
DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2008.1.a04Publicado:
01-01-2008Número:
Vol. 11 Núm. 1 (2008): Enero-DiciembreSección:
Artículos de investigación científica y tecnológicaCaracterización y Valorización del Potencial de Regeneración del Banco de Semillas Germinable de la Reserva Forestal Cárpatos (Guasca, Cundinamarca)
Characterizing and valuing of regeneration potential of the seed germination bank at the Carpatos forest reserve (Guasca, Cundinamarca)
Palabras clave:
Andean forest, seed bank, ecological restoration, phenology, scattering. (en).Palabras clave:
bosque andino, banco de semillas, restauración ecológica, fenología, dispersión (es).Descargas
Referencias
Acosta, M. 2004. Efecto sobre el banco de semillas germinable en un fragmento de bosque altoandino (páramo de Guerrero). Trabajo de grado. Universidad Nacional de Colombia. Pg. 89.
Bertiller, B. 1991. Seasonal Variation in the Seed Bank of a Patagonian Grassland in Relation to Grazing and Tography. Journal of Vegetation Science 2: 47-54 .
Bonilla, A. & G. Zuloaga. 1994. Fenología de algunas especies de plantas del páramo "El Granizo", Monserrate (Cundinamarca, Colombia). Academia Colombiana de Ciencias Exactas, Físicas y Naturales. Bogotá. Colección Jorge Álvarez Lleras 6: 485-501.
Brown, D. 1992. Estimating the Composition of a Forest Seed Bank: a Comparison of the Seed Extraction and Seedlink Emergence Methods. Canadian Journal of Botany 70: 1603-1612.
Cantillo, E., K. Rodríguez & A. Avella. 2004. Caracterización florística, estructural, diversidad y ordenación de la vegetación en la Reserva Forestal Cárpatos, Guasca, Cundinamarca. Universidad Distrital Francisco José De Caldas. Bogotá. Pg. 150.
Carabias, J. & S. Guevara. 1985. Fenología de una selva tropical húmeda y en una comunidad derivada: Los Tuxtlas, Veracruz. En: GómezPompa, A. & S. del Amo (eds.). Investigaciones sobre la regeneración de selvas altas en Veracruz, México. Volumen ii Págs. 27-78.
Cardona, A. 2004. Potencial de regeneración del banco de semillas germinable en dos tipos de bosque subandino: Implicaciones para la restauración (Reserva Biológica Cachalú, Encino, Santander). Trabajo de grado. Universidad Distrital francisco José de Caldas. Bogotá.
Conell, H. 1978. Diversity in Tropical Rain Forests and Coral Reefs. Science 1999: 1302-1310.
corpoguavio. 2000. Plan de Recuperación y Manejo Forestal, predio Cárpatos. Bogotá.
Croat, T. 1975. Phenological Behavior of Habit and Habitat Classes on Barro Colorado Island. Biotropical 7: 270-277.
Cuatrecasas, J. 1934. Observaciones geobotánicas en Colombia. Trabajos del Museo Nacional de Ciencias Naturales. Madrid. Serie botánica 27. Pg. 143.
Fagua, C. 2002. Estrategia de reproducción sexual de una población de E. grandiflora en el Parque Nacional Natural de Chingaza. Universidad Nacional de Colombia. Departamento de Biología. Bogotá.
Falinska, K. 1999. Seed Bank Dynamics in Abandoned Meadows during a Twenty Year Period in the Bialowieza National Park. Journal of Ecology 87: 461-475.
Fenner, M. 1995. Ecology of Seed Banks. En: Kigel, J.. Seed Develoment and Germination. Inc. New York. Págs. 507-527.
Fleming, T., R. Bridtwisch & G. Whitesides. 1987. Patterns of Tropical Vertebrate Frugivore Diversity. Annual Review of Ecology and Systematics 18: 91-108.
Fonseca, T. 2001. Dinámica de la dispersión de semillas por aves en un pastizal con perchas artificiales en comunidades de vegetación altoandina. Tesis de pregrado. Pontificia Universidad Javeriana. Bogotá.
