
DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2007.1.a03Publicado:
01-01-2007Número:
Vol. 10 Núm. 20 (2007): Enero-DiciembreSección:
Artículos de investigación científica y tecnológicaEstructura y composición florística de la vegetación del corredor biológico entre los parques nacionales naturales Puracé y Cueva de los Guácharos
Structure and floristic composition of the vegetation of the biological corridor between the national natural parks Puracé and Cueva de los Guácharos
Palabras clave:
Sucesión vegetal, diversidad, estructura de la vegetación, bosque andino, bosque secundario, corredor biológico, Huila, Colombia (es).Palabras clave:
Plant succession, diversity, vegetation struc- ture, andean forest, secondary forest, biological corridor, Huila, Colombia. (en).Descargas
Referencias
Bello, A. & Kattib. 1994. Aspectos ecológicos en la sucesión secundaria de la vegetación subandina (2010 2510 m) en localidades del municipio de Santa Rosa de Cabal. Risaralda, Colombia. Trabajo de grado. Departamento de Biología. Universidad Nacional. Bogotá.
Bernal, A. 1973. Estudio de la vegetación y planificación preliminar del Parque Nacional Natural Cueva de los Guácharos, Acevedo, Huila. Bogotá D.C., Colombia. Trabajo de grado (Ingeniero Forestal). Universidad Distrital Francisco José de Caldas.
CCAD-PNUD/GEF. 2002. Corredor biológico mesoamericano, una plataforma para el desarrollo sostenible regional. Serie técnica No. 1. Proyecto para la consolidación del corredor biológico mesoamericano. Managua, Nicaragua. 24 p.
Cantillo, E., K. Rodríguez & A. Avella. 2005. Caracterización florística, estructural, diversidad y ordenación de la vegetación arbórea en la reserva forestal Cárpatos (Guasca Cundinamarca) Universidad Distrital "Francisco José de Caldas" Ingeniería Forestal.
Cleef, A. 1972. Informe preliminar sobre la botánica del Parque Nacional Natural Cueva de los Guácharos. Bogotá. Colombia.
Coca, C. & Y. González. 2007. Caracterización florística y estructural de la vegetación del corredor biológico entre los Parques Nacionales Naturales Puracé Cueva de los Guácharos. Trabajo de grado (Ingeniero Forestal). Universidad Distrital Francisco José de Caldas. Bogotá D.C., Colombia. 282 p.
González Y., C. Coca & G. Téllez. 2008. Catálogo de la flora vascular de los bosques andinos y subandinos del corredor biológico entre los Parques Nacionales Naturales Puracé - Cueva de los Guácharos. Universidad Distrital Francisco José de Caldas.
Cuatrecasas, J. 1958. Aspectos de la vegetación natural de Colombia. Rev. Acad. Colomb. Cienc. 10(40): 221-268.
Duque-N., A. 1987. Comunidades vegetales de la zona paramuna del PNN Puracé (Colombia). Tesis de grado, Universidad del Valle. (Mimeografiado).
Duque-N., A. & J.O Rangel-Ch. 1989. Análisis fitosociológico de la vegetación paramuna del PNN Puracé. En: L. F. Herrera, R. Drenan y C. Uribe (eds). Cazicazgos prehispánicos del Valle de la Plata. Tomo I. El contexto medioambiental de la ocupación humana. U. of Pittsburg memoirs in Latin América-Archaeology. 2:70-95.
Holdridge, L. R. 1979. Ecología basada en zonas de vida. Costa Rica. Publicaciones del IICA.
Lozano, G., J. Hernández & J. Henao 1979. El género Trigonobalanus en el Neotrópico. I. Caldasia 12: 517 537.
Mahecha, G. E. 1999. Estudio florístico de la región del macizo colombiano. Principios y procedimientos para el conocimiento de las plantas superiores del trópico americano. Convenio Intercorporativo del Macizo Colombiano. Huila, Colombia.
Rangel-Ch., J. O. & P. Franco R. 1985. Observaciones fitoecológicas en varias regiones de vida de la cordillera central de Colombia. Caldasia 14 (67): 210-249.
Rangel-Ch., J. O. & G. Lozano 1986. Un perfil de vegetación entre la Plata Huila y el volcán del Puracé. Caldasia 14 (68-70) 503 547.
Rangel-Ch., J. O., A. M. Cleef, T. Van Der Hammen & R. Jaramillo 1982. Tipos de vegetación en el transecto Buritaca La Cumbre Sierra Nevada de Santa Marta (entre 0 y 4100 msnm). Colombia geográfica 10(1): 1-18.
Rangel-Ch., J. O., A. M. Cleef & S. Salamanca 2003. The ecuatorial interandean and subandean forest of the Parque los Nevados Transect, Cordillera Central, Colombia. 143-204 p. En: T. van der Hammen & A. Dos Santos (eds). La Cordillera Central Colombiana. Estudio de Ecosistemas Tropandinos 5. J. Cramer, Berlín-Stuttgart.
Rojas, O. 1984. Evolución de una sucesión vegetal en el Valle del Cauca. Cespedesia 13 (49 50) 152-211.
Sánchez, J. 2003. Aves del corredor biológico entre los Parques Nacionales Naturales Puracé Cueva de los Guácharos. Guía de Campo. 358 p.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Colombia Forestal,Diciembre del 2007 vol:10 nro:20 pág:40-78
Estructura y composición florística de la vegetación del corredor biológico entre los parques nacionales naturales Puracé y cueva de los Guácharos 1
Yitsully González-O
Ingeniera Forestal. Asistente de investigación, Universidad Distrital "Francisco José de Caldas". E-mail: yitsully@gmail.com
Ana Coca-A
3 Ingeniera Forestal. Asistente de investigación, Universidad Distrital "Francisco José de Caldas". E-mail: anitakro07@hotmail.com
Edgard Ernesto Cantillo-H
4 Profesor de Ecología. Proyecto curricular de Ingeniería Forestal. Universidad Distrital "Francisco José de Caldas". E-mail: ecantillo@udistrital.edu.co
Resumen
La caracterización florística y estructural de la vegetación intervenida y no intervenida del corredor biológico entre los Parques Nacionales Naturales (PNN) Puracé y Cueva de los Guácharos se realizó con base en 16 levantamientos ubicados en tres municipios del sur del Huila, Colombia entre los 1950 y 2450 m; En total se muestrearon 1.5 ha.
Se definió la alianza Cyatheo Cecropion angustifoliae que incluye las asociaciones Ladenbergio macrocarpae Elaeagietum myrianthae y Guettardo hirsutae Hedyosmetum translucidi. Para los sitios de menor intervención se encontraron las comunidades de Helicostylis tovarensis Alfaroa williamsii, de Quercus humboldtii Wettinia fascicularis y de Weinmannia pubescens Clusia dixonii. En las áreas de mayor intervención se establece vegetación de la comunidad de Baccharis nitida y Saurauia pulchra.
Para las asociaciones los valores de área basal por especie fueron similares. Para las comunidades, se presentaron los valores más altos con 7.3 y 4.6 m², en la comunidad de Quercus humboldtii y Wettinia fascicularis. El promedio de alturas obtenido para las asociaciones fue de 13 m, con valores de 10 y 15 m. Para las comunidades, el promedio obtenido fue de 13m, con variaciones entre 7 y 17 m. El estrato dominante en ambos casos fue el arbóreo inferior.
Los índices de importancia muestran que para todas las unidades a excepción de la Comunidad de Quercus humboldtii y Wettinia fascicularis, existe una representatividad equitativa de las especies dentro de cada unidad. Las familias mejor representadas en número de especies son Lauraceae, Rubiaceae y Melastomataceae.
Para las asociaciones los valores de área basal por especie fueron similares. Para las comunidades, se presentaron los valores más altos con 7.34 y 4.6 m², en la comunidad de Quercus humboldtii y Wettinia fascicularis. El promedio de alturas obtenido para las asociaciones fue de 13 m, con valores de 10 y 15 m. Para las comunidades, el promedio obtenido fue de 13m, con variaciones entre 7 y 17m. El estrato dominante en ambos casos fue el arbóreo inferior.
Los índices de importancia muestran que para todas las unidades a excepción de la Comunidad de Quercus humboldtii y Wettinia fascicularis, existe una representatividad equitativa de las especies dentro de cada unidad. Las familias mejor representadas en número de especies son Lauraceae, Rubiaceae y Melastomataceae.
Palabras clave: Sucesión vegetal, diversidad, estructura de la vegetación, bosque andino, bosque secundario, corredor biológico, Huila, Colombia.
Abstract
The floristic composition and structure of disturbed and non disturbed vegetation at the biological corridor located among the National Natural Parks Puracé, Cueva de los Guácharos was studied based on 16 plots in three localities of the Huila Department, Colombia, between 1950 and 2450 m. A total of 1.5 ha was sampled.
The Cyatheo Cecropion angustifoliae alliance was defined. It includes the associations Ladenbergio macrocarpae Elaeagietum myrianthae and Guettardo hirsutae Hedyosmetum translucidi. At the less disturbed areas the communities Helicostylis tovarensis Alfaroa williamsii, Quercus humboldtii Wettinia fascicularis and Weinmannia pubescens Clusia dixonii were found. The community Baccharis nitida and Saurauia pulchra was found in the most disturbed areas.
The basal area value per species was similar for all the associations. The community Quercus humboldtii and Wettinia fascicularis showed the highest basal area value, 7.3 and 4.6 m². Regarding forest tall, an average of 11 m was found in the associations, with values from 10 to 15 m. An average of 13 m was found in the communities, with variations from 7 to 17 m. The dominant stratum in both cases was the arboreal inferior.
The importance indexes show an equal representativeness of the species inside each unit, with the exception of the Quercus humboldtii and Wettinia fascicularis community. The best represented families regarding their species number are Lauraceae, Rubiaceae and Melastomataceae.
Key words: Plant succession, diversity, vegetation structure, andean forest, secondary forest, biological corridor, Huila, Colombia.
INTRODUCCIÓN
Los corredores biológicos definidos como mecanismo para la conservación, son áreas que a través de su ordenamiento permiten la conexión entre dos o más áreas protegidas con el fin de lograr el intercambio genético y mantener o aumentar la riqueza y diversidad (modificado de CCAD-PNUD/GEF, 2002).
Dependiendo de las características ecológicas y sociales del área, en los corredores biológicos se pueden llevar a cabo actividades netamente protectoras o de producción sostenible. En el caso del corredor biológico entre los PNN Puracé, Cueva de los Guácharos, se han destinado algunas áreas para la producción mediante arreglos forestales, mientras que otras áreas sirven para la protección de los recursos naturales.
El estudio se enmarcó dentro del componente de reforestación y protección al medio ambiente; de esta manera se realizó una caracterización florística y estructural de la vegetación de las áreas boscosas y de las zonas intervenidas que integran el corredor biológico con el fin de conocer las especies, su arreglo espacial, su diversidad y así determinar el manejo apropiado de acuerdo a sus posibilidades de uso.
De acuerdo con las observaciones realizadas, la conservación de los bosques depende del grado de accesibilidad, su cercanía a las viviendas y en el caso de áreas privadas del compromiso de protección por parte de los propietarios. La mayoría de los bosques bien conservados se encontraron en sitios con pendientes muy escarpadas y a grandes distancias, en muchos casos se observaron áreas que habían sido aprovechadas intensamente. En la actualidad se presentan aprovechamientos sectorizados para fines domésticos que han sido permitidos por la Corporación Autónoma Regional del Alto Magdalena (CAM).
