1), 23-29.
Franco-Rosselli, P., Betancur, J., & Fernández-Alonso, J. L. (1997). Diversidad florística en dos bosques subandinos del Sur de Colombia. Caldasia, 19(1), 205-234.
García-Martínez, S., & Mercado-Gómez, J. D. (2017). Diversidad de briófitos en fragmentos de bosque seco tropical, Montes de María, Sucre, Colombia. Revista Mexicana de Biodiversidad, 88(4), 824-831. https://doi.org/10.1016/j.rmb.2017.10.035 DOI: https://doi.org/10.1016/j.rmb.2017.10.035
García-Morales, R., Moreno, C. E., & Bello-Gutiérrez, J. (2011). Renovando las medidas para evaluar la diversidad en comunidades ecológicas: el número de especies efectivas de murciélagos en el sureste de Tabasco, México. Therya, 2(3), 205-215. https://doi.org/10.12933/therya-11-47 DOI: https://doi.org/10.12933/therya-11-47
García-Ruenes, D. C., & Alzate-Guarín, F. A. (2021). El género Alchornea (Euphorbiaceae) en el departamento de Antioquia, Colombia. Biota Colombiana, 22(2), 2-15. https://doi.org/10.21068/c2021.v22n02a01 DOI: https://doi.org/10.21068/c2021.v22n02a01
Gentry, A. H. (1982). Patterns of neotropical plant species diversity. En M. K. Hecht, B. Wallace & G. T. Prance (Eds.), Evolutionary Biology (vol. 15, pp. 1-84). Springer. https://doi.org/10.1007/978-1-4615-6968-8_1 DOI: https://doi.org/10.1007/978-1-4615-6968-8_1
Gentry, A. H. (1993). A field guide to the families and genera of woody plants of Northwest South America (Colombia, Ecuador, Peru), with supplementary notes on herbaceous taxa. University of Chicago Press.
Gil-Fernández, E. A., Jiménez-Gómez, C., & Rivera-Gutiérrez, D. A. (2018). Análisis en la relación Desarrollo-Riesgo-Desastre, en la zona urbana del municipio de Manizales [Tesis de pregrado, Universidad Católica de Manizales]. https://repositorio.ucm.edu.co/handle/10839/2537
Gómez, D. (2004). Estructura y composición florística de los bosques premontanos del municipio de Anorí - Antioquia. Corantioquia.
Gómez-Ruiz, P. A., Tapia-Sedeño, A., Ramírez-García, B., Pinho, B. X., & Arroyo-Rodríguez, V. (2016). Diferenciación florística a lo largo de un gradiente de perturbación antrópica en un paisaje tropical fragmentado. The Mexican Naturalist, 7, 1-5. https://doi.org/10.6084/M9.FIGSHARE.2068728.V1
González-Plazas, J. L. (2009). Estado actual de la periurbanización y el hábitat periurbano en Manizales (Colombia). Cuadernos de Vivienda y Urbanismo, 2(3), 92-123.
Guariguata, M. R., & Ostertag, R. (2002). Sucesión secundaria. En M. R. Guariguata & G.H. Kattan (Eds.), Ecología y conservación de Bosques neotropicales (pp. 591-623). Editorial Tecnológica de Costa Rica.
Harrison, S., & Bruna, E. (1999). Habitat fragmentation and large-scale conservation: What do we know for sure? Ecography, 22(3), 225-232. DOI: https://doi.org/10.1111/j.1600-0587.1999.tb00496.x
Hill, M. O. (1973). Diversity and evenness: A unifying notation and its consequences. Ecology, 54(2), 427-432. DOI: https://doi.org/10.2307/1934352
ICN (2004 y continuamente actualizado). Instituto de Ciencias Naturales, Facultad de Ciencias, Universidad Nacional de Colombia. http://www.biovirtual.unal.edu.co
Jiménez-García, R., Rojas-Giraldo, J. E., & Ochoa-Botero, M. I. (2013). Estructura Ecológica de Manizales 2013-2025.
Jost, L. (2006). Entropy and diversity. Oikos, 113(2), 363-375. DOI: https://doi.org/10.1111/j.2006.0030-1299.14714.x
Jost, L. (2018). ¿Qué entendemos por diversidad? El camino hacia la cuantificación. Mètode Science Studies Journal, 98, 39-45. https://doi.org/10.7203/metode.9.11472 DOI: https://doi.org/10.7203/metode.9.11472
Jost, L., DeVries, P., Walla, T., Greeney, H., Chao, A., & Ricotta, C. (2010). Partitioning diversity for conservation analyses. Diversity and Distributions, 16, 65-76. https://doi.org/10.1111/j.1472-4642.2009.00626.x DOI: https://doi.org/10.1111/j.1472-4642.2009.00626.x
Jost, L., & González-Oreja, J. A. (2012). Midiendo la diversidad biológica: más allá del índice de Shannon. Acta Zoológica Lilloana, 56(3-14), 3-14.
Lehnert, M. (2009). Resolving the Cyathea caracasana complex (Polypodiopsida: Cyatheaceae). Stuttgarter Beiträge zur Naturkunde A, Neue Serie, 2, 409-445.
MacGregor-Fors, I., & Zuria, I. (2019). Ciudades vivas: Biodiversidad en ambientes urbanos. En C. E. Moreno (Ed.), La biodiversidad en un mundo cambiante: Fundamentos teóricos y metodológicos para su estudio (pp. 113-127). Libermex.
Maechler, M., Rousseeuw, P., Struyf, A., Hubert, M., & Hornik, K. (2022). cluster: Cluster Analysis Basics and Extensions. R package version 2.1.4.