Foster, S. & C. Janson. 1985. The Relationships between Seed Size and Establishment Conditions in Tropical Woody Plants. Ecology 66: 773-780.
Garwood, N. 1989. Tropical Soil Seed Banks. Ecology of Soil Seed Bank. Academic Press. San Diego, California. Págs. 149-209.
Gibson, D. & K. Brown. 1991. The Effects of Grazing on Local Colonization and Extinction during Early Succession. Journal of Vegetation Science 2: 291-300.
Grandin, U. & H. Rydin. 1998. Atributes of the Seed Bank after a Century of Primary Succession on Islands in Lake Hjälmaren, Sweden. Journal of Ecology 86: 293-333.
Grime, J. 1989. Seed Bank in Ecological Perspective. En: Leck, M. A., V. T. Parker y R. L. Simpson (eds.). Ecology of Soil Seed Bank. Academic Press. San Diego, California. Págs. 15-27.
Guevara, S. & A. Gómez-Pompa. 1972. Seed from Surface Soils in a Tropical Region of Veracruz, Mexico. Journal of the Arnold Arboretum 53 (3): 312-335.
Herrera, C. & P. Jordano. 1981. Prunus mahaleb and Birds: The HighEfficiency in the System of a Temperate Fruiting Tree. Ecological Monographs 51: 203-218.
Herrera, R., D. Ulloa, O. ValdésLafont,A. Priego, A., & Valdés, A. 1997. Ecotechnologies for the Sustainable Management of Tropical Diversity. Nature y Resources 33: 2-7.
Hilty, S. 1980. Flowering and Fruiting Periodicity in a Premontane Rainforest in Pacific Colombia. Biotropical 12: 292-306.
Howe, H. & R. Primack. 1975. Differential Seed Dispersal by Birds of the Tree Casearia nitida (Flacurtaceae). Biotropical 7 (4): 278-283.
Howe, H. & J. Smallwood. 1982. Ecology of Seed Dispersal. Annual Review of Ecology and Systematics 13: 201-208.
Jaimes, V. & D. Rivera. 1990. Banco de semillas y tendencias en la regeneración natural de un bosque altoandino en la región de Monserrate (Cundinamarca, Colombia). Trabajo de grado. Facultad de Biología. Universidad Nacional de Colombia. Bogotá. Pg. 56.
Janzen, D. 1983. Birds. En: Janzen, D. (ed). Costa Rica Natural History. University of Chicago Press. Págs. 502-549.
Jordano, P. & E. Schupp. 2000. Seed Dispersal Effectiveness: The Quantity Component and Patterns of Seed Rain for Prunus mahaleb. Ecological Monographs 70: 591-615.
Levey, D. 1988. Spatial and Temporal Variation in Costa Rican Fruit and Fruit-Eating Bird Abundance. Ecological Monographs 58: 251-269.
Magurran, A. E. 1989. Diversidad ecológica y su medición. Ediciones Vedra. Barcelona. Pg. 200.
Mahecha. G. 2004. La vegetación del territorio car. car (Cundinamarca). Bogotá. Pg. 620.
Marquis, R. 1988. Phenological Variation in the Neotropical Understory Shrub Piper arienum: Causes and Consequences. Ecology 69: 15521565.
Martínez-Ramos, N. & E. Álvarez-Buylla. 1986. Seed Dispersal, Gap Dynamics and Tree Recruitment: the Case of Cecropia obtusifolia at Los Tuxtlas, Mexico. En: Estrada, A. & H. Fleming. Frugivory and Seed Dispersal. Junk Publishers. La Haya. Págs. 333-346.
Mc Graw, J. & M. Varek. 1989. The Role of Buried Viable Seeds in Artic and Alpin Communities. En: Leck, M., V. Parker y L. Simpson (eds.). Ecology of Soil Seed Bank. Academic Press, San Diego, California. Págs. 91-105.
Mckey, D. 1975. The Ecology of Coevolved Seed Dispersal Systems. En: Gilbert, L. & P. Raven (eds.). Coevolution of Animals and Plants. University of Texas Press. Págs. 159-191.