Aunque la zona de estudio fue catalogada como una de las más diversas del macizo colombiano al ser declarada como reserva de la biosfera en 1979, en la actualidad son pocos los estudios realizados. Entre las investigaciones que se conocen se destacan las de Cleef (1972), Lozano (1979) y Mahecha (1999) que fueron elaboradas para contribuir al conocimiento de la flora de los bosques mediante la elaboración de catálogos. Igualmente los trabajos ecológicos son reducidos, se encuentra el de Bernal (1976), quien realizó un estudio donde presenta la composición florística y estructural de los bosques del municipio de Acevedo; Duque (1987), quien elaboró un estudio de las comunidades vegetales del PNN Puracé; Rangel & Franco (1985), con los registros y observaciones fitoecológicas en dos transectos realizados desde la Plata (Huila) hasta el Puracé, que luego sería complementada con la caracterización de la vegetación realizada por Rangel & Lozano (1986) a lo largo de un transecto desde La Plata (Huila) hasta el Volcán del Puracé. Duque & Rangel (1989), realizaron un análisis fitosociológico de la vegetación del páramo del PNN Puracé.
En la región Andina se han realizado diferentes estudios ecológicos como las contribuciones de Rangel (1995), quien efectuó un estudio de diversidad de las familias, géneros y especies de la vegetación en el Parque los Nevados; Rangel & Garzón (1994), con la caracterización estructural, dinámica y diversidad de la vegetación en el Parque Regional Natural Ucumarí; Rangel et al. (1982) quienes hicieron una descripción de las regiones a lo largo de un transecto realizado en el Tayrona y Pozos Colorados entre 0 y 4100 msnm y Rangel et al. (2003) con la clasificación sintaxonómica de los bosques subandinos del transecto Parque de los Nevados. Entre otros estudios acerca de los cambios de la vegetación se encuentran el de Rojas (1984), quien registró observaciones sobre la dinámica sucesional durante ocho años en áreas que habían sido destinadas a la ganadería extensiva y Bello & Kattib (1994), que determinaron el proceso de sucesión secundaria en la franja subandina después de la quema de algunas de las unidades de vegetación.
ÁREA DE ESTUDIO
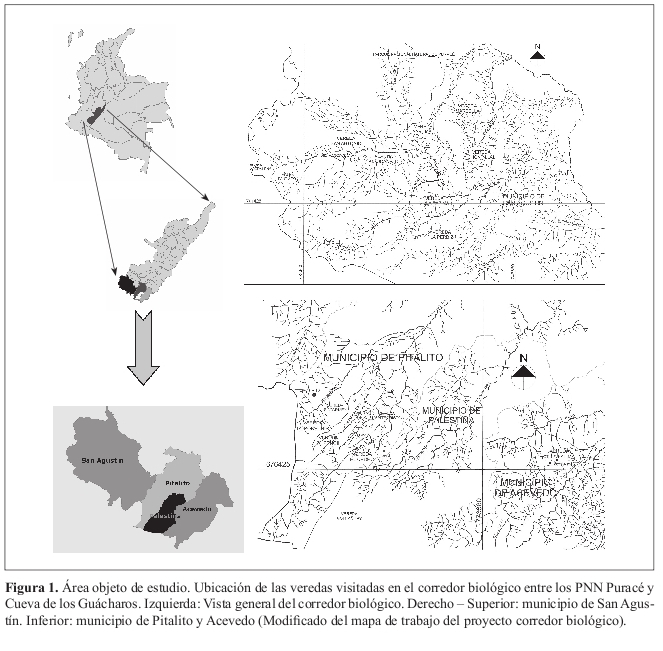
El corredor biológico entre los PNN Puracé y Cueva de los Guácharos se encuentra ubicado entre los 5º 57' 19'' a 6º 38' 17.6'' N y 72º 14' 38'' a 73º 2' 20'' W, al sur del departamento del Huila, Colombia, en una superficie calculada de 84465 ha (Figura 1). Comprende áreas de los municipios de San Agustín, Pitalito, Acevedo y Palestina (Sánchez 2003). Los levantamientos se ubicaron entre los 1955 y 2443 msnm.
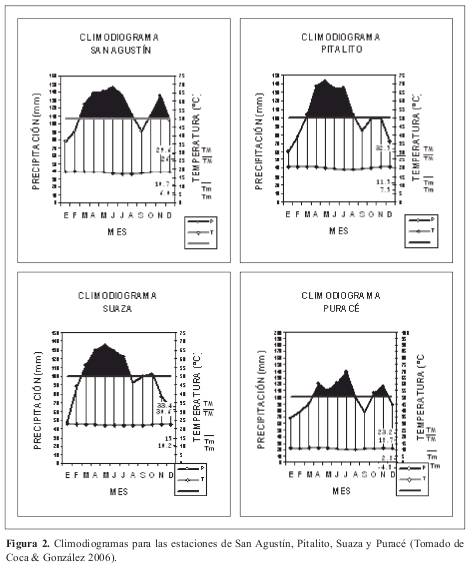
De acuerdo con las estaciones de San Agustín, Pitalito, Suaza y Puracé, se presenta a lo largo del corredor un régimen de precipitación bimodal tetraestacional (Figura 2) con valores promedio anuales de 1201,9 mm en la estación de Suaza a 1402,3 mm en la estación de San Agustín. El pico de mayor precipitación se encuentra en mayo para las estaciones de Pitalito y Suaza y junio en la estación de San Agustín.
Se presenta un régimen isotérmico con variaciones promedio de 1.5 ºC a lo largo del año. En las estaciones, los valores medios de temperatura oscilan entre los 11.06 ºC y 22.42ºC, en San Agustín y Acevedo respectivamente. Los promedios mínimos de temperatura se encuentran entre 2.5ºC y 11.6ºC, en las mismas estaciones. En González et al. (2008) se encuentra mayor información climática de la región.
El área del corredor biológico según la clasificación de Cuatrecasas (1958), pertenece a la franja de Selva Subandina y Andina. De acuerdo con la clasificación climática de Holdridge (1987), las estaciones de Pitalito, San Agustín y Suaza pertenecen a la franja de bosque húmedo premontano (bh-PM), mientras que la estación Puracé se define como bosque húmedo montano bajo (bh-MB). De acuerdo a Fernández (1989), el material parental predominante de la región de Acevedo es proveniente de rocas ígneas, principalmente granitos moscovíticos y andesitas ortósicas, además de rocas sedimentarias como areniscas, arcillas rojas y grises, conglomerados y aglomerados. El material parental en el municipio de Pitalito, está compuesto principalmente por rocas sedimentarias y una gruesa capa de arenisca roja, sobre la cual descansa una capa de arcilla. Los suelos del corredor biológico poseen texturas desde Francas hasta Areno Francosas, pH promedios de 3.8 a 5.3, bajos contenidos de fósforo (0-3.79 ppm), contenidos de materia orgánica entre 1.5% y 5.3% y CIC entre 22.6 y 37.2 meq/100g.
Los levantamientos realizados en el municipio de San Agustín, muestran que la textura predominante es Areno-Francosa, pH entre 4.5 y 5, contenidos de fósforo de 3.7 ppm en promedio, porcentaje de materia orgánica de 5.2% en los primeros 30cm y 3.2% en los 60 cm, y una CIC de 30 a 38 meq/100g. En el municipio de Pitalito, los suelos poseen una textura Franco-Arenosa, aunque también se minante es Areno-Francosa, pH entre 4.5 y 5, contenidos de fósforo de 3.7 ppm en promedio, porcentaje de materia orgánica de 5.2% en los primeros 30cm y 3.2% en los 60 cm, y una CIC de 30 a 38 meq/100g.
En el municipio de Pitalito, los suelos poseen una textura Franco-Arenosa, aunque también se encontraron texturas Francas y Franco Arcillo Arenosas, pH entre 5 y 5.3, valores de fósforo entre 2 y 3.6, contenidos de materia orgánica entre 3.5% en los primeros 30cm y 1.7% a 60cm. Los valores de CIC son muy uniformes hasta los 60cm con valores de 22 a 24 meq/100g.
En el municipio de Acevedo, se encontró que la textura es Franco Arenosa, pH muy ácidos entre 3.8 a 4.6, valores muy bajos de fósforo, desde nulos hasta 0.65 ppm en promedio, porcentajes de materia orgánica entre 5.3% en los primeros 30 cm y 2.5% a 60 cm. Los valores promedios de CIC se encuentran entre 29.6 y 33.5 meq/100g.
MATERIALES Y MÉTODOS
Los sitios a muestrear se determinaron con la ayuda de imágenes de satélite de propiedad de la Corporación Autónoma Regional del Alto Magdalena (CAM). Fueron escogidos los bosques con el mejor estado de conservación y de mayor extensión relativa en cada uno de los municipios. Las veredas visitadas se escogieron de acuerdo al grado de accesibilidad de cada una. Para el análisis climático se calcularon los índices termopluviométricos de Caldas & Lang, climodiogramas y balances hídricos. El mapa de Isohietas se elaboró con todas las estaciones cercanas al área de estudio y con ayuda del programa Surfer®.
Se elaboraron parcelas de 1000 m² (10 x 100 m). En cada una se estructuraron cuatro subparcelas de 5 x 5 m para obtener la información referente a latizales (2.5 cm<DAP <10cm y con alturas 1.5 m) a su vez dentro de cada una se realizó una subparcela de 2 x 2m para el conteo de brinzales (individuos con alturas < 1.5 m).
Para la toma de información en las áreas intervenidas se diseñaron levantamientos desde 500 m² hasta 1000 m² dependiendo de la extensión de los bosques. Las subparcelas para latizales y brinzales se realizaron a manera de fajas a lo largo de la parcela, cada 25 m. Para los latizales se establecieron cuatro fajas de 10 x 5m y para los brinzales otras cuatro de 10 x 2 m, de esta manera se obtuvo información adicional sobre las especies arbustivas, las cuales son representativas de sitios intervenidos.
Los datos tomados en campo para cada árbol mayor a 10 cm de DAP (fustales) y latizales fueron: nombre vulgar, número consecutivo y de colección, ubicación en coordenadas (x, y) dentro de la parcela, altura total, altura comercial, CAP (Circunferencia a la altura del pecho) diámetros de copa, así como defectos estructurales y fitosanitarios de cada árbol. Para los brinzales se consignó en los registros su abundancia por especie. Igualmente se tomaron coordenadas, altitud, pendiente y azimuth de cada levantamiento realizado.
La fase de colección, tratamiento e identificación de las muestras se encuentra explicado ampliamente en González et al. (2008) donde se presenta el catálogo general de las especies identificadas.