Marcon, E., & Hérault, B. (2015). Entropart: An R package to measure and partition diversity. Journal of Statistical Software, 67(8), 1–26. https://doi.org/10.18637/jss.v067.i08 DOI: https://doi.org/10.18637/jss.v067.i08
Martínez-Calderón, V. M., Sosa-Ramírez, J., Siqueiros-Delgado, M. E., & Díaz-Núñez, V. (2021). Composición, diversidad y estructura de especies leñosas en los bosques templados de Monte Grande, Sierra Fría, Aguascalientes, México. Acta Botánica Mexicana, 128(e1829), 1-20. https://doi.org/10.21829/abm128.2021.1829 DOI: https://doi.org/10.21829/abm128.2021.1829
Méndez-Urbano, F., Sierra-Giraldo, J. A., Carlsen, M. M., Rodríguez-Rey, G. T., & Castaño-Rubiano, N. (2022). Anthurium caldasii: A new species of Araceae from Colombia and its phylogenetic relationships with other black-spathed Anthurium species. Brittonia, 74(4), 419-435. https://doi.org/10.1007/s12228-022-09722-y DOI: https://doi.org/10.1007/s12228-022-09722-y
Mendoza-Cifuentes, H. (2008). Compilación de los inventarios RAP de vegetación en Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Mendoza, H., & Ramírez, B. (2006). Guía ilustrada de géneros Melastomataceae y Memecylaceae de Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Universidad del Cauca.
Moreno, C. E., Barragán, F., Pineda, E., & Pavón, N. P. (2011). Reanálisis de la diversidad alfa: alternativas para interpretar y comparar información sobre comunidades ecológicas. Revista Mexicana de Biodiversidad, 82, 1249-1261. DOI: https://doi.org/10.22201/ib.20078706e.2011.4.745
Mosquera-Ramos, L. J., Robledo-Murillo, D., & Asprilla-Palacios, A. (2007). Diversidad florística de dos zonas de bosque tropical húmedo en el municipio de Alto Baudó, Chocó-Colombia. Acta Biológica Colombiana, 12(S), 75-90.
Orozco-Agudelo, J. A., & Sierra-Giraldo, J. A. (2022). Dinámicas de reclutamiento, mortalidad, diversidad y crecimiento de la vegetación vascular en restauración ecológica [Presentación en simposio]. Experiencias en Restauración Ecológica “Desde La Investigación a Los Procesos Comunitarios”, Universidad de Caldas, Serviambientales e Isagen, Manizales, Colombia.
Orozco-León, E. M., & Ocampo-Calderón, Y. (2010). Evaluación del estado actual de la estructura y composición de la flora del Parque Regional Natural Monteleón (Manizales - Caldas) debido a la reducción de su área por tala [Tesis de pregrado, Universidad de Caldas].
Pérez-Restrepo, M. del P. (2017). Plan de ordenamiento territorial de Manizales 2017-2031. Alcaldía de Manizales.
Posada-Herrera, J. M. (2013). Novedades corológicas de la familia Melastomataceae para el departamento de Caldas, Colombia. Boletín Científico Centro de Museos Museo de Historia Natural Universidad de Caldas, 17(2), 38-43. https://revistasojs.ucaldas.edu.co/index.php/boletincientifico/article/view/4484
POWO (2023). Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. http://www.plantsoftheworldonline.org/
R Core Team (2022). R: A language and environment for statistical computing. R Foundation for Statistical Computing. https://www.R-project.org/
Ramírez-Salazar, D. C. (2012). Caracterización florística y usos de la flora en la reservas forestales El Popal-Bolonias, La Linda y el área La Camelia en el Departamento de Caldas. Fundación Pangea, Manizales.
Rangel-Ch., J. O., & Velázquez, A. (1997). Métodos de estudio de la vegetación. En J. O. Rangel-Ch., P. D. Lowy C. & M. Aguilar P. (Eds.), Colombia diversidad biótica II. Tipos de vegetación en Colombia (pp. 59-82). Universidad Nacional de Colombia.
Restrepo de Fraume, M., Álvarez-Mejía, L. M., & Gallego-Aristizábal, J. H. (1990). Monteleón, relicto de selva andina tropical de Manizales, Colombia. Revista Agronomía, 4(1), 24-35.
Ríos-Málaver, C. (2007). Riqueza de especies de Mariposas (Hesperioidea & Papilionoideae) de la quebrada “El Águila” Cordillera Central (Manizales, Colombia). Boletín Científico Centro de Museos Museo de Historia Natural, 11, 272-291.
Ríos-Málaver, C. (2008). Mariposas (Hesperioidea y Papilionoidea) en un gradiente altitudinal de la reserva protectora de Río Blanco, Cordillera Central (Manizales, Colombia) (Informe número 1). Tropical Andean Butterfly Diversity Project. https://doi.org/10.13140/RG.2.1.3247.1442
Rojas-Morales, J. A., Escobar-Lasso, S., & Gutiérrez-Cárdenas, P. D. A. (2011). Contribución al conocimiento de los anfibios de la región Centro-Sur de Caldas: primeros registros de ranas de cristal (Anura: Centrolenidae) para el municipio de Manizales, Colombia. Boletín Científico Centro de Museos Museo de Historia Natural de La Universidad de Caldas, 15(1), 75-83.
Rudas, G., Marcelo, D., Armenteras, D., Rodríguez, N., Morales, M., Delgado, L. C., & Sarmiento, A. (2007). Biodiversidad y actividad humana: relaciones en ecosistemas de bosque subandino en Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Sánchez, F., Sánchez-Palomino, P., & Cadena, A. (2004). Inventario de mamíferos en un bosque de Los Andes Centrales de Colombia. Caldasia, 26(1), 291-309.
Sánchez-Cordero, V., Townsend Peterson, A., & Escalante-Pliego, P. (2001). El modelado de la distribución de especies y la conservación de la diversidad biológica. En H. Hernández (Ed.), Enfoques contemporáneos para el estudio de la biodiversidad (pp. 359-379). Instituto de Biología, UNAM.
Sánchez, O., Islebe, G.A., & Valdez, M. (2007). Flora arbórea y caracterización de gremios ecológicos en distintos estados sucesionales de la selva mediana de Quintana Roo. Foresta Veracruzana, 9(2), 17-26.
Sanín, D., Mancera-Santa, J. C., Castaño-Rubiano, N., Alzate-Q., N. F., Gonzáles-O., G., & Álvarez-M., L. M. (2006). Catálogo preliminar de plantas vasculares de la Reserva Forestal Protectora “Río Blanco” (Manizales, Caldas, Colombia). Boletín Científico Centro de Museos Museo de Historia Natural de La Universidad de Caldas, 10, 19-44.
Sierra-Giraldo, J. A., & Sanín, D. (2014). Aráceas de un bosque premontano en la cordillera central de Colombia. Boletín Científico Centro de Museos Museo de Historia Natural de La Universidad de Caldas, 18(2), 17-27.