Montenegro, A. 2000. Estrategias de dispersión y regeneración por banco de semillas en dos comunidades de bosque alto andino en Colombia. Trabajo de grado. Facultad de Biología. Universidad Nacional. Bogotá. Pg. 132.
Morales, N. 2002. Ecología de la dispersión de semillas por aves de Brunellia colombiana en un bosque altoandino (Parque Nacional Natural Chingaza). Tesis. Universidad Nacional de Colombia. Pg. 85.
Mori, S. & J. Kallunki. 1976. Phenology and Floral Biology of Gustavia superba (Lecythidaceae) in Central Panama. Biotropical 8: 184-192.
Murray, G. 1987. Selection for Optimal Fruit Crop Size in Bird-Dispersed Plants. American Naturalist 129: 18-31.
Newstrom, L., G. Frankie, H. Baker y R. Collwell. 1994. Diversity of Long-term Flowering Patterns. En: Mc Dade, K., H. Bawa, H. Hespenheide & G. Hartshorn (eds.). Ecology and Natural History of a Neotropical Rainforest.. Págs. 142-160.
Olaya, A. 2000. Efecto de borde sobre el banco de semillas germinable del bosque altoandino (Embalse de San Rafael, La Calera, Cundinamarca). Tesis de grado. Pontificia Universidad Javeriana. Bogotá.
Pedraza, L., 2000. Las plantas con flores de los alrededores de la laguna de Chisacá (Sumapaz, Colombia). Trabajo de grado en Biología. Universidad Nacional de Colombia. Facultad de Ciencias. Bogotá. Pg. 237.
Piquett, S. & M. McDonnell. 1989. Seed Bank Dynamics in Temperate Deciduous Forests. En: Allessio, M., V. Parker V & R. Simpson (eds.). Ecology of Soil Seed Banks. Academic Press. San Diego, California. Págs. 123-146.
Posada, C. & C. Cárdenas. 1999. Ecología de los bancos de semillas en una comunidad vegetal de páramo sometida a disturbios por quema y pastoreo (Parque Nacional Natural Chingaza). Trabajo de grado. Universidad Nacional de Colombia. Facultad de Biología. Bogotá. Pg. 190.
Poullin, B., J. Wright, G. Lefebvre & O. Calderón. 1999. Interspecific Synchrony and Asynchrony in the Fruiting Phenologies of Congereric Bird-Dispersed Plants in Panama. Journal of Tropical Ecology 15: 213-227.
Rice, J. 1989. Impacts of Seed Banks on Grassland Community Structure and Population Dynamics. En: Leck, M., V. Parker y R. Simpson (eds.). Ecology of Soil Seed Bank. Academic Press. San Diego, California. Pg. 211-230.
Rico-Gray, V. & J. García-Franco. 1992. Vegetation and Soil Seed Bank of Successional Stages in Tropical Lowland Deciduous Forests. Journal of Vegetation Science 3: 617-624.
Rossi, R, P. Lavelle & E. Tondof. 1995. Statistic Soil Biology. Geostatical Analysis. European Journal of Soil Biology 31 (4): 173-181.
Saldarriaga, J. G. 1994. Recuperación de la selva de tierra firme en el alto río Negro, Amazonia Colombiana-Venezolana. Tropenbos.. Pg. 201
Sánchez, M., 2003. Fenología reproductiva y dispersión de semillas del arbusto altoandino Monina salicifolia. Tesis de grado. Universidad Nacional de Colombia. Bogotá. Pg. 65.
Schupp, E., H. Howe, C. Augspurger & D. Levey. 1989. Arrival and Survival in Tropical Treefall Gaps. Ecology 70 (3): 562-564.
Society for Ecological Restoration (ser). 2004. ser International Primer on Ecological Restoration. Society for Ecological Restoration [disponible en: http://www.ser.org/content/ ecological_restoration_primer.asp].
Simpson, R., M. Allessio & V. Parker. 1989. Seed Bank: General Concepts and Methodological Issues. En: Leck, M., V. Parker y R. Simpson (eds.). Ecology of Soil Seed Bank. Academic Press. San Diego, California. Págs. 3-8.