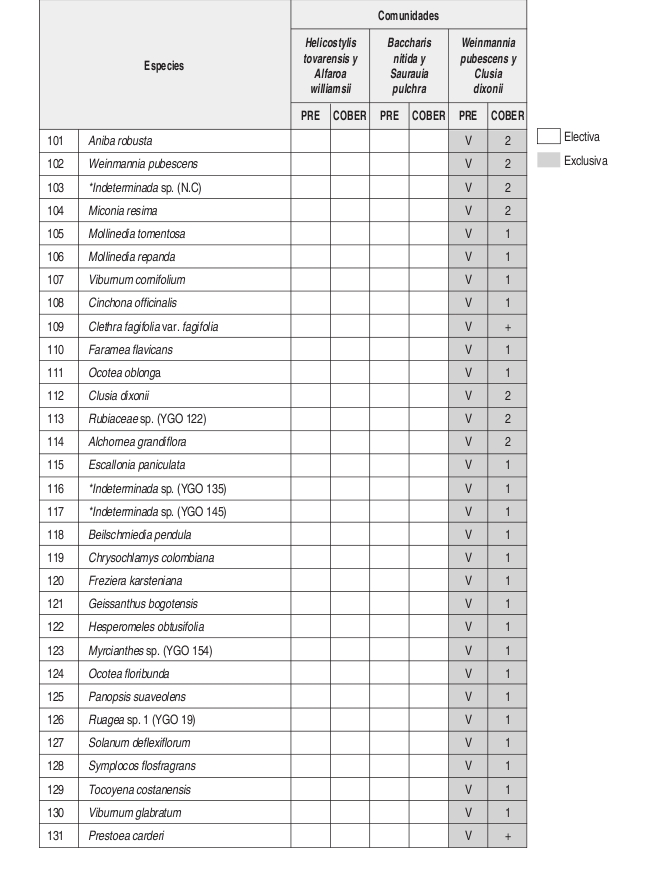
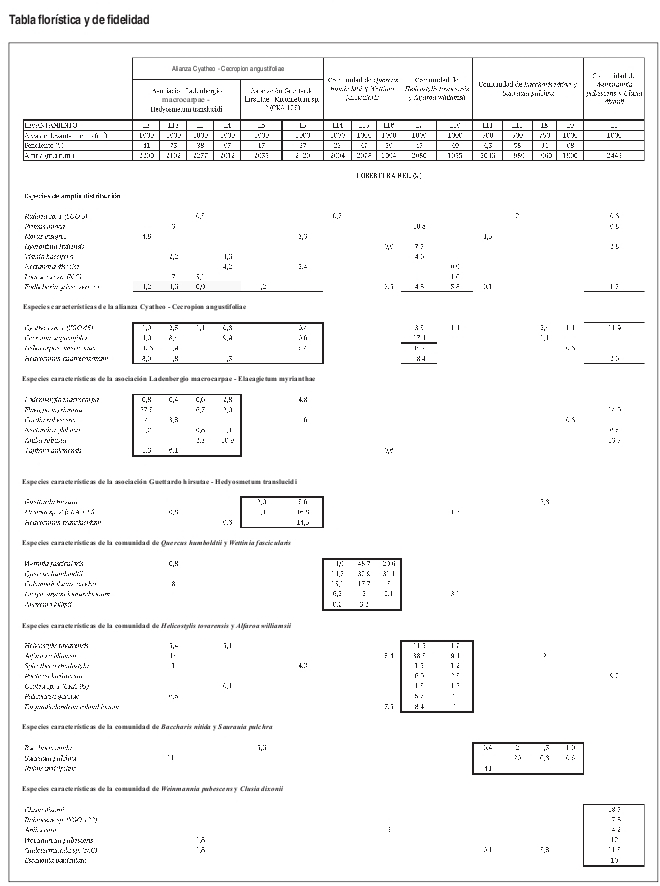
Luego de obtener la información de cada especie, se actualizaron las bases de datos y en base a su presencia y cobertura, se definieron las unidades sintaxonómicas mediante el uso del programa PC ORD con su aplicación automatizada TWINSPAN. También se obtuvo la información de las especies exclusivas y electivas de cada unidad (Anexo 2).
La fase de suelos se dividió en dos, en una primera instancia se tomaron las muestras de barrenadas en zigzag de acuerdo a la metodología usada por Cantillo et al. (2005) donde cada levantamiento se dividió en dos subparcelas de 10 x 50 m, y se obtuvieron dos grandes muestras de barrenadas a 30 cm y 60 cm de profundidad, obteniendo cuatro muestras por levantamiento.
Se realizaron 16 calicatas, donde se describieron cada uno de los horizontes encontrados. Se tomaron datos de color con la ayuda de la tabla Munsell®, textura, consistencia a capacidad de campo y en húmedo, entre otras variables que sólo se pueden obtener en campo como estructura, porosidad, cantidad y grosor de raíces, presencia de cutanes y formaciones especiales.
A su vez, se tomaron datos relacionados con el paisaje, forma y disposición del sitio muestreado, material parental de la zona y de uso actual del suelo.
El análisis de las 64 muestras de suelos, contienen los resultados de textura determinada por el método de Bouyoucos, carbono orgánico (método de Walckley Black), fósforo (Bray y Kurtz), acidez intercambiable (titulación con hidróxido de sodio para pH > 5,4), C.H.% (relación masa de agua, masa de sólidos), Capacidad de Intercambio Catiónico (CIC) (extracción acetato de amonio), Bases intercambiables (acetato de amonio 1 N absorción atómica) y pH.
RESULTADOS
CARACTERIZACIÓN FLORÍSTICA
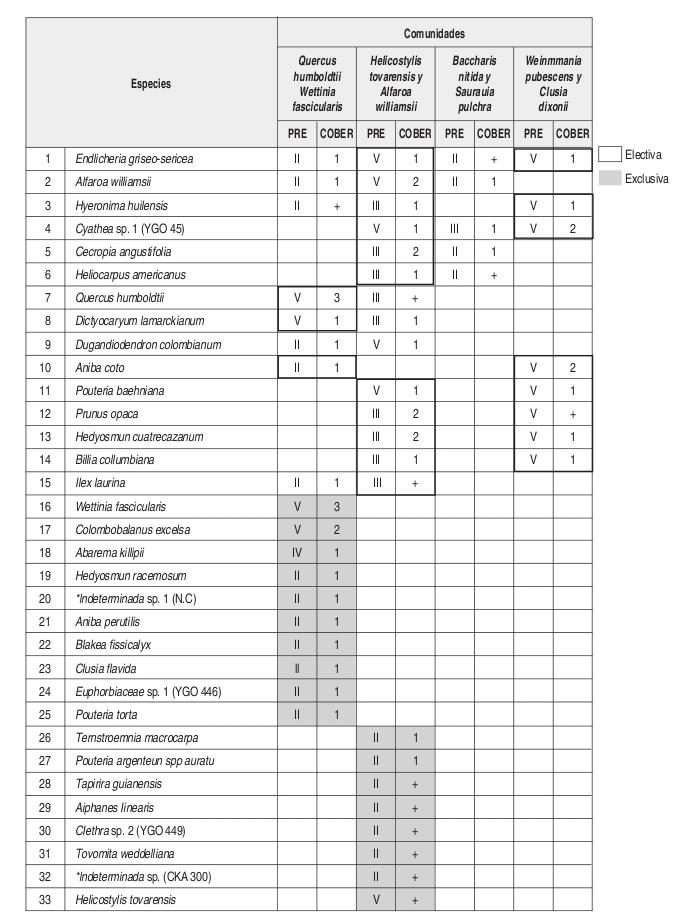
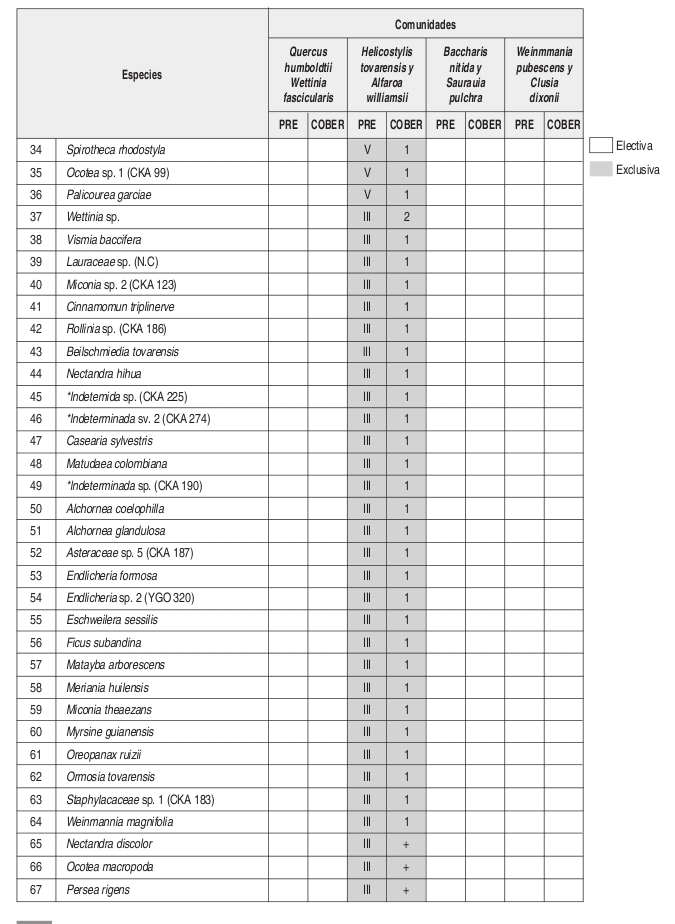
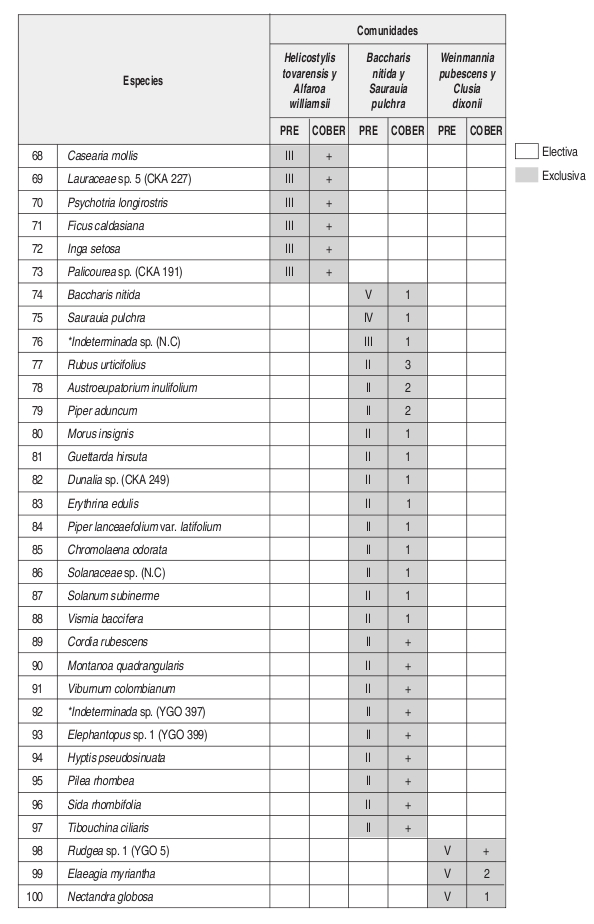
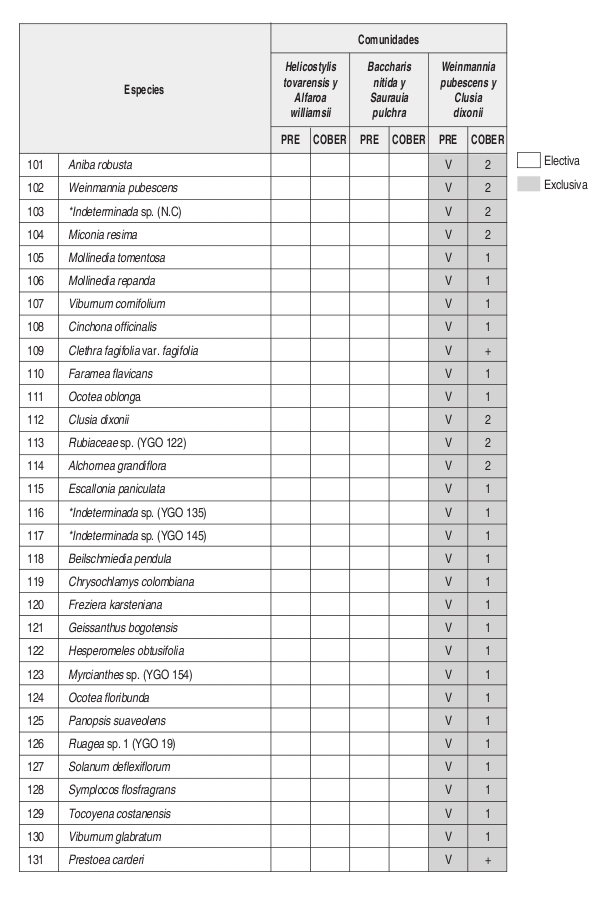
En el análisis florístico, se definió la alianza Cyatheo Cecropion angustifoliae que incluye las asociaciones Ladenbergio macrocarpae Elaeagietum myrianthae y Guettardo hirsutae Hedyosmetum translucidi. Igualmente se reseñaron las comunidades de Helicostylis tovarensis y Alfaroa williamsii, Quercus humboldtii y Wettinia fascicularis, Baccharis nitida y Saurauia pulchra. y la comunidad de Weinmannia pubescens y Clusia dixonii.