Silva-Aparicio, M., Castro-Ramírez, A. E., Castillo-Campos, G., & Perales-Rivera, H. (2018). Estructura de la vegetación leñosa en tres áreas con Selva Baja Caducifolia en el Istmo-Costa de Oaxaca, México. Revista de Biología Tropical, 66(2), 863-879. DOI: https://doi.org/10.15517/rbt.v66i2.33419
Stevenson, P. R., & Aldana, A. M. (2008). Potential effects of ateline extinction and forest fragmentation on plant diversity and composition in the Western Orinoco Basin, Colombia. International Journal of Primatology, 29(2), 365-377. https://doi.org/10.1007/s10764-007-9177-x DOI: https://doi.org/10.1007/s10764-007-9177-x
Villareal, H., Álvarez, M., Córdoba, S., Escobar, F., Fagua, G., Gast, F., Mendoza, H., Ospina, M., & Umaña, A. M. (2004). Manual de métodos para el desarrollo de inventarios de biodiversidad. Instituto de investigación Alexander von Humboldt.
Wilcove, D. S., McLellan, C. H., & Dobson, A. P. (1986). Habitat fragmentation in the temperate zone. En M. E. Soulé (Ed.), Conservation Biology: The Science of Scarcity and Diversity (pp. 237-256). Sinauer Associates, Inc.
Yepes, A. P., & Villa, J. A. (2010). Sucesión vegetal luego de un proceso de restauración ecológica en un fragmento de bosque seco tropical (La Pintada, Antioquia). Revista Lasallista de Investigación, 7(2), 24-34.
Zotz, G., & Andrade, J. L. (2002). La ecología y la fisiología de las epífitas y hemiepífitas. En M. R. Guariguata & G.H. Kattan (Eds.), Ecología y conservación de Bosques neotropicales (pp. 271-296). Editorial Tecnológica de Costa Rica.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Recibido: 15 de junio de 2023; Aceptado: 27 de noviembre de 2023
Resumen
Monteleón es considerado un relicto de bosque subandino, y su biodiversidad está amenazada por distintos factores antropogénicos, lo que causa su fragmentación y la pérdida del hábitat de las especies que allí habitan. Con el objetivo de conocer su actual composición, estructura y diversidad florística, se establecieron tres parcelas de 0.1 ha en zonas con distintos grados de perturbación. En total se registraron 941 individuos y 116 especies, donde la parcela con perturbación baja presentó la mayor diversidad alfa. Según el análisis de la diversidad beta, entre las comunidades, los individuos de las especies que se pierden son en su mayoría sustituidos por individuos de otras especies, y un pequeño porcentaje se pierde sin sustitución. En conclusión, se registró un estado de afectación en el bosque debido a su heterogeneidad, que aun así se encuentra en un estado natural de recuperación.
Palabras clave:
abundancia, heterogeneidad, número efectivo de especies, perturbación, riqueza.Abstract
Monteleon is considered to be a sub-Andean forest remnant, and its biodiversity is threatened by various anthropogenic factors, leading to forest fragmentation and habitat loss for the species inhabiting it. In order to contribute to the knowledge of its composition, structure, and floristic diversity, three 0.1 ha plots were established in areas with different degrees of disturbance. A total of 941 individuals and 116 species were recorded, where with the plot with low disturbance exhibiting the highest richness and the highest alpha diversity (0D, 1D, and 2D). According to the beta diversity analysis, among the communities, the individuals of the species that are lost are mostly replaced by individuals of other species, and a small percentage is lost without replacement. In conclusion, a state of affectation is reported in the forest due to its heterogeneity, even though it is in a natural state of recovery.
Keywords:
abundance, heterogeneity, effective number of species, disturbance, richness.INTRODUCCIÓN
La fragmentación del paisaje es un fenómeno mundial (Harrison & Bruna, 1999) que puede entenderse como la transformación de un hábitat originalmente continuo en uno conformado por parches, generalmente pequeños y aislados entre sí por una matriz que difiere del hábitat original (Wilcove et al., 1986; Fahrig, 2003; Fahrig et al., 2019). Su efecto sobre la diversidad suele estar relacionado con el aislamiento de grupos biológicos incapaces de atravesar dicha matriz, lo que puede contribuir a las extinciones locales (Wilcove et al., 1986; Fahrig, 2003; Fahrig et al., 2019).
En Colombia, esta situación es alarmante, pues la fragmentación provoca el deterioro del hábitat (Fahrig, 2003), la disminución de las poblaciones (Donovan et al., 2002) y la reducción de la diversidad florística (Stevenson & Aldana, 2008) y la diversidad genética (Banhos et al., 2016). En el departamento de Caldas, Álvarez-Mejía et al. (2007) resaltaron que los ecosistemas naturales de la cuenca del río Chinchiná son afectados negativamente por la fragmentación.
El bosque Monteleón es uno de los relictos boscosos que bordean el sector norte de la ciudad de Manizales, Caldas (Duque-Escobar, 2015). Es considerado como un relicto de bosque subandino, y su biodiversidad está amenazada por la urbanización, la deforestación, la extracción de especies ornamentales, la contaminación y la expansión de la frontera agrícola (Restrepo de Fraume et al., 1990; Echeverri-González, 1993; Duque-Salazar et al., 2000; González-Plazas, 2009; POT, 2017; Gil-Fernández & Jiménez-Gómez, 2018), lo que ha promovido su fragmentación y la pérdida del hábitat.
A pesar de las presiones antrópicas que afectan estos bosques, aún son reservorios representativos de la biodiversidad de la región, y es necesario generar estrategias para protegerlos (Sánchez et al., 2004; Sanín et al., 2006; Álvarez-Mejía et al., 2007; Ríos-Málaver, 2007; 2008; Rojas-Morales et al., 2011; Castaño et al., 2010; Posada-Herrera, 2013; Sierra-Giraldo & Sanín, 2014; MacGregor-Fors & Zuria, 2019; Méndez-Urbano, 2022).
Así, se plantea analizar la composición, la estructura y la diversidad florística del bosque Monteleón como estrategia para la toma de decisiones orientadas a su conservación.
MATERIALES Y MÉTODOS
Área de estudio
El bosque Monteleón (Figura 1) es un bosque subandino (Rudas et al., 2007) ubicado en la Cordillera Central de los Andes colombianos, específicamente en el sector nororiental de la ciudad de Manizales (Departamento de Caldas). Este bosque presenta un rango de 2142-2165 m de altitud y una extensión de 64.36 ha (EEM, 2013).