Smythe, N. 1970. Relationships between Fruiting Seasons and Seed Dispersal Methods in a Neotropical Forest. American Naturalist 104: 25-35.
Sorzano, C. 2001. Regeneración reproductiva en un sitio abandonado e invadido por Pteridium aquilinum Khun. Implicaciones para la restauración. Reserva Biológica Cachalú. Encino, Santander, Colombia. Trabajo de grado. Pontificia Universidad Javeriana. Facultad de Ciencias. Departamento de Biología. Bogotá.
Swaing, D. & J. Hall. 1983. Early Succession on Cleared Forest Land in Ghana. Journal of Ecology 71: 601-627.
Thompson, K., J. Bakker & R. Bekker. 1998. Ecological Correlates of Seed Persistence in Soil in the North-West European Flora. Journal of Ecology 86: 163-169.
van Schaik, C. 1993. The Phenology of Tropical Forest: Adaptative, Significance and Consequences for Primary Consumers. Annual Review of Ecology and Systematics 24: 353- 377.
Vargas, O. 1998. Un modelo de sucesión/regeneración de los páramos después de las quemas. Caldasia 19 (1; 2): 331-345.
Wikander, T. 1984. Mecanismos de dispersión de diásporas de una selva decidua en Venezuela. Biotropical 16 (4): 276-283.
Williams-Linera, G. & y J. Meave. 2002. Patrones fenológicos. En: Guariguata, M. y G. Kattan (eds.). Ecología y conservación de bosques neotropicales. Libro universitario regional de Costa Rica. Págs. 407-431.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
 |
Cantillo-Higuera, E.E., Castiblanco-Gutiérrez, V., Pinilla-Mondragón, D.F., & Alvarado, C.L. (2008). Caracterización y valoración del potencial de regeneración del banco de semillas germinable de la reserva forestal Cárpatos (Guasca, Cundinamarca). Colombia Forestal, 11, 45-70. |
CARACTERIZACIÓN Y VALORACIÓN DEL POTENCIAL DE REGENERACIÓN DEL BANCO DE SEMILLAS GERMINABLE DE LA RESERVA FORESTAL CÁRPATOS (GUASCA, CUNDINAMARCA)1
Characterizing and valuing of regeneration potential of the seed germination bank at the Carpatos forest reserve (Guasca, Cundinamarca)
Caracterização e avaliação do potencial de regeneração do banco de sementes germináveis da reserva florestal Cárpatos
Edgard Ernesto Cantillo Higuera2, Viviana Castiblanco Gutiérrez3, David Fernando Pinilla Mondragón4 & Claudia Liliana Alvarado5
1Resumen de los resultados del Proyecto de Investigación financiado por el Centro de
Investigaciones de la Universidad Distrital Francisco Joséde Caldas.
2Universidad Distrital Francisco José de Caldas, Bogotá, Colombia,
ecantillo@udsitrital.edu.co.
3Universidad Distrital Francisco José de Caldas, Bogotá, Colombia,
vicasgforestal@yahoo.es.
4Universidad Distrital Francisco José de Caldas, Bogotá, Colombia,
davidfpinillaforestal@gmail.com.
5Universidad Distrital Francisco José de Caldas, Bogotá, Colombia,
lilianalvar70@hotmail.com.
Recepción: Septiembre 10 de 2008/Aprobación: Octubre 31 de 2008
RESUMEN
En la Reserva Forestal Cárpatos se caracterizó y valoró el bancode semillas germinable (BNG) y se definieron los mecanismos y estrategias de dispersión de la reserva, que posteriormente determinaron la estructura espacial, temporal y dinamogenética de los estados serales, así como de sus poblaciones. La vegetación en pie registró 61 especies, representadas por 53 géneros y 36 familias; paralelamente en el BSG se identificaron 55 especies, con de 50 géneros y 27 familias. El volumen mínimo de la unidad de muestreo fue de 1.500 cm3, con un total de 13.150 semillas germinadas, mientras que la densidad total para el BSG fue de 1.553 semillas por m2. Las especies emergidas en el BSG estuvieron constituidas en su mayoría de porte herbáceo y rasante –aproximadamente 70% del total de individuos y de especies–; Phytolacca bogotensis, Digitalis purpurea y Ageratina sp. fueron las especies dominantes en los tres ensayos del banco. Otras especies de importancia fueron Carex sp., Poa annua, Rubus floribundum, Bulbostylis sp. y Borreria sp. Las especies de porte leñoso variaron en cada ensayo de acuerdo a la fenofase de cada una de ellas; las más representativas fueron Cestrum mutisi, Clusia ducu, Myrsine coriacea, Axinaea macrophylla y Miconia denticulata.