El número de levantamientos y altitud a la cual se encontraron cada una de las unidades sintaxonómicas, se puede observar en la tabla florística (Anexo 2).
1. ALIANZA CYATHEO
CECROPION ANGUSTIFOLIAE
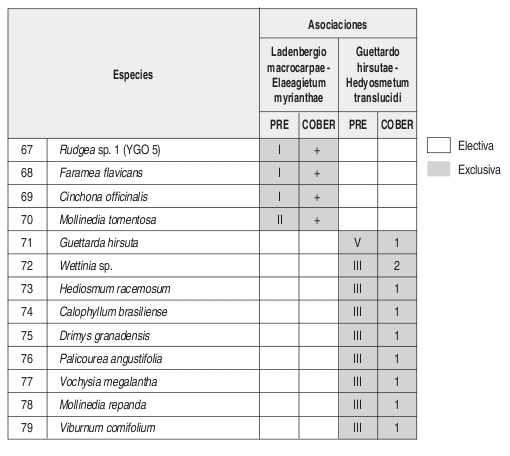
Asociación tipo: Ladenbergio macrocarpae Elaeagietum myrianthae Anexo 1, Figuras 9 y 10.
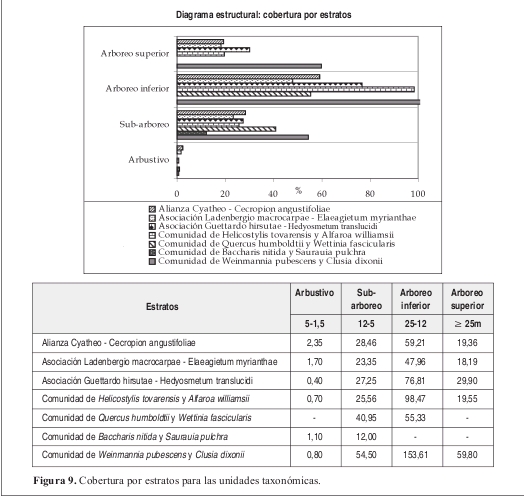
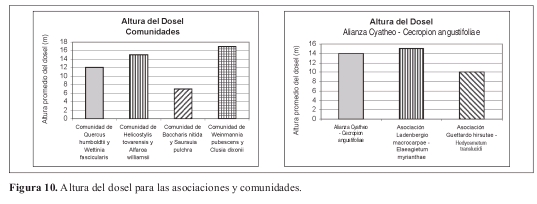
Fisionomía: Bosques con un estrato arbóreo inferior con 59% de cobertura. El estrato subarbóreo ocupa el 28%. (Figura 9). El promedio de alturas es de 13m. (Figura 10).
Las especies más dominantes y abundantes en el estrato arbóreo inferior son Aniba robusta con 23 individuos, Elaeagia myriantha con 21 individuos y Cecropia angustifolia con 19 individuos.
Composición: Las especies características son Ladenbergia macrocarpa, Elaeagia myriantha, Nectandra globosa, Aniba robusta, y Weinmannia pubescens.
Diversidad: Se contabilizaron en total 445 individuos, distribuidos en 105 especies y 45 familias. El estrato de mayor riqueza (para ind. DAP 10cm) es el arbóreo inferior, con 195 individuos y 80 especies, seguido del estrato subarbóreo con 209 individuos y 64 especies. Las familias con mayor número de individuos son Lauraceae y Rubiaceae con 77 y 58 individuos respectivamente. También poseen el mayor número de especies con 17 y 12 especies cada una.
Asociación Ladenbergio macrocarpae Elaeagietum myrianthae Perfil: Figura 3. Levantamiento tipo: Lev. 3, vereda Yarumal. Anexo 1, Figuras 9 y 10.
Fisionomía: El estrato arbóreo inferior cubre el 48%, le sigue el subarbóreo con el 28% de cobertura sobre el área total. (Figura 9). El promedio de alturas obtenido para la asociación fue de 15m, con variación entre 10 y 19 m. (Figura 10).
En el estrato arbóreo inferior las especies dominantes y de mayor abundancia son Cecropia angustifolia y Elaeagia myriantha con 18 individuos y Aniba robusta con 17 individuos.
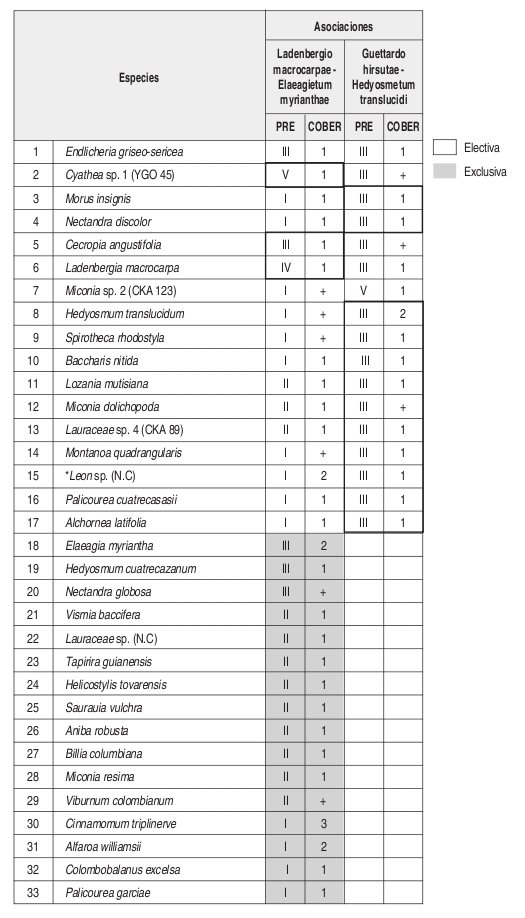
Composición: Las especies exclusivas y con altos valores de fidelidad son Vismia baccifera, Tapirira guianensis, Helicostylis tovarensis, Viburnum colombianum, Cinnamomum triplinerve y Alfaroa williamsii. Como especies electivas importantes se encuentran Cecropia angustifolia y Ladenbergia macrocarpa.
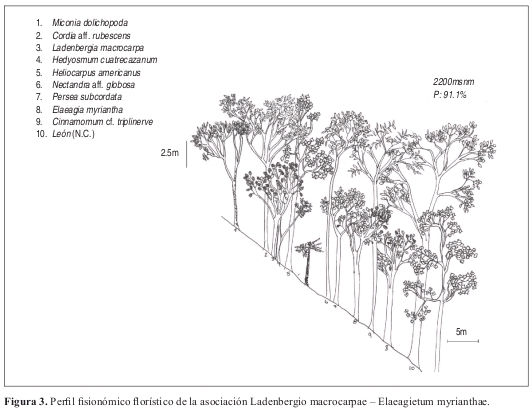
Diversidad: Se estimaron para la asociación 193 individuos, distribuidos en 54 especies por 0.1 ha. El estrato de mayor riqueza es el arbustivo, con 21 especies seguido del arbóreo inferior con 16 especies en 0.1 ha. Las familias más importantes son Lauraceae y Rubiaceae con 56 y 34 individuos y con 14 y ocho especies cada una.
Asociación Guettardo hirsutae Hedyosmetum translucidi. Perfil: Figura 4. Levantamiento tipo: Lev. 5 vereda Los Alpes, Marbella. Municipio de San Agustín Anexo 1, figuras 9 y 10.
Fisionomía: El estrato arbóreo inferior cubre el 77% del área, seguido por el estrato arbóreo superior con el 30%. (Figura 9). El promedio de alturas fue el más bajo registrado con 10m en promedio. (Figura 10).
En el estrato arbóreo inferior Wettinia sp con 17 individuos y Hedyosmum translucidum con siete individuos poseen la mayor abundancia. En el arbóreo superior se encontró la especie León sp. (NC) la cual no pudo ser colectada debido a la altura de sus individuos.
Composición: Las especies exclusivas con altos valores de fidelidad son Guettarda hirsuta y Hedyosmum racemosum. En cuanto a especies electivas se presentan Hedyosmum translucidum, Morus insignis, Nectandra discolor, Spirotheca rhodostyla, entre otras encontradas en zonas con mayor intervención como Baccharis nitida y Montanoa quadrangularis.
Diversidad: Se encontraron 197 individuos y 59 especies por 0.1 ha, el cual es un valor alto debido a que la asociación se definió en las primeras etapas de sucesión. El estrato arbustivo, posee el mayor número de individuos, sin embargo el estrato herbáceo es el de mayor riqueza con los valores más altos de especies y familias. Las familias con mayor número de individuos son Arecaceae y Rubiaceae con 17 y nueve individuos cada una. La familia Rubiaceae es la de mayor riqueza, con cuatro especies.

Comunidad de Helicostylis tovarensis y Alfaroa williamsii Perfil: Figura 5. Levantamiento tipo: Lev. 10 Vereda El Pencil. Anexo 1, Figuras 9 y 10.
Fisionomía: El estrato arbóreo inferior cubre el 98% del área total, le sigue el estrato subarbóreo con el 26% de cobertura. (Figura 9). El promedio de alturas encontrado para la comunidad fue de 15 m. (Figura 10).
En el estrato arbóreo inferior la especie de mayor abundancia es Endlicheria griseo serícea con nueve individuos. Igualmente en el estrato subarbóreo Alfaroa williamsii posee nueve individuos. Hedyosmum cuatrecazanum y Helicostylis tovarensis con siete individuos cada una, son representativas del estrato subarbóreo.
Composición: Las especies exclusivas de esta comunidad son Helicostylis tovarensis, Spirotheca rhodostyla, Pouteria baehniana, Palicourea garciae, Prunus opaca, Hedyosmum cuatrecazanum y Cinnamomum triplinerve. En cuanto a especies electivas se presentan Endlicheria griseo-serícea, Alfaroa williamsii, Cecropia angustifolia y Cyathea sp.
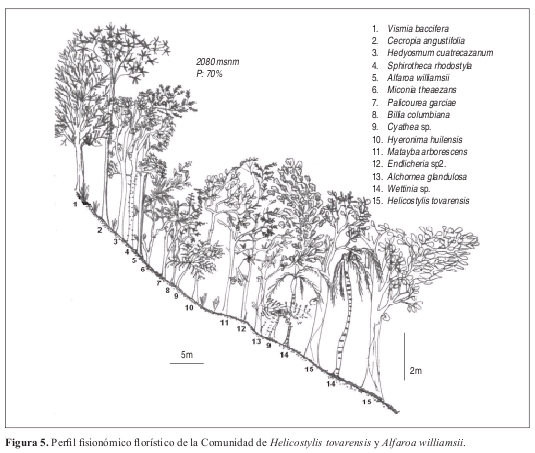
Diversidad: Para esta comunidad se encontraron en total 187 individuos distribuidos en 66 especies por 0.1 ha. La mayor riqueza de especies y familias se encontró en el estrato arbóreo inferior para el levantamiento 7 con 30 especies y 22 familias, mientras que para el levantamiento 10 se encontró en el estrato arbustivo con 26 especies y 15 familias. Las especies más representativas son Lauraceae y Asteraceae con 30 y 11 individuos respectivamente. La familia Lauraceae también posee la mayor riqueza con 13 especies.