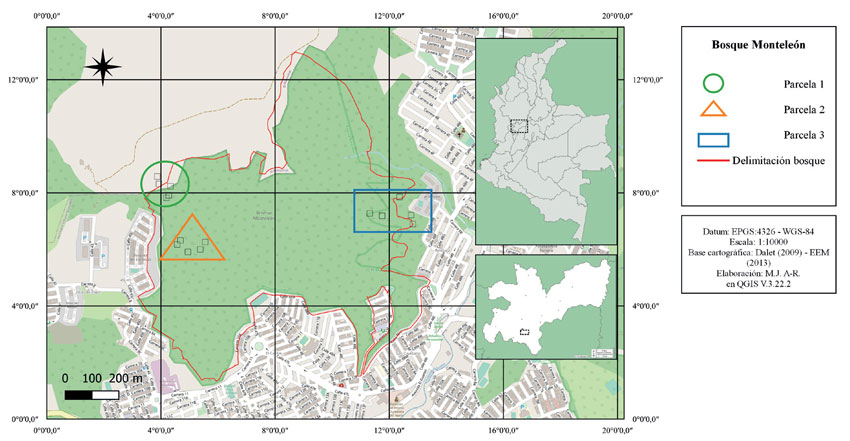
Figura 1: Mapa de localización del área de estudio del bosque Monteleón (delimitado en rojo), representando las parcelas (figuras geométricas) y los transectos (cuadrados dentro de cada parcela).
Se escogieron tres zonas con características ambientales y estructurales diferentes. La primera zona está ubicada en el noroccidente del bosque (05°04'50.7'' N, 75°30'00.4'' W y entre 2134 y 2163 m de altitud), limitando con el barrio Palonegro (Figura 1). Esta zona presenta una vegetación secundaria tardía, se encuentra influenciada por una quebrada y en ella predomina el epifitismo (Parcela 1). La segunda zona se encuentra en el centro-occidente del bosque (05°04'44.2'' N, 075°29'57.8'' W y entre 2134 y 2214 m de altitud) limitando con el barrio El Caribe (Figura 1). En esta zona, la vegetación es secundaria temprana, no presenta fuentes hídricas cercanas y es común la presencia de personas en la zona, las cuales realizan caminatas por los senderos (Parcela 2). Por último, la tercera zona se encuentra en el oriente del bosque (05°04'44.9'' N, 075°29'30.3'' W y entre 2131 y 2212 m de altitud), contigua al barrio Bengala (Figura 1). En esta zona, la vegetación es secundaria temprana y presenta varias señales de deforestación en algunos sectores, huertas urbanas y vestigios de deslizamientos pasados. Al igual que en la segunda zona, es común la presencia de personas transitando el bosque (Parcela 3). Con base en lo propuesto por Gómez-Ruiz et al. (2016), se determinó visualmente el grado de perturbación de cada una de las áreas, a partir del tipo de cobertura vegetal y sus agentes externos. De esta manera, se estableció que la primera zona cuenta con perturbación baja, la segunda con perturbación media y la tercera con perturbación alta.
Fase de campo y herbario
Se establecieron tres parcelas de 0.1 ha (Gentry, 1982), una en cada una de las tres zonas, distribuidas en cinco transectos de 50 x 4 m, para un total de 0.3 ha de muestreo.
En cada parcela se registraron los individuos de hábito arbóreo y lianas con un diámetro a la altura del pecho (DAP) mayor o igual que 2.5 cm. El DAP se midió a 1.3 m del suelo (Mendoza-Cifuentes, 2008). Se registró el valor de la circunferencia a la altura del pecho y la altura total de cada individuo, así como su determinación taxonómica. La recolección de los ejemplares botánicos se realizó de acuerdo con las metodologías propuestas por Forero (1977) y Alexiades (1996).
El procesamiento y la determinación taxonómica se llevaron a cabo en el Herbario de la Universidad de Caldas (FAUC), de acuerdo con los protocolos establecidos por Villareal et al. (2004). Para la confirmación de las especies, se revisó la colección del Herbario de la Universidad de Antioquia (HUA) y la colección virtual del Herbario Nacional Colombiano (COL) (ICN 2004), además de bibliografía especializada (e.g.Gentry, 1993; Mendoza & Ramírez, 2006; Lehnert, 2009; García-Ruenes & Alzate-Guarín, 2021). Los nombres científicos se validaron con la lista de referencia de plantas vasculares de Plants of the World Online (POWO, 2023).
Análisis de datos
Composición y estructura florística
Se analizó la estructura florística de cada una de las parcelas estudiadas en las tres zonas del bosque, así como la estructura de las tres parcelas en conjunto. Se obtuvieron datos de la composición florística, las clases diamétricas y el índice de valor de importancia (IVI). Además, las parcelas se compararon y se estimó su relación con base en sus variables estructurales. Para el cálculo del IVI, se determinó el área basal mediante la ecuación AB = π(DAP)2/ 4 (Franco-Rosselli et al., 1997). Para cada especie, se calculó la densidad relativa DeRel (número de individuos por especie/número total de individuos en la comunidad) x 100, la frecuencia relativa FreRel (número de parcelas en las que se encuentra la especie/número de parcelas totales) x 100 (Rangel-Ch. & Velásquez, 1997) y la dominancia relativa DoRel (ΣAB de todos los individuos de la especie/ΣAB de toda la comunidad). El IVI se calculó como la sumatoria de DeRel, FreRel y DoRel. Por último, para las variables estructurales de área basal (AB) y altura, se probaron los supuestos paramétricos, en aras de evaluar las diferencias entre las parcelas respecto a estas variables estructurales (p<0.05).