En cuanto afenología y dispersión, se encontró que la vegetación en pie presentó a la zoocoria como la estrategia de dispersión dominante en todas las parcelas, los máximos de floración se presentaron durante la estación seca, entre diciembre y marzo, mientras que los máximos registros de fructificación correspondieron al periodo de la estación de lluvias más prolongada, entre junio y agosto.
Palabras clave: bosque andino, banco de semillas, restauración ecológica, fenología, dispersión.
ABSTRACT
In the Cárpatos Forest Reserve the bank seed germinate (BSG) was marked and valued and the dispersion mechanisms and strategies of there serve defined. They subsequently determined the spatial, dinamogeneticand temporary structures of the seral states and their populations. For the vegetation on foot 61 species were recorded, represented by 53 genera and 36 families, alongside the BSG were 55 species, represented by 50 genera and 27 families. The minimum volume was 1.500 cm3, for a total of 13.150 germinating seeds; the total density for BSG was 1.553 seeds per m2. The species emerged in the BSG were mostly herbaceous and flush – approximately 70% of all individuals and species –; Phytolacca bogotensis, Digitalis purpurea and Ageratina sp. were the dominant species in the three trials Bank, other important species were< I>Carex sp., Poa annua, Rubus floribundum, Bulbostylis sp.and Borreria sp. The species of woody habit usually varied in each trial according to the phenophase of each species, the most representative species were Cestrum Mutisia, Clusia ducu, Myrsine coriacea, Axinaea macrophylla and Miconia denticulata.
As for phenology and dispersal, it was found that vegetation stands submitted to the zoocoria as the dominant scattering strategy in all fields; the maximum flowering occurred during the dry season, from December to March, though the greatest fruiting records correspond to the longest rainy season, from June to August.
Key Words: Andean forest, seed bank, ecological restoration, phenology, scattering.
RESUMO
Na Reserva Florestal Cárpatos se caracterizou e avaliou o banco de sementes germináveis (bsg) e se definiram os mecanismos e estratégias de dispersão da reserva, que posteriormente determinou a estrutura espacial, temporal e dinamogenética dos estados existentes, assim como de suas populações. A vegetação em pé registrou 61 espécies, representadas por 53 gêneros e 36 famílias; paralelamente no bsg se identificaram 55 espécies, com 50 géneros e 27 famílias. O volume mínimo da unidade de mostras foi de 1.500 cm3, com um total de 13.150 sementes germinadas, enquanto que a densidade total para o bsg foi de 1.553 sementes por m2. As espécies emergidas no bsg estavam constituídas, na sua maioria, de porte herbáceo e rasante - aproximadamente 70% do total de individuos e de espécies – Phytolacca bogotensis, Digitalis purpúrea e Ageratina sp. foram as espécies dominantes nos três ensaios do banco. Outras espécies de importância foram Carex sp., Poa annua, Rubus floribundum, Bulbostylis sp. e Borreria sp. As espécies de porte lenhoso variaram em cada ensaio de acordo com a fenofase de cada uma delas; as mais representativas foram Cestrum mutisi, Clusia ducu, Myrsine coriacea, Axinaea macrophylla e Miconia denticulata. Em relação à fenología e a dispersão, se constatou-se que a vegetação em pé apresentou a zoocoria como estratégia de dispersão dominante em todos os seus pedaços de terra. O máximo de floração se apresentou durante a estação seca, entre dezembro e março, enquanto que os registros máximos de frutificação corresponderam ao período da estação de chuvas mais prolongada, entre junho e agosto.