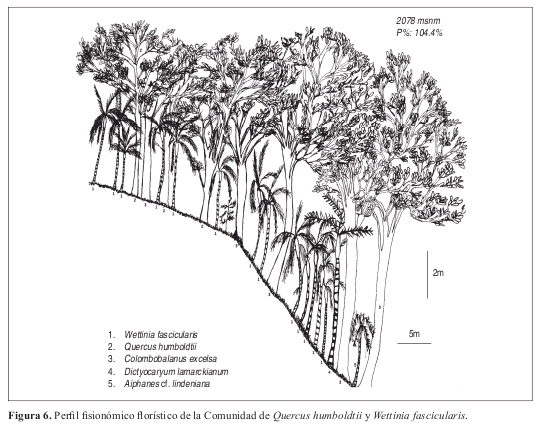
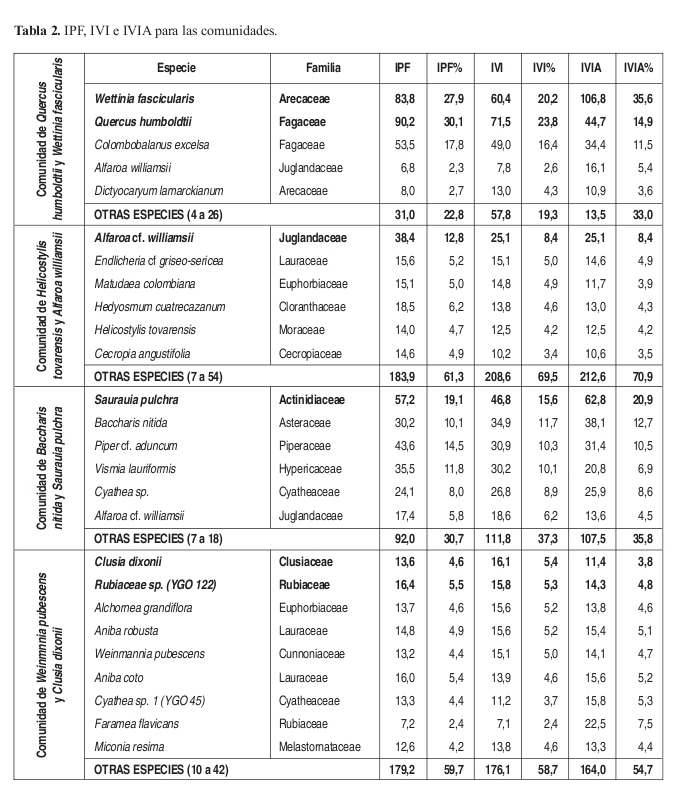
Comunidad de Quercus humboldtii y Wettinia fascicularis Perfil: Figura 6 Levantamiento tipo: 15, Vereda Villa Fátima. Anexo 1, Figuras 9 y 10.
Fisionomía: El estrato arbóreo inferior cubre el 53% del área, seguido por el estrato subarbóreo con el 41%. (Figura 9). El promedio de alturas para la comunidad fue de 12m. (Figura 10). Las especies que tienen mayor abundancia son Quercus humboldtii con 20 individuos y Wettinia fascicularis con 64 individuos para la comunidad. El roble negro Colombobalanus excelsa posee 12 individuos. Se destaca la presencia de Aiphanes linearis, Dugandiodendron colombianum y Abarema killipii.
Composición: Las especies exclusivas más importantes para la comunidad son: Quercus humboldtii, Wettinia fascicularis, Colombobalanus excelsa, Abarema killipii, Blakea fissicalyx, Hedyosmum racemosum, Pouteria argenteum y P. torta, como especie electiva figura Dictyocaryum lamarckianum.
Diversidad: Se presentan 169 individuos y 40 especies por 0.1 ha. El estrato con mayor riqueza es el estrato arbustivo con un promedio de 26 especies. Las familias más representativas de la comunidad son Arecaceae y Fagaceae con 65 y 32 individuos cada una. Arecaceae y Lauraceae poseen cada una tres especies.
Comunidad de Baccharis nitida y Sauarauia pulchra Perfil: Figura 7. Levantamiento tipo: 8, Vereda El Cedro. Anexo 1, Figuras 9 y 10.
Fisionomía: El estrato con mayor cobertura es el subarbóreo, con 12% del área (Figura 9). El promedio de alturas obtenido fue de 7 m (Figura 10).
Las especies con mayor abundancia en el estrato subarbóreo fueron Saurauia pulchra con diez individuos, Piper lanceaefolium var latifolium con nueve individuos y Baccharis nitida con seis individuos. También es importante resaltar la presencia de Alfaroa williamsii en casi todos los estratos.
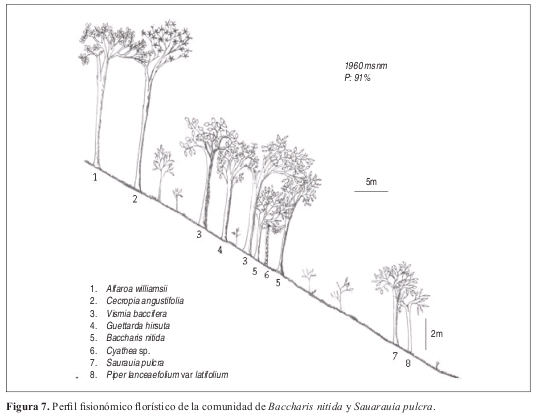
Composición: Las especies exclusivas para la comunidad y que además obtuvieron los mayores valores de presencia y cobertura fueron Baccharis nitida, Saurauia pulchra, Rubus urticifolius, Piper lanceaefolium var latifolium y Austroeupatorium inulifolium.
Diversidad: El estrato rasante posee la mayor riqueza con 18 especies sin embargo en el levantamiento nueve se presenta la mayor riqueza en el estrato arbustivo con 26 especies. Las familias con mayor abundancia de individuos son Actinidiaceae y Piperaceae con diez y nueve individuos respectivamente. Las familias con mas número de especies fueron Solanaceae, Rubiaceae y Asteraceae. Se encontraron 126 individuos y 36 especies con DAP mayores a 10 cm en 0.1 ha.
Comunidad de Weinmannia pubescens y Clusia dixonii Perfil: Figura 8. Anexo 1, Figuras 9 y 10.
Fisionomía: El estrato con mayor cobertura (dominancia) fue el arbóreo inferior cubriendo el 100% de la parcela. El estrato arbóreo superior obtuvo un 60% de cobertura (Figura 9). El promedio de alturas de la vegetación de esta comunidad fue el más alto de las demás con 17 m (Figura 10).
Las especies que tienen mayor cantidad de individuos en el estrato arbóreo inferior son Aniba coto con siete individuos, A. robusta con seis individuos, Miconia resima y Weinmannia pubescens con cinco individuos cada una. La especie más abundante y de mayor cobertura en el estrato arbóreo superior es Clusia dixonii con tres individuos.
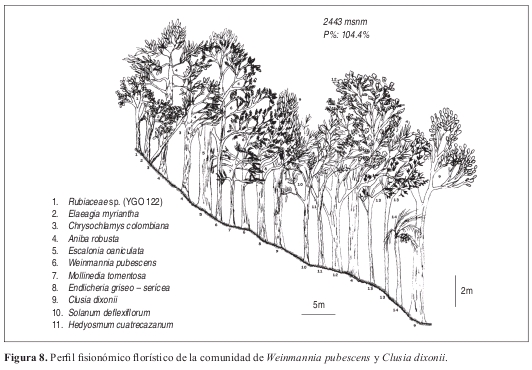
Composición: Las especies exclusivas de la comunidad son Clusia dixonii, Aniba coto, Alchornea grandifolia, Weinmannia pubescens, Hyeronima huilensis, Escallonia paniculata, Cinchona officinallis y Freziera carsteniana, por su parte las especies Endlicheria griseo serícea, Miconia resima, Aniba robusta y Elaeagia myriantha se consideraron las especies electivas de mayor importancia dentro de la comunidad.
Diversidad: Respecto al número de individuos colectados, se deduce que se pueden encontrar en este bosque 132 individuos y 45 especies por 0.1 ha. La riqueza de los estratos demuestra que el estrato herbáceo es el de mayor importancia con 35 especies, mientras que en riqueza de familias, el estrato subarbóreo posee el mayor valor con 19 familias. Las familias más abundantes y más ricas dentro de la asociación son Lauraceae y Rubiaceae con 20 y 15 individuos y con siete y seis especies respectivamente.
ESTRUCTURA
ALIANZA CYATHEO CECROPION
ANGUSTIFOLIAE
Estructura horizontal (Anexo 1) Distribución de diámetros (Figura 11) Se encontraron ocho clases diamétricas, desde la clase I (10-20 cm) hasta la clase IX (90 cm 100 cm). El 58% de los individuos se concentraron en la clase I (10.1 20 cm), mientras que en la clase II (20.1 30 cm) se encontró el 23% del total. La tendencia se da en J invertida.
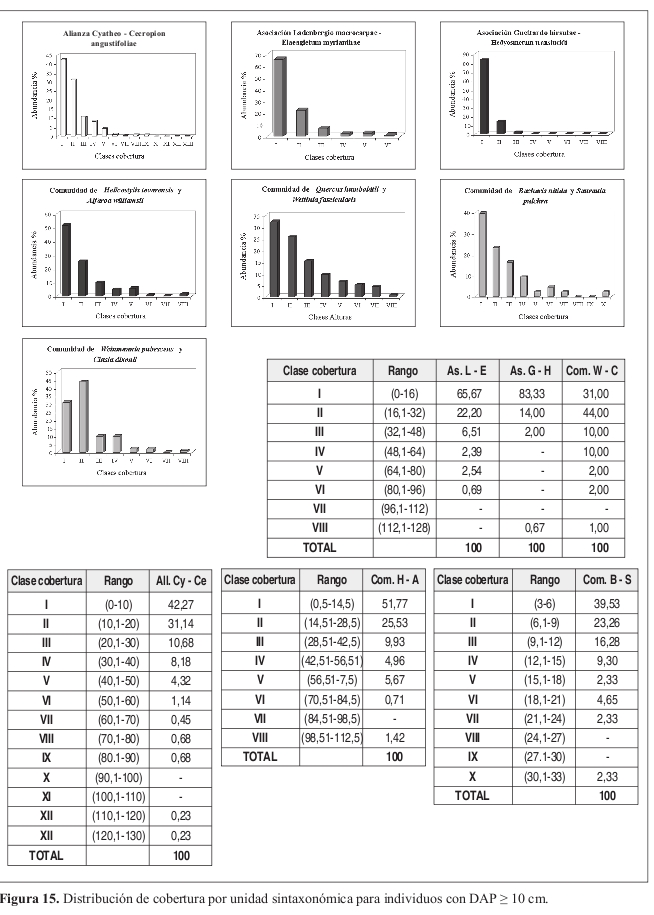
Distribución de coberturas (Figura 15) Se distribuye en 13 clases, con coberturas desde 1 m² hasta 130 m² y rangos con intervalos de 10 m². Se presenta una distribución en J invertida. En la clase I (1 10 m²) se presenta la mayor concentración de individuos con el 42% del total.