Diversidad alfa
La diversidad se midió en términos del número efectivo de especies qD (Hill, 1973; Jost, 2006; Cultid-Medina & Escobar, 2019), donde q = 0 corresponde a la riqueza de especies; q = 1 corresponde al número efectivo de especies igualmente comunes y es equivalente al exponencial de la entropía de Shannon; y q = 2 corresponde al número efectivo de especies dominantes y es equivalente al inverso del índice de Simpson (Jost, 2006, 2018). Esta aproximación permite medir la diversidad en términos de especies, lo cual facilita la interpretación en comparación con los índices tradicionalmente utilizados (Moreno et al., 2011; Jost, 2018). Según el protocolo de Chao y Jost (2012), se interpolaron las diversidades con respecto a la menor cobertura de muestreo y se graficaron los resultados como un perfil de diversidad para cada parcela. Estos índices de diversidad y sus intervalos de confianza (IC 95 %) (Cumming et al., 2007) fueron estimados con el paquete entropart (Marcon & Herault, 2015).
Con el fin de evaluar el porcentaje de diversidad representada en las muestras de cada una de las parcelas y realizar comparaciones entre estas, se calculó la cobertura de muestreo (Chao & Jost, 2012; Cultid-Medina & Escobar, 2019) mediante la siguiente ecuación:
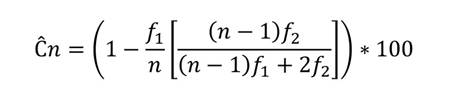
donde f 1 es el número de especies de las cuales solo se registró un individuo durante el muestreo (singletons), f 2 es el número de especies con dos individuos (doubletons) y n es el número total de individuos registrados. Si Ĉn ≈ 0, existe una mínima completitud, y si Ĉn ≈ 1, existe una máxima completitud.
Diversidad beta
Con el fin de determinar el grado de diferenciación entre las parcelas (diversidad beta), se construyó un dendrograma con los datos de las abundancias de las especies, calculado con la medida de disimilitud de Bray-Curtis (D BC) (Bray & Curtis, 1957) en el paquete betapart (Baselga & Orme, 2012). D BC se separó en dos componentes: los cambios balanceados de las abundancias (D BC-bal) y los gradientes de las abundancias (D BC-gra) (Baselga, 2013). Además, con el objetivo de determinar si existen o no agrupamientos de los conjuntos de datos -y, en caso de que existan, conocer cómo se agrupan-, se realizó un análisis de agrupamiento mediante el método del gap estadístico en el paquete cluster (Maechler et al., 2022). El análisis de agrupamiento se calculó considerando 100 conjuntos de datos aleatorios, que fueron simulados 100 veces para formar máximo dos clusters. Todos los análisis estadísticos se realizaron en el software estadístico R (R Core Team, 2022).
RESULTADOS
Composición y estructura florística
Se registraron 941 individuos y 116 especies. La parcela con perturbación baja presentó el mayor número de especies (70 especies), mientras que la parcela de perturbación media registró el mayor número de individuos (380 individuos) y la parcela de mayor perturbación registró el menor número de especies e individuos, 46 y 268 respectivamente (Tabla 1). Las especies con mayor IVI fueron Cecropia sp. (26.60) en la parcela menos perturbada y Hedyosmum goudotianum (49.77, 27.38) en las parcelas con perturbación media y alta (Figura 2). Respecto a las clases diamétricas, en las tres parcelas se observó una mayor concentración de individuos en las primeras clases (Figura 3).
Estructura florística de cada parcela, donde los valores del área basal y la altura son representados como media ± desviación estándar. Diversidad florística en cada parcela, donde Ĉn corresponde a la cobertura de muestreo observada.
q
D corresponde a la diversidad de Hill en cada parcela bajo una cobertura de muestreo del 88 %, donde el parámetro q corresponde a la sensibilidad del cálculo de las abundancias relativas de las especies (Jost, 2018). Las diversidades se representan como el número efectivo de especies, y los intervalos de confianza del 95 % se calculan a partir de 1000 simulaciones de bootstrap con la comunidad completa de cada parcela. Las letras como superíndices se refieren a las diferencias entre parcelas; diferentes letras entre dos parcelas significan diferencias estadísticamente significativas (p<0.05).
Tabla 1: Composición florística de cada parcela.
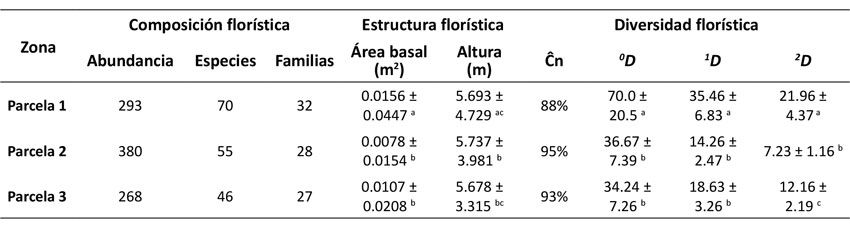
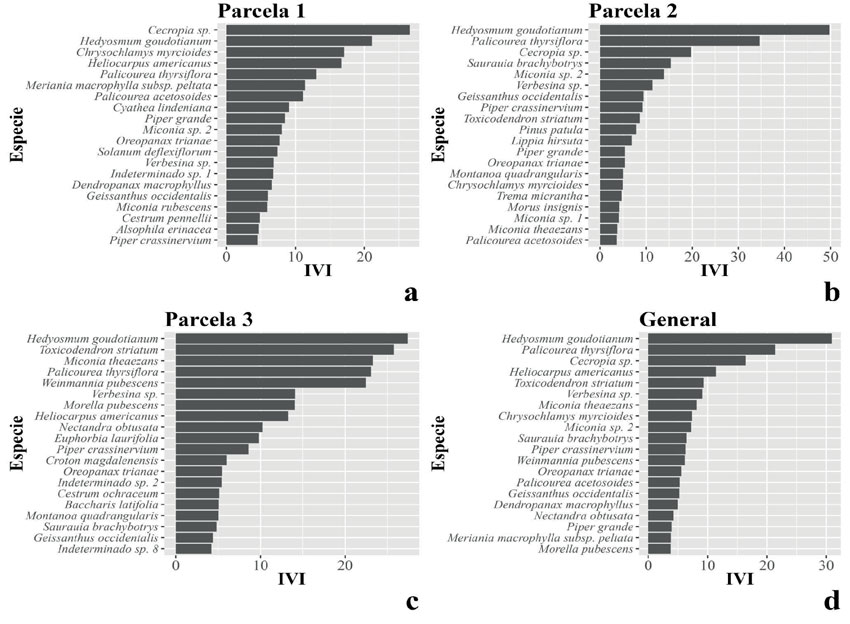
Figura 2: Las 20 especies con mayor índice de valor de importancia (IVI) en cada parcela y en general.