Palavras chave: Bosque andino, banco de sementes, restauração ecológica, fenologia, dispersão.
INTRODUCCIÓN
Los ecosistemas altoandinos y de páramo se encuentran actualmente sometidos a una gran transformación generada por la expansión de la frontera agrícola y ganadera. Ante la urgente necesidad de restablecer los ecosistemas andinos,larestauración ecológica, definida como el proceso asistido de recuperación de un ecosistema que ha sido degradado, dañado o destruido (SER 2004), se convierte en el mecanismo que permite obtener el restablecimiento de los atributos estructurales y funcionales de los ecosistemas deteriorados. Así, la recuperación de estos ecosistemas dependerá del “potencial de regeneración in situ”, es decir, de los remanentes bióticos supervivientes a la perturbación (Uhl & Saldarriaga 1986, citados por Saldarriaga 1994). Como parte de este potencial biótico, el banco de semillas germinable y la lluvia de semillas representan las principales estrategias que permiten que un individuo se reemplace por otro en las siguientes generaciones(Sorzano 2001). El banco de semillas germinable (BNG) tiene la tarea de mantener una reserva natural de diversidad genética o memoria genética de los efectos selectivos deambientes pasados(McGraw & Vavrek1989); éste se relaciona de manera directa con las fases del ciclo de vida de las plantas o fenofases, que incluyen el crecimiento de yemas florales, la apertura floral, la producción de frutos inmaduros y la maduración de los mismos (van Schaik 1993), convirtiéndose en un requisito indispensable para la restauración de los ecosistemas (Newstrow et al. 1994). La dispersión de semillas, considerada como el último paso del ciclo reproductivo de las plantas, es el proceso mediante el cual las semillas son transportadas lejos del parental aunsitio favorable para su crecimiento y desarrollo (Herrera et al. 1997). El proceso de dispersión permite la colonización de nuevos hábitats, reduce la competencia intraespecífica y facilita los procesos de germinación (Morales 2002). En esta contribución se logró caracterizar y valorar el potencial de regeneración del BSG en la Reserva Forestal Cárpatos; nuestros objetivos fueron: (1) Evaluar la diversidad de las especies presentes en el BSG para los diferentes estados serales de la reserva.(2) Establecersiexisteestacionabilidadpara las fenofases de producción y los tipos de dispersión, y determinar la incidencia de éstos en el BSG. (3)Determinar las principales formas de dispersión de las semillas e identificar cuáles tipos de especies son las mayores aportantes para la preservación de la dinámica del bosque. (4) Cuantificar el aporte de la avifauna a los procesos de dispersión de semillas en claros de bosque secundario. (5) Evaluar las diferencias en cuanto a especie, número y germinabilidad de las plántulas producidas por BSG, en diferentes periodos del año.
METODOLOGÍA
DESCRIPCIÓN DE LA ZONA DE ESTUDIO
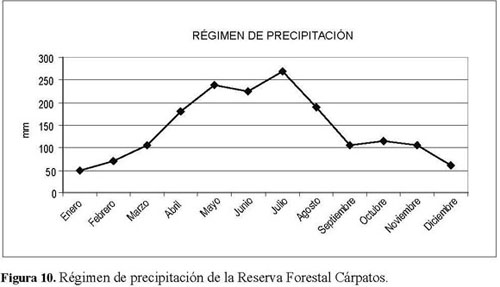
La Reserva Forestal Cárpatos hace parte del bosque altoandino y está localizada en el costado noroccidental de la jurisdicción de la Corporación Autónoma Regional del Guavio en el municipio de Guasca, Cundinamarca, a 97 km de Bogotá (Figura 1), en la vereda Concepción, cerca del sitio denominado Potreritos, ubicado en el extremo oriental delmunicipiomencionado,enellímitesurdelmunicipio Guatavita y al occidente de municipio de Junín. Altitudinalmente se halla entre los 2.600 y 3.000 msnm. Se localiza geográficamente en las coordenadas: latitud Norte 4º 47 minutos, longitud Oeste 73º 40minutos; su topografía es montañosa, destacándose como accidentes orográficos los cerros de Los Robles. Por su ubicación, la estación Potreritos es la que mejor tipifica la precipitación en la reserva. La precipitación promedio