Área basal
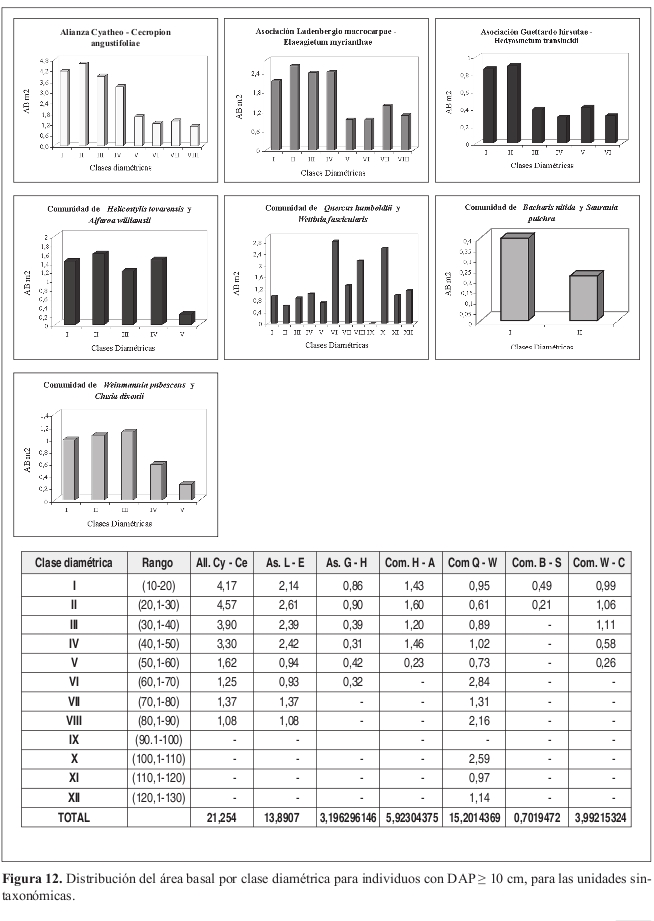
Las especies Elaeagia myriantha y Aniba robusta poseen los valores más altos de área basal con 2.05 y 1.65 m² respectivamente. Área basal por clases diamétricas (Figura 12)
Se presenta una uniformidad en los valores de área basal en las primeras cuatro clases, el valor de área basal para la clase II (20.1 30 cm) es de 4.6 m², seguido de la clase III (30.1 40 cm) con 3.9 m².
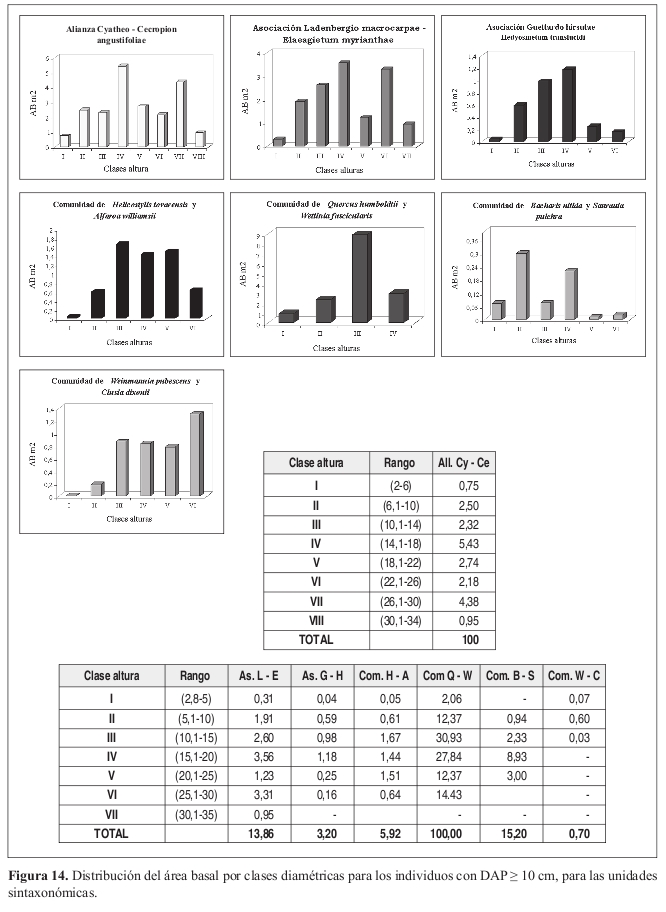
Área basal por clases de altura (Figura 14) Las clases IV (14.1 18 m) y VII (30.1 34 m) presentan los valores más altos de área basal con 5.4 y 4.4 m² respectivamente.
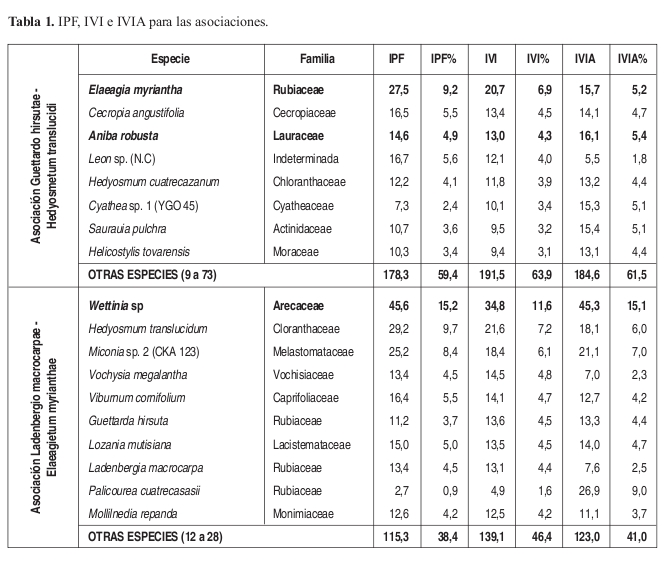
Índice de Predominio Fisionómico (IPF), de Valor de Importancia (IVI) y de Valor de Importancia Ampliado (IVIA) (Tabla 1) En los valores de IPF e IVI, las especies Elaeagia myriantha y Aniba robusta ocupan el primer y segundo lugar con 5 y 4% en IPF y 3% en IVI para cada una. Las especies A. robusta y Cyathea sp. (YGO 045) presentan los mayores valores de Regeneración natural (Rn) y poseen el valor más alto en IVIA con un 4%.
Estructura vertical
Distribución de alturas (Figura 13)
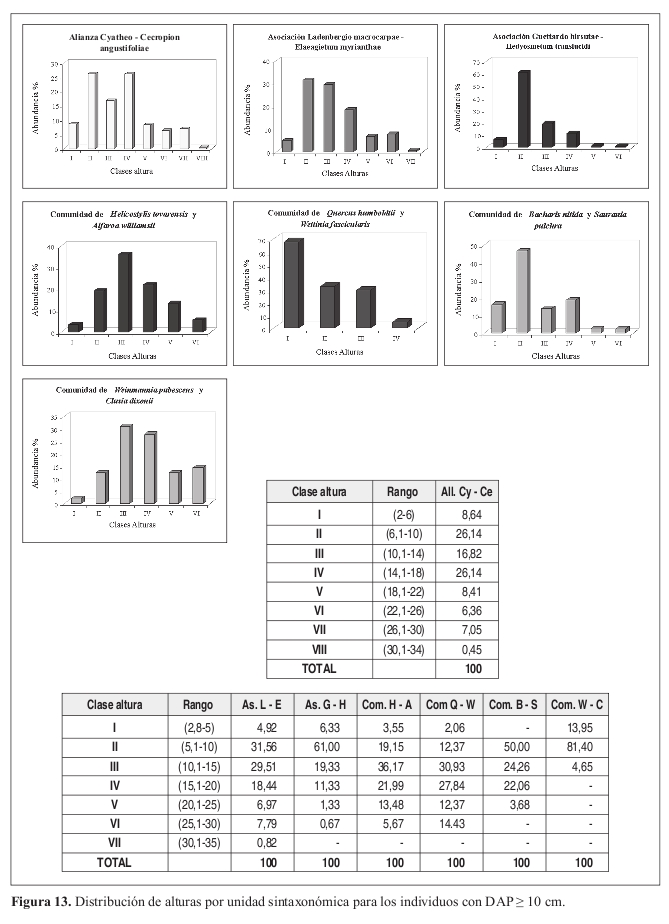
Se encontraron ocho clases con una tendencia a la normalidad, con variación entre 4 y 32 m. La mayor cantidad de individuos se presenta en las clases II (6.1 10 m) y IV (14.1 18 m) con el 26%.
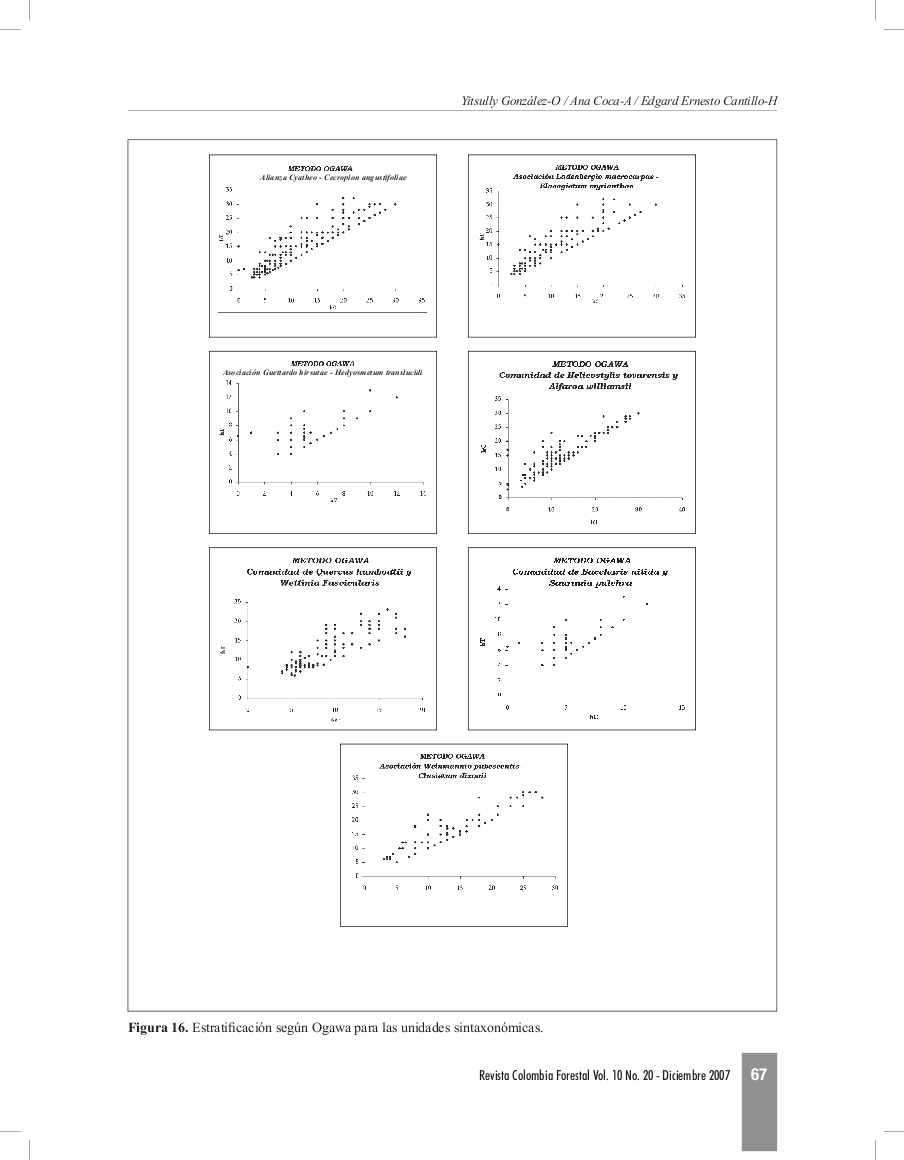
Estratificación según Ogawa (Figura 16) Se presentan alturas desde los 4 hasta los 32 m. Se muestra una concentración de puntos que indica la formación de estratos en los individuos con alturas entre los 5 y 12 m, igualmente se presenta una ligera agrupación entre los 12 y 20 m y los individuos mayores a 20 m de altura. Asociación Ladenbergio macrocarpae
Elaeagietum myrianthae
Estructura horizontal
Distribución de diámetros (Figura 11) La distribución de los individuos se presenta en ocho clases diamétricas. La clase I (10.1 20 cm) obtuvo el mayor porcentaje de individuos con el 54%, la clase II (20.1 30 cm) obtuvo el 23%. El patrón se da en J invertida.