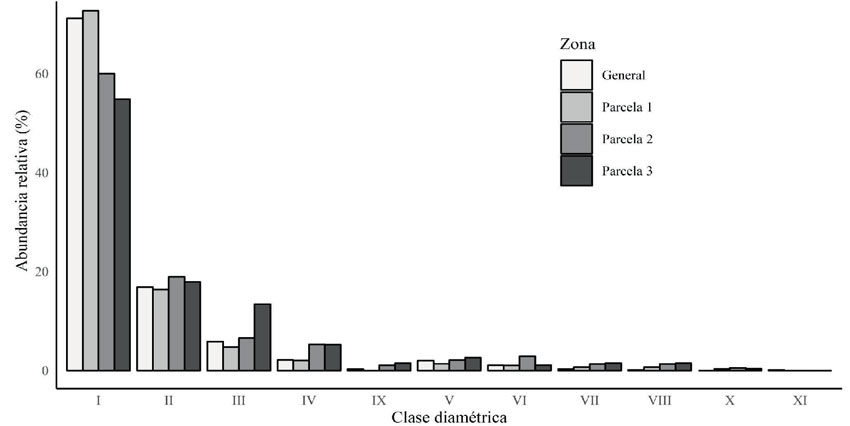
Figura 3: Abundancia relativa de las clases diamétricas en cada parcela y en general
Diversidad alfa
Se interpolaron las diversidades con respecto a la menor cobertura de muestreo (P1: Ĉn = 88%). Los valores de diversidad (qD) fueron estimados según cada orden (q = 0, 1, 2) con sus intervalos de confianza (qD ± IC 95 %) (Tabla 1). Los valores de la diversidad (número efectivo de especies) en la parcela 1 fueron 0D 70.0 ± 20.5, 1D 35.46 ± 6.83 y 2D 21.96 ± 4.37. En la parcela 2 fueron 0D 36.67 ± 7.39, 1D 14.26 ± 2.47 y 2D 7.23 ± 1.16. Finalmente, en la parcela 3 fueron 0D 34.24 ± 7.26, 1D 18.63 ± 3.26 y 2D 12.16 ± 2.19 (Figura 4a).
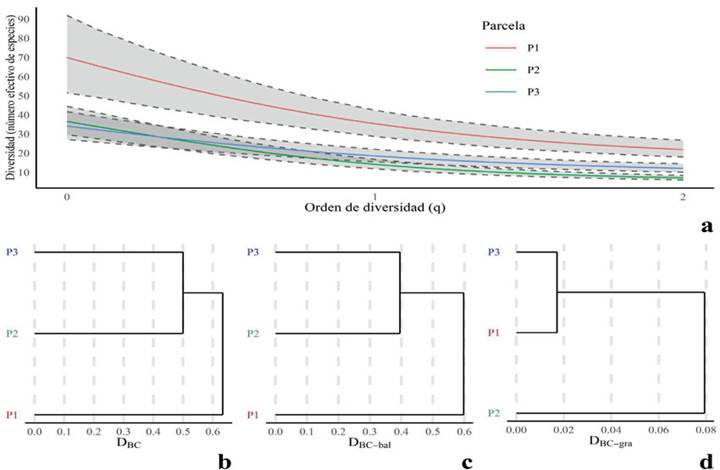
Figura 4. : Diversidad alfa (a) y diversidad beta (b-d).
Diversidad beta
Se realizó un análisis de la diversidad beta (Figuras 4b-d): la variación de las abundancias de las especies (Figura 4b, DBC) se particionó en la variación balanceada de las abundancias de las especies (Figura 4c, DBC-bal) y en los gradientes de las abundancias de las mismas (Figura 4d, DBC-gra). La distancia calculada por las medidas de disimilitud fue mínima entre las parcelas 2 y 3 para DBC y DBC-bal, y mínima entre las parcelas 1 y 3 para DBC-gra. En el análisis de agrupamientos con el método del gap estadístico, no se detectó formación de grupos con significancia estadística.
DISCUSIÓN
Composición y estructura florística
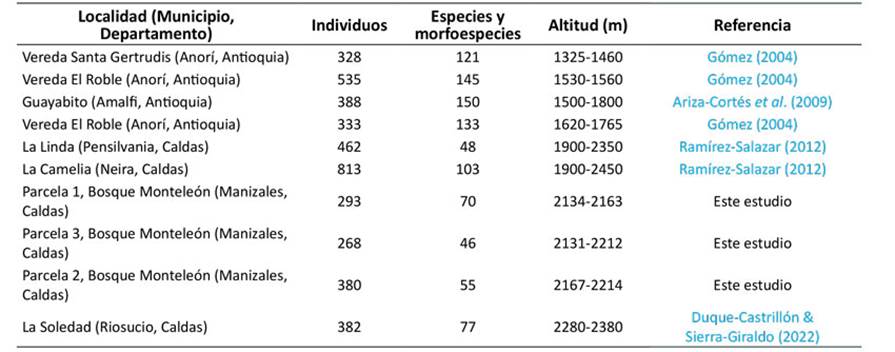
El número de individuos y especies de cada una de las parcelas (Tabla 1) fue similar a los resultados obtenidos en otros estudios de estructura florística en bosques subandinos en Colombia (Anexo 2). Al examinar las demás variables estructurales (Tabla 1), se encontró que la parcela 1 (perturbación baja) presentó una mayor área basal promedio, lo cual sugiere que esta zona está en un nivel de regeneración más avanzado (Fournier & Herrera, 1977). Aun así, las alturas en promedio fueron similares en las tres parcelas.
Respecto al IVI calculado, ninguna especie concentra más del 20 % del IVI total (Figura 2). Además, las cinco especies con mayor IVI en general (Hedyosmum goudotianum, Palicourea thyrsiflora, Cecropia sp., Heliocarpus americanus y Toxicodendron striatum) concentran el 29.8 % del IVI total, un porcentaje similar a los hallados en otros estudios (Dueñas et al., 2007). Asimismo, Orozco-León y Ocampo-Calderón (2010) registraron que las especies con el mayor IVI en Monteleón fueron son Dendropanax sp., Piper crassinervium, Toxicodendron striatum y Cecropia telealba, todas heliófitas durables, indicando que no existe aún un estado de madurez en el bosque.