anual es de 1.714.3 mm y la media mensual es de 142.9 mm. El régimen pluviométrico es monomodalbiestacional, concentrándose las mayores lluvias entre abril y agosto, periodo en el cual los vientos alisios provienen del Este, pudiéndose inferir que éstos vienen cargados de humedad y, por efectos orográficos, descargan parte de ésta en la zona de estudio. El periodo seco se presenta entre septiembre y marzo y el mes más seco corresponde a enero, con 47 mm de precipitación. Sus tierras son influenciadas por los ríos Guavio y Sueva, entre otros (Cantillo et al. 2004). En general, son suelos jóvenes, con influencia de ceniza volcánica en sus primeros horizontes. El relieve ha influido en la dispersión y acumulación de las cenizas y constituye un factor importante para la susceptibilidad a la erosión. Se encontraron básicamente dos tipos de suelos: Typic Humitropept y Typic Dystropept, que son moderadamente profundos a superficiales, bien drenados, de alta acidez, friables, con bajos contenidos de carbono, de una fertilidad mediana a baja (Corpoguavio 2000).
FASE DE CAMPO
Composición de la vegetación
Se establecieron tres parcelas de 50 x 50 m, cada una en un estado de desarrollo sucesional de la reserva así: bosque secundario clímax, borde de bosque y borde de bosque empatado con claros. En cada parcela se realizó un inventario de los individuos más representativos en cuanto a mayor talla, área basal y cobertura de cada una de las especies presentes, identificadas por Cantillo et al. (2004).
Banco de semillas germinable
Premuestreo (determinación del volumen mínimo). El tamaño óptimo de la unidad de muestreo fue determinado a través de una curva de especies (Simpson et al. 1989);en los bosques alto andinos, de acuerdo a lo propuesto por Jaimes y Rivera(1990),la unidad muestral se calculó apartir de una curva especies vs. volumen; los volúmenes se extrajeron a lo largo de la reserva con unidades muestrales de 1.000, 1.500, , 2.500 y 3.000 cm3. Cada volumen contó con tres repeticiones y aquellos que presentaron los mayores valores de especies por volumen se definieron como el volumen mínimo a muestrear.
Muestreo. Cada parcela de 50 x 50 m fue subdividida en cuadriculas de 10 x 10 m. Se tomó una muestra de suelo en cada uno de los 36 puntos de intercepción de la parcela, a una profundidad de 15 cm. Al momento de tomar las muestras se retiró la cobertura vegetal, así como la hojarasca, briofitos y hepáticas. Cada muestra se depositó en bolsas plásticas que se etiquetaron, se sellaron y se trasportaron al invernadero. Para cada muestra se tomó una replica a una distancia aproximada de un metro. En total se tomaron 72 muestras de suelo por cada parcela y 216 para cada ensayo. Se realizaron tres muestreos con intervalo de cuatro meses entre cada uno, para un total de 648 muestras, con un tiempo total empleado de un año.
Fenología y dispersión de semillas
Este estudio se realizó en dos tipos de cobertura vegetal: el primero dentro del bosque, con el registro de las fenofases a partir del registro quincenal de trampas de vuelo y la determinación de los principales síndromes de dispersión de semillas de la vegetación superior a partir de observación directa y referencias bibliográficas, para la vegetación en pieyel BNG; el segundo en claros de bosque secundario por medio de la instalación de perchas artificiales para determinar la incidencia de la avifauna en la dispersión local de semillas; la instalación de las perchas se realizó a diferentes distancias del borde del bosque (cercanía, medianía y lejanía); el número de perchas por ensayo fue de diez y cada uno tuvo una réplica, para un total de 60 perchas. El monitoreo en ambos casos, tanto en fenología como en la dispersión de semillas, se realizó cada dos semanas durante un periodo de un año.