Distribución de coberturas (Figura 15) Se presentan seis clases, con una tendencia en J invertida y coberturas hasta los 96 m². El 66% de los individuos se concentra en la clase I (0.1 16 m²), en la clase II (16.1 32 m²) se agrupa el 22% del total de individuos.
Área basal
Los valores más altos de área basal por especie fueron para Cecropia angustifolia y una especie no identificada Leon sp. (NC) con 1.55 y 1.36m² respectivamente.
Área basal por clases diamétricas (Figura 12)
La distribución es uniforme en las cuatro primeras clases, con una concentración promedio de individuos en las clases II (20.1--30 cm) a IV (40.1 50 cm) de 2.5 m². A diferencia de los demás, el levantamiento 13 presentó una distribución en J invertida.
Área basal por clases de altura (Figura 14) La clase IV (15.1 20 m) presenta el mayor valor de área basal con 3.5 m², seguido de la clase VI (25.1 30 m) con 3.3 m². En este caso la distribución tiende a la normalidad.
Índice de Predominio Fisionómico (IPF), de Valor de Importancia (IVI) y de Valor de Importancia Ampliado (IVIA) (Tabla 1) Para este caso Elaeagia myriantha obtuvo los valores más altos en IPF e IVI con 9 y 7%, seguida de Cecropia angustifolia con 5 y 4% respectivamente. Aniba robusta y E. myriantha obtuvieron el porcentaje más alto en IVIA con un 5% cada una. Palicourea cuatrecasasii, posee el valor más alto en Rn.
Estructura vertical
Distribución de alturas (Figura 13) Se encontraron siete clases con variación entre 5 y 35 m. La clase II (5.1 10 m) presenta la mayor concentración de individuos con un 31%, seguida de la clase III (10.1 15 m) con el 29%. La distribución de los individuos es normal con una ligera desviación hacia la izquierda.
Estratificación según Ogawa (Figura 16) Se presentan dos estratos. Una concentración de individuos marcada entre los 5 y 15 m de altura, y otra ligeramente definida entre los 15 y 20 m.
Asociación Guettardo hirsutae
Hedyosmetum translucidi.
Estructura horizontal
Distribución de diámetros (Figura 11) La distribución se da en seis clases diamétricas. Se presenta una distribución en J invertida. La mayor cantidad de individuos se concentra en la clase I (10.1 20 cm) con el 79% seguida de la clase II (20.1 30 cm) que agrupa el 14% del total.
Distribución de coberturas (Figura 15) La tendencia se da en J invertida con una distribución en 16 clases, la mayor concentración se presenta en la clase II (16.1 32 m²) con el 52%. La clase I (0.1 16 m²) obtuvo el 32% del total de individuos.
Área basal
Los valores más altos de área basal por especie son para Wettinia sp. con 0.37 m², y para Hedyosmum translucidum y Vochysia megalantha con 0.32 m² cada una.
Área basal por clases diamétricas (Figura 12)
Las clases II (20.1 30 cm) y I (10.1 20 cm) tienen los mayores valores, con 0.9 y 0.86 m² respectivamente.
Área basal por clases de altura (Figura 14) Los mayores valores de área basal se presentan en la clase IV (15.1 20 m) con 1.2 m². La clase III (10.1-15m) obtuvo 1 m².
Índice de Predominio Fisionómico (IPF), de Valor de Importancia (IVI) y de Valor de Importancia Ampliado (IVIA) (Tabla 1) En los índices de importancia, Wettinia sp. posee los valores más altos, en IPF (15%), IVI (11%) e IVIA (15%). le sigue Hedyosmum translucidum en IPF (10%) e IVI (7%). Palicourea cuatrecasasii (con el valor más alto en Rn) y Miconia sp. (CKA 123) también sobresalen en IVIA con 9 y 7% respectivamente.
Estructura vertical
Distribución de alturas (Figura 13) Se obtuvieron seis clases con una distribución normal. El 61% de los individuos se concentra en la clase II (5.1 10 m) y el 19% de los individuos en la clase III (10.1 15 m).
Estratificación según Ogawa (Figura 16) Se definieron ligeramente dos estratos, uno entre los 5 y 15 m y otro con los individuos emergentes de alturas superiores a los 15 m.
Comunidad de Helicostylis tovarensis y Alfaroa williamsii
Estructura horizontal Distribución de diámetros (Figura 11) Se definieron cinco clases diamétricas para la comunidad. El 60% del total de individuos se concentra en la clase I (10.1 20 cm), la clase II (20.1 30 cm) obtuvo el 24% de los individuos. La distribución se da en J invertida.
Distribución de coberturas (Figura 15) Se definieron ocho clases de cobertura, la mayor agrupación de individuos se presenta en la clase I (0.5 14.5 m²) con el 52% del total. La clase II (14.51 28.5 m²) concentra el 26%.
Área basal
Alfaroa williamsii y Matudaea colombiana poseen los mayores valores en área basal con 0.92 y 0.62 m² respectivamente.
Área basal por clases diamétricas (Figura 12) Se encontró una uniformidad en los valores en las cuatro primeras clases con valores entre 1.2 m² en la clase III (30.1 40 cm) y 1.6m² para la clase II (20.1 30 cm).
Área basal por clases de altura (Figura 14) Las clases III, IV y V presentan valores de área basal muy similares, con valores de 1.4 m² en la clase IV (15.1 20 m) a 1.7 m² en la clase III (10.1 15 m).
Índice de Predominio Fisionómico (IPF), de Valor de Importancia (IVI) y de Valor de Importancia Ampliado (IVIA) (Tabla 2) La especie Alfaroa williamsii, posee los valores más altos en los tres índices con un 13% en IPF, 8% en IVI e IVIA. En IPF Hedyosmum cuatrecazanum posee el 6%. Se destacan Endlicheria cf. griseo sericea con 5% en IVI y 4% en IVIA y Psychotria longirostris con 4% en IVIA, pese a que no tiene valores altos en los demás índices.
Estructura vertical
Distribución de alturas (Figura 13) Se presentan seis clases de alturas con valores de 2.8 a 30 m. Se presenta una normalidad en los valores con el 36% de los individuos concentrados en la clase III (10.1 15 m).
Estratificación según Ogawa (Figura 16) No se diferencia un estrato en particular dentro de la asociación, sin embargo algunos puntos se concentran entre los 5 y 15 m de altura. Comunidad de Quercus humboldtii y Wettinia fascicularis
Estructura horizontal
Distribución de diámetros (Figura 11) Se presenta una distribución en 12 clases diamétricas, con algunos individuos aislados pertenecientes a las dos últimas clases. Se presenta una distribución en J invertida, la mayoría de los individuos se concentran en la clase I (10.1 20 cm) con el 62%, la clase II (20.1 30 cm) posee el 8% del total.
Distribución de coberturas (Figura 15) Se presentan ocho clases de cobertura. La tendencia se da en J invertida. La concentración de individuos se presenta en la clase I (2.1 11 m²) representando el 32%, aunque en el levantamiento No. 15 los individuos se concentran en la clase II (11.1 20 m²), con el 35%.
Área basal
Las dos especies que dominan en la comunidad son Quercus humboldtii, y Colombobalanus excelsa. Presentan valores de área basal de 7.34 y 4.6 m² respectivamente frente a 0.34 m² de Wettinia fascicularis, especie que es característica de la comunidad por su abundancia y representatividad.
Área basal por clases diamétricas (Figura 12)
Se presenta una tendencia normal con desviación a la derecha, los mayores valores de área basal se presentan en la clase VI (40.1 50 cm) y X (100.1 110 cm) con 2.8 y 2.5 m² respectivamente.
Área basal por clases de altura (Figura 14) La distribución de los valores de área basal tiende a la normalidad, en la clase III (15.1 20 m) se presenta un valor de 8.9 m².
Índice de Predominio Fisionómico (IPF), de Valor de Importancia (IVI) y de Valor de Importancia Ampliado (IVIA) (Tabla 2) En IPF e IVI dominan Quercus humboldtii con un IPF de 30% e IVI de 24%, Wettinia fascicularis por su parte posee un 28% en IPF y 20% en IVI, Colombobalanus excelsa también es importante con un 18% y 16% para cada índice. W. fascicularis posee el valor más alto de IVIA con un 36% y el valor más alto de Rn, seguido de Q. humboldtii y C. excelsa con un 15% y 11% respectivamente.
Estructura vertical
Distribución de alturas (Figura 13) Se observa una distribución en cuatro clases, con alturas entre 5 y 25 m. La mayor cantidad de individuos se concentra en la clase I (5.1 10 m) con el 55% del total de individuos. La distribución se asemeja a una J invertida.
Estratificación según Ogawa (Figura 16) Se definen tres estratos. Entre los 5 y 12 m se presenta un estrato, se observa otro estrato hasta los 20m y finalmente se observa una concentración de puntos entre 15 y 23 m de altura.
Comunidad de Baccharis nitida y Saurauia pulchra
Estructura horizontal
Distribución de diámetros (Figura 11) Se encuentran dos clases diamétricas. Los individuos se concentran en la clase I (10.01 20 cm), con el 62% del total, la clase II (20.1 30 cm) agrupa el 38% restante. La distribución en seis clases que se presenta con amplitudes de 3cm entre clases, muestra que el 58% del total de individuos se concentra en la clase I (10.1 13 cm).
Distribución de coberturas (Figura 15) Se presentan diez clases de cobertura con una distribución en J invertida, el 40% de los individuos se concentran en la clase I (3.1 6 m²). En la clase II (6.1 9 m²) se agrupa el 23% del total de individuos.
Área basal
Aunque se presentan valores bajos de área basal, las especies más dominantes son Vismia baccifera con 0.15 m² y Piper lanceaefolium var latifolium con 0.11 m².
Área basal por clases diamétricas (Figura 12) No se presenta un patrón definido. En la clase I (10.1 20 cm) el valor más alto se presentó en la clase I con 0.49 m². En la clase II (20.1 30 cm) se obtuvo 0.21 m².
Área basal por clases de altura (Figura 14) El valor más alto se presentó en la clase II (5.1 10 m) con 0.6 m², seguido de la clase I con 0.07 m².
Índice de Predominio Fisionómico (IPF), de Valor de Importancia (IVI) y de Valor de Importancia Ampliado (IVIA) (Tabla 2) Los valores más altos de IPF se presentan para Saurauia pulchra (que posee los valores más altos de Rn) y Piper lanceaefolium var latifolium con el 19 y 18% del total. En IVI también poseen los valores más altos con 14% para ambas especies. En IVIA S. pulchra obtuvo el 21% y P. lanceaefolium var latifolium el 13%.
Estructura vertical
Distribución de alturas (Figura 13) Se encuentran tres clases con valores que van desde los 4.5 hasta los 13 m. Se presenta una distribución normal. La clase II (5.1 10 m) representa el 81%, seguida por la clase I con el 14% del total.
Estratificación según Ogawa (Figura 16) Se presenta una ligera concentración de individuos desde los 4,5 hasta los 10 m de altura, sin embargo no es muy notoria la estratificación debido al estado inicial de sucesión en la que se encuentran estas áreas.
Comunidad de Weinmannia pubescens y Clusia dixonii
Estructura horizontal
Distribución de diámetros (Figura 11) Se diferencian ocho clases diamétricas, la mayor concentración de individuos se presenta en la clase I (10.1 20 cm) con el 60% del total. La clase II (20.1 30 cm) concentra el 24% del total de individuos. La distribución se da en J invertida.