En cada una de las parcelas, y de manera general, más del 50 % de los individuos se concentraron en la primera clase diamétrica (Figura 3), un aspecto que también se ha observado en otros estudios en el bosque Monteleón (Orozco-León & Ocampo-Calderón, 2010) y en otros bosques del país (Mosquera et al., 2007; Yepes & Villa, 2010). Este comportamiento sugiere que el bosque está en un proceso de recuperación (Yepes & Villa, 2010; Martínez-Calderón et al., 2021). En todas las parcelas se registra más del 60 % de las especies en esta primera clase diamétrica, lo cual refleja una alta heterogeneidad de especies que aportan al proceso de recuperación del bosque, dominado por las heliófitas durables (Sánchez et al., 2007). Esto indica que el bosque presenta una capacidad natural de regeneración y de sucesión ecológica (Zotz & Andrade, 2002) para alcanzar un futuro estado de madurez (Finegan, 1992). En este sentido, Monteleón requiere de atención para su manejo y conservación (Guariguata & Ostertag, 2002). Se espera que este estudio sirva como base para el adecuado manejo y la toma de decisiones en función de su protección (Silva-Aparicio et al., 2018; Martínez-Calderón et al., 2021, Ángeles-García et al., 2022), ya que, al conocer la composición, distribución y diversidad de especies de un área de interés, se pueden tomar acciones específicas que permitan la valoración ambiental, un mejor uso y un aprovechamiento de los recursos naturales, aspectos que apoyan la construcción de un adecuado plan de manejo para la conservación (Dirzo & Raven, 1994; Sánchez- Cordero et al., 2001).
Diversidad alfa
En las tres parcelas, las coberturas de muestreo (Ĉn) fueron similares entre sí y mayores al 80 % (Cultid-Medina & Escobar, 2019), pues se obtuvieron valores cercanos al 90 % (Ĉn 1 =88 %, Ĉn 2 =95 %, Ĉn 3 =93 %). Respecto a la riqueza (0D), se estiman 70 ± 20 especies en la parcela 1, 37 ± 8 especies en la parcela 2 y 35 ± 8 especies en la parcela 3, es decir, la parcela 1 presenta aproximadamente el doble de riqueza que las parcelas 2 y 3. Respecto al número de especies igualmente comunes (1D), en las parcelas 1, 2 y 3 se estiman 36 ± 7, 15 ± 3 y 19 ± 4 especies respectivamente (Jost & González-Oreja, 2012), indicando que la parcela 1 puede ser aproximadamente el doble de diversa que la 2 y la 3. Respecto al número efectivo de especies dominantes (2D), en las parcelas se estiman 22 ± 5, 8 ± 2 y 13 ± 3 especies respectivamente, que corresponden a las especies con las mayores abundancias. Así, se deduce que, según esta diversidad (2D), la comunidad con mayor dominancia es la de la parcela 2, y la comunidad con menor dominancia es la de la parcela 1. Este análisis refleja la amplia diferencia de diversidad que existe entre las parcelas, en especial en relación con la parcela 1, la más diversa de todas.
La diversidad de la parcela 1 (perturbación baja) es significativamente mayor que en las demás parcelas (Figura 4a), lo que puede deberse a la presencia de una quebrada como fuente de oferta hídrica (García-Martínez & Mercado-Gómez, 2017). Esta quebrada puede representar hábitats adecuados para especies de animales, los cuales podrían promover la dispersión de semillas en esta área menos perturbada. Este comportamiento, donde la zona con perturbación baja presenta una mayor diversidad alfa, se ha registrado en otros estudios (Gómez-Ruiz et al., 2016; Orozco-Agudelo & Sierra-Giraldo, 2022), lo que refleja un riesgo significativo de pérdida de especies de plantas para el bosque Monteleón, ya que la diversidad (0D, 1D y 2D) fue mayor en la parcela 1, la cual, a pesar de presentar la menor perturbación, está más expuesta a la fragmentación, pues tiene diferentes proyectos urbanísticos a su alrededor (Alcaldía de Manizales, 2018).
En la Figura 4a se observa un descenso significativo del número efectivo de especies a medida que aumenta q, lo que podría sugerir un alto grado de dominancia en las comunidades (Jost, 2006). Lo anterior se puede complementar con lo encontrado en la distribución de las clases diamétricas (Figura 3): el bosque se encuentra en un proceso de recuperación, y este estado lo hace más sensible a perturbaciones antropogénicas, pues las plantas de menor porte y grosor son las más frágiles ante acciones de deforestación, contaminación y senderismo, además de eventos naturales de deslizamiento de tierra, que no son raros en Monteleón (El Tiempo, 1993).
Diversidad beta
Según el método del gap estadístico no se encontraron agrupamientos, es decir, no se pueden agrupar las terminales del dendrograma entre sí (Baselga, 2013). Este análisis sugiere que el recambio de las abundancias de las especies (D BC-bal) representa mejor la variación de las abundancias de las comunidades (diversidad beta) que los gradientes de las abundancias entre las comunidades (D BC-bal). Así, entre las comunidades, los individuos de las especies que se pierden son en su mayoría sustituidos por individuos de otras especies, y un pequeño porcentaje de individuos se pierde sin sustitución. Este aspecto refleja en gran medida la resiliencia del bosque y su proceso de sucesión, pero también indica que algunos individuos de ciertas especies se están perdiendo sin ser reemplazados. Esto puede estar relacionado con las actividades de extracción de plantas ornamentales (Echeverri, 1993) y con la tala selectiva de especies arbóreas que se han observado en el área de estudio (Restrepo de Fraume et al., 1990, Echeverri, 1993; Duque-Salazar et al., 2000). No obstante, entre las parcelas, las comunidades fueron disímiles, principalmente debido a las variaciones balanceadas de las abundancias. Si se tiene en cuenta solo el gradiente de las abundancias, la disimilitud fue mayor entre las parcelas 1 y 2 que entre la 1 y la 3. Esto quiere decir que los individuos de las especies que se están perdiendo entre las parcelas 1 y 2 están siendo menos reemplazados que los individuos de las especies que se están perdiendo entre la 1 y la 3. Así, puede que la extracción de plantas ornamentales y la tala selectiva de árboles sea mayor en la zona de las parcelas 1 y 2 que en la de la 2 y la 3, si bien haría falta más evidencia para esta deducción. En caso de ser verdadera, esta información permitiría a las autoridades ambientales conocer cómo se están distribuyendo las afectaciones antropogénicas en el bosque, a fin de promover su conservación.
CONCLUSIONES
La parcela con perturbación baja fue la que presentó mayor área basal promedio, mayor riqueza (0D), mayor número de especies igualmente comunes (1D) y mayor número de especies dominantes (2D), es decir, la parcela con menor perturbación presentó la mayor diversidad alfa. Según el análisis de la diversidad beta, se puede deducir que, aunque entre las comunidades los individuos de las especies que se pierden son en su mayoría sustituidos por individuos de otras especies, un pequeño porcentaje de individuos se está perdiendo sin ser sustituido, probablemente debido a la extracción de plantas ornamentales y la tala selectiva, que podría ser mayor en la zona de las parcelas 1 y 2.
Monteleón está en un proceso de recuperación natural, lo cual hace urgente desarrollar un plan de manejo efectivo para cada una de sus coberturas vegetales, priorizando su conectividad con otros bosques (e.g. Los Yarumos, El Águila y Río Blanco). Esto, sumado con la prohibición de la extracción de plantas ornamentales y la tala selectiva, el desarrollo de estrategias de educación y adecuación del bosque para la comunidad y procesos de restauración ecológica acompañados de monitoreos.
Acknowledgements
AGRADECIMIENTOS
A la Vicerrectoría de Investigaciones y Posgrados de la Universidad de Caldas por financiar este proyecto (código 0126121). A los integrantes del Semillero de Investigación en Plantas y Afines Phytos por su apoyo, especialmente a Brandon Gallego, Daniel Giraldo, Diana Puetate, Johana Zárate y César Duque. A Laura Cuervo por complementar las ideas sobre fragmentación ecológica. A los directores y curadores de los herbarios FAUC y HUA por permitirnos revisar sus colecciones. A los diferentes especialistas que apoyaron y confirmaron la determinación taxonómica, a saber: Jáider Jiménez (Araliaceae), Angy Caro (Clusiaceae), Ana María Trujillo (Fabaceae), Camilo Sánchez (Lauraceae), Mauricio Posada y Steven Murillo (Melastomataceae), Ricardo Callejas (Piperaceae), Heriberto David (Rubiaceae), Juan David Tovar (Solanaceae), Andrés Felipe Bohórquez y Santiago Guzmán (varias familias). A Jairo Orozco por su apoyo en el establecimiento de parcelas y el prensado de muestras. A Giovanny Blandón por compartirnos emotivamente la situación socioambiental del bosque. A Jorge Mario Herrera por su asesoría en la sección de diversidad florística y la revisión del manuscrito. Por último, queremos agradecer a los dos revisores anónimos por sus útiles comentarios que enriquecieron el manuscrito
REFERENCIA
ANEXOS
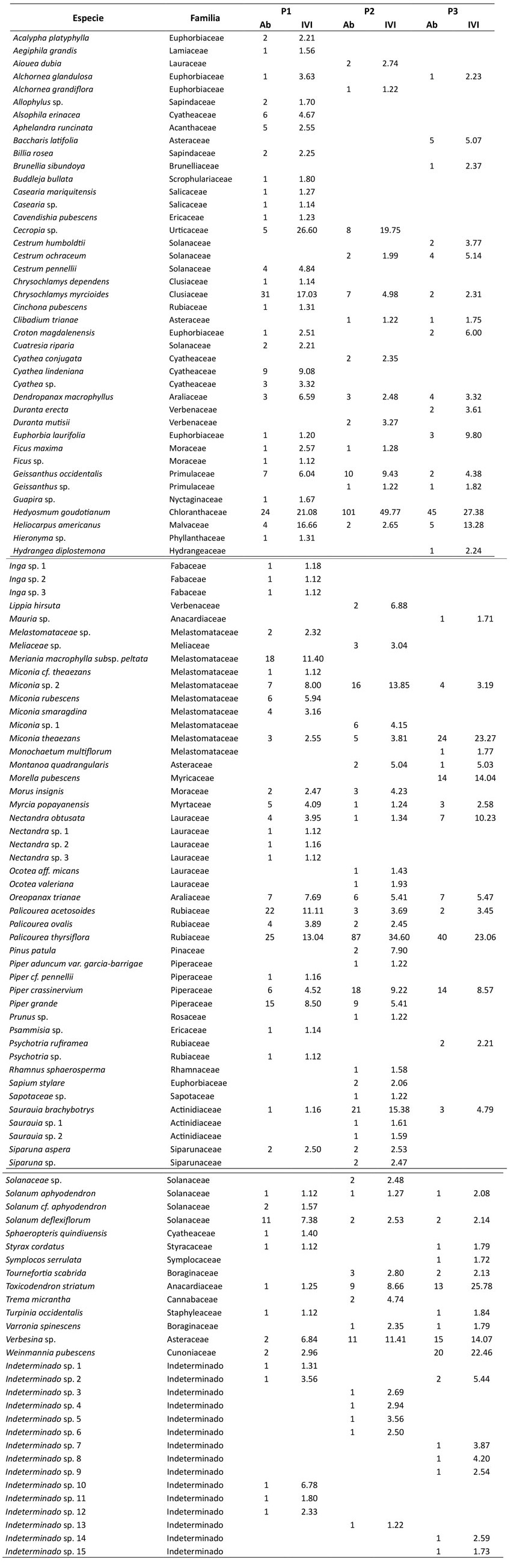
Anexo 1: Especies de plantas incluidas en este estudio. Para cada especie, se presenta la abundancia (Ab) y el índice de valor de importancia (IVI) en la parcela 1 (P1), la parcela 2 (P2) y la parcela 3 (P3).
Anexo 2: Estructura florística de algunos bosques subandinos de Colombia* a partir de datos de parcelas de 0.1 ha con individuos con DAP mayor o igual a 2.5 cm ordenados según la altitud promedio. *Clasificación de bosque subandino según Rudas et al. (2007).




Licencia
Derechos de autor 2024 Colombia forestal

Esta obra está bajo una licencia internacional Creative Commons Atribución-CompartirIgual 4.0.
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