Distribución de coberturas (Figura 15) Se presenta una distribución normal en ocho clases con una fuerte tendencia hacia la izquierda, con valores hasta los 128 m². La concentración de individuos se presenta en la clase II (16.1 32 m²) con el 44%, en la clase I (0.1 16 m²) se encuentra el 31% del total.
Área basal
En área basal Clusia dixonii y Escallonia paniculata poseen los valores más altos con 0.37 y 0.31 m² respectivamente.
Área basal por clases diamétricas (Figura 12)
Los mayores valores de área basal se presentan en la clase III (30.1 40 cm) con 1.11 m² seguido de la clase II (20.1 30 cm) con 1.05 m². Área basal por clases de altura (Figura 14) La clase VI (25.1 30 m) presenta los mayores valores de área basal con 1.3 m². La distribución de las clases III (10.1 15 m) a V (20.1 25 m) presentan uniformidad en sus valores, con un promedio de 0.8 m².
Índice de Predominio Fisionómico (IPF), de Valor de Importancia (IVI) y de Valor de Importancia Ampliado (IVIA) (Tabla 2) En los valores de IPF la especie no identificada Rubiaceae sp. (YGO 122) y Aniba coto, obtuvieron los valores más altos con 5% cada una. En los valores de IVI Clusia dixonii y Rubiaceae sp. (YGO 122) poseen los valores más altos con 5% respectivamente. En IVIA sobresale Faramea flavicans (7%), con el valor más alto de Rn, Cyathea sp. (YGO 045) y nuevamente A. robusta con el 5% cada una.
Estructura vertical
Distribución de alturas (Figura 13) Se encontraron seis clases de alturas, con variaciones desde los 5 hasta los 30 m. La tendencia es normal, el 31% de los individuos se concentra en la clase III (10.1 15 m), mientras que el 28 % lo hace en la clase IV (15.1 20 m).
Estratificación según Ogawa (Figura 16) Se encontraron individuos desde los 5 hasta los 32 m de altura, no se definen estratos, se observa una agrupación de individuos entre los 5 y 20 m y otra con los individuos mayores a 20 m que corresponden a los individuos dominantes.
DISCUSIÓN Y CONSIDERACIONES FINALES COMPOSICIÓN FLORÍSTICA
Basados en la tabla florística y de fidelidad (Anexo 2) se observa que la asociación Ladenbergio macrocarpae Elaeagietum myrianthae, presenta la mayor riqueza de especies y que la comunidad de Weinmannia pubescens y Clusia dixonii, presenta un mayor endemismo local o "exclusividad" que las demás unidades sintaxonómicas, probablemente su ubicación en una zona de transición de la región de vida subandina a la andina (2443 m) causa que se presenten especies que pertenecen a mayores alturas.
VALORES DE ABUNDANCIA Y RIQUEZA PARA CADA ESTRATO
En cuanto la diversidad por estratos, en las asociaciones el estrato con mayor número de especies es el arbustivo. En las comunidades la mayor concentración de especies se presentó en el estrato herbáceo a excepción de la comunidad de Quercus humboldtii y Wettinia fascicularis que posee el mayor número de especies en el estrato arbustivo. En la comunidad de Baccharis nitida y Saurauia pulchra debido a su estado sucesional, el estrato rasante fue el de mayor riqueza.
La homogeneidad en los valores de cobertura obtenidos para la comunidad de Quercus humboldtii y Wettinia fascicularis es causada por la gran abundancia de individuos en el estrato subarbóreo representados por Wettinia fascicularis que nivelaron los valores de dominancia entre los dos estratos.
La comunidad de Baccharis nitida y Saurauia pulchra se encuentra compuesta por levantamientos realizados en áreas intervenidas, con presencia de elementos arbóreos. La dominancia del estrato subarbóreo en esta comunidad se debe a la presencia de individuos remanentes de bosque y al desarrollo de algunas especies heliófitas de mayor porte dentro de la comunidad.
En las asociaciones no se presentaron variaciones notables en los valores de distribución de diámetros; de esta manera en áreas de mayor intervención se definieron menos cantidad de clases diamétricas. Para las comunidades, las diferencias en la distribución de clases se atribuyen a las diferencias florísticas entre ellas.
La distribución de coberturas nos indica que en la asociación Guettardo hirsutae Hedyosmetum translucidi se encuentran menores coberturas por individuo respecto a la asociación Ladenbergio macrocarpae Elaeagietum myrianthae, dado por la concentración de individuos en la primera clase.
En la Comunidad de Helicostylis tovarensis y Alfaroa williamsii, se presenta mayor concentración de individuos en la primera clase respecto a las demás comunidades. Al comparar las comunidades definidas para áreas boscosas se observa que la homogeneidad en la distribución de copas hace que se presente mayor diversidad y la presencia de individuos con altas coberturas hace que se origine una mayor dominancia y por lo tanto disminuya la diversidad.
Los índices de valor de importancia tienen una relación indirecta con la diversidad. En la comunidad de Weinmannia pubescens y Clusia dixonii se presenta uniformidad en los valores de IPF e IVI por lo cual la importancia ecológica y fisionómica de las especies está igualmente repartida. En la comunidad de Quercus humboldtii y Wettinia fascicularis sucede lo contrario, los valores de importancia recaen sobre unas pocas especies, disminuyendo la diversidad.
Aunque los bosques muestreados en el corredor biológico se encuentran en la región de vida subandina, dentro de ellos se presenta un alto endemismo reflejado en las diferencias florísticas de cada bosque y con excepción de los robledales, en la equitatividad en los índices de importancia ecológica y fisonómica de las especies; por lo que su adecuado manejo será fundamental para el mantenimiento de los bienes y servicios que se desprenden de su conservación.
El conocimiento de la composición florística y de la estructura de las áreas intervenidas sirve de apoyo para realizar proyectos de manejo y restauración de estas zonas. La reducción continua de la cobertura boscosa origina un incremento importante de las áreas de "rastrojo", convirtiéndose en un componente principal en los proyectos forestales. Esta tendencia hace que a futuro el estudio de la conformación y dinámica de las áreas intervenidas sea cada vez más relevante.
AGRADECIMIENTOS
Agradecemos a la Corporación Autónoma del Alto Magdalena CAM por el financiamiento del presente trabajo; a la Ingeniera Gloria Ramírez quien fue nuestro apoyo logístico durante la fase de ejecución; al grupo técnico del corredor biológico y los auxiliares de campo por su ayuda oportuna durante la fase de campo; al Herbario Nacional Colombiano (COL) como al Herbario Forestal de la Universidad Distrital "Francisco José de Caldas" (UDBC) y a sus directores Julio Betancourt y Rocío Cortés-B por brindarnos el espacio necesario para la identificación; a los especialistas y al Ingeniero Germán Tellez por la asesoría en la identificación de las especies.
Al Doctor Orlando Rangel Ch., por sus valiosos aportes en la edición de este documento. Al Ingeniero Harold Garay por el análisis Twinspan en la parte ecológica y al laboratorio de suelos de la Universidad Distrital por la caracterización de las muestras de suelos.
REFERENCIAS BIBLIOGRÁFICAS
Bello, A. & Kattib. 1994. Aspectos ecológicos en la sucesión secundaria de la vegetación subandina (2010 2510 m) en localida des del municipio de Santa Rosa de Cabal. Risaralda, Colombia. Trabajo de grado. Departamento de Biología. Universidad Nacional. Bogotá.
Bernal, A. 1973. Estudio de la vegetación y planificación preliminar del Parque Nacio nal Natural Cueva de los Guácharos, Aceve do, Huila. Bogotá D.C., Colombia. Trabajo de grado (Ingeniero Forestal). Universidad Distrital Francisco José de Caldas.
CCAD-PNUD/GEF. 2002. Corredor biológi co mesoamericano, una plataforma para el desarrollo sostenible regional. Serie técni ca No. 1. Proyecto para la consolidación del corredor biológico mesoamericano. Managua, Nicaragua. 24 p.
Cantillo, E., K. Rodríguez & A. Avella. 2005. Caracterización florística, estructu ral, diversidad y ordenación de la vegeta ción arbórea en la reserva forestal Cárpatos (Guasca Cundinamarca) Universidad Distrital "Francisco José de Caldas" Inge niería Forestal.
Cleef, A. 1972. Informe preliminar sobre la botánica del Parque Nacional Natural Cue va de los Guácharos. Bogotá. Colombia.
Coca, C. & Y. González. 2007. Caracteriza ción florística y estructural de la vegetación del corredor biológico entre los Parques Nacionales Naturales Puracé Cueva de los Guácharos. Trabajo de grado (Ingenie ro Forestal). Universidad Distrital Francis co José de Caldas. Bogotá D.C., Colombia. 282 p.
González Y., C. Coca & G. Téllez. 2008. Ca tálogo de la flora vascular de los bosques andinos y subandinos del corredor biológi co entre los Parques Nacionales Naturales Puracé Cueva de los Guácharos. Univer sidad Distrital Francisco José de Caldas.
Cuatrecasas, J. 1958. Aspectos de la vegeta ción natural de Colombia. Rev. Acad. Co lomb. Cienc. 10(40): 221-268.
Duque-N., A. 1987. Comunidades vegetales de la zona paramuna del PNN Puracé (Co lombia). Tesis de grado, Universidad del Valle. (Mimeografiado).
Duque-N., A. & J.O Rangel-Ch. 1989. Aná lisis fitosociológico de la vegetación para muna del PNN Puracé. En: L. F. Herrera, R. Drenan y C. Uribe (eds). Cazicazgos prehispánicos del Valle de la Plata. Tomo I. El contexto medioambiental de la ocu pación humana. U. of Pittsburg memoirs in Latin América-Archaeology. 2:70-95.
Holdridge, L. R. 1979. Ecología basada en zonas de vida. Costa Rica. Publicaciones del IICA.
Lozano, G., J. Hernández & J. Henao 1979. El género Trigonobalanus en el Neotrópi co. I. Caldasia 12: 517 537.
Mahecha, G. E. 1999. Estudio florístico de la región del macizo colombiano. Principios y procedimientos para el conocimiento de las plantas superiores del trópico america no. Convenio Intercorporativo del Macizo Colombiano. Huila, Colombia.
Rangel-Ch., J. O. & P. Franco R. 1985. Ob servaciones fitoecológicas en varias re giones de vida de la cordillera central de Colombia. Caldasia 14 (67): 210-249.
Rangel-Ch., J. O. & G. Lozano 1986. Un perfil de vegetación entre la Plata Huila y el volcán del Puracé. Caldasia 14 (68-70) 503 547.
Rangel-Ch., J. O., A. M. Cleef, T. Van Der Hammen & R. Jaramillo 1982. Tipos de vegetación en el transecto Buritaca La Cumbre Sierra Nevada de Santa Marta (en tre 0 y 4100 msnm). Colombia geográfica 10(1): 1-18.
Rangel-Ch., J. O., A. M. Cleef & S. Sala manca 2003. The ecuatorial interandean and subandean forest of the Parque los Ne vados Transect, Cordillera Central, Colom bia. 143-204 p. En: T. van der Hammen & A. Dos Santos (eds). La Cordillera Central Colombiana. Estudio de Ecosistemas Tro pandinos 5. J. Cramer, Berlín-Stuttgart.
Rojas, O. 1984. Evolución de una sucesión vegetal en el Valle del Cauca. Cespedesia 13 (49 50) 152-211.
Sánchez, J. 2003. Aves del corredor biológi co entre los Parques Nacionales Naturales Puracé Cueva de los Guácharos. Guía de Campo. 358 p.

Notas
1 Estudio denominado caracterización florístico estructural de la vegetación del corredor biológico entre los PNN Puracé y Cueva de los Guácharos (2007) realizado mediante el financiamiento de la Corporación Autoónoma Regionaldel Alto Magdalena CAM.
Creation date:




Licencia
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