FASE DE ANÁLISIS
Vegetación en pie
Se construyeron gráficas de todas las variables tomadas para determinar el comportamiento de la vegetación en los diferentes estados serales y la dinámica de los hábitos dentro de cada uno de ellos. Paralelamente, se elaboraron gráficas, por estrato, delcomportamientodelacoberturadelasespecies para las tres parcelas.
Banco de semillas germinable
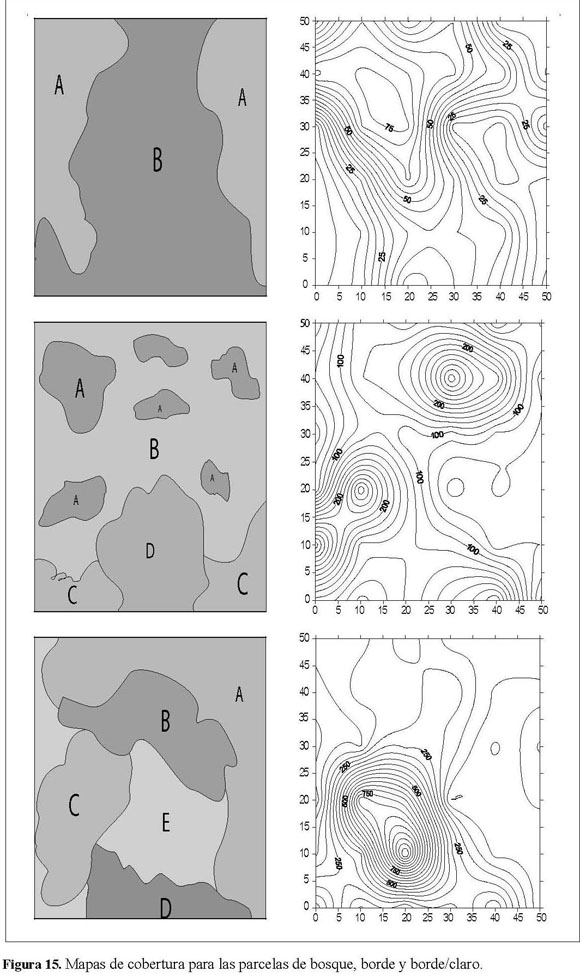
Se realizó un análisis de varianza multifactorial por medio del programa Statgraphics Plus 5.1, con el fin de determinar diferencias espacio-temporales del BSG. Adicionalmente, se efectuó el análisis geoestadístico por medio de la generación y posterior relación de variogramas de correlación espacial a través de los programas Geo-EAS 1.2.1 y Surfer32 v8.Estos variogramas ayudan a determinar si las variables evaluadas presentan algún tipo de estructura espacial. De la misma manera, se crearon mapas de distribución para cada parcela, según los datos de abundancia obtenidos en el BSG, por medio del programa Surfer32 v8; estos programas fueron utilizados por Montenegro (2000) yAcosta (2004) en sus respectivos trabajos sobre BSG. La realización tanto de los variogramas como de los mapas de superficie se efectuó con la suma de los datos de los tres muestreos para cada punto muestral; de esta manera, aquellos puntos donde se presentaron mayores abundancias son resaltados por medio de picos que definen tendencias agrupadas; análogamente, se eliminaron datos gregarios poco representativos del ensayo en general (Rossi et al. 1995, Montenegro 2000). Para analizar la dominancia y la diversidad de las especies del banco de semillas se construyeron modelos de acuerdo a lo sugerido por Magurran (1989). Estos modelos han sido producidos por diversos estudios de bancos de semillas en el país, entre los que se destacan los de Posada & Cárdenas (1999), Montenegro(2000) yAcosta(2004). Se llevó a cabo la prueba de diferencia significativa para establecer cuáles parcelas presentaban mayor similitud(Montenegro,2000).
Fenologia y dispersión
En las trampas y perchas se utilizaron pruebas de concordancia de Kendall para este tipo de distribución; los análisis se realizaron por medio del programa SPSS 7.5 para Windows y Statistica, de acuerdo a lo sugerido y realizado por Morales (2002) y Sánchez (2003).
RESULTADOS
COMPOSICIÓN DE LA VEGETACIÓN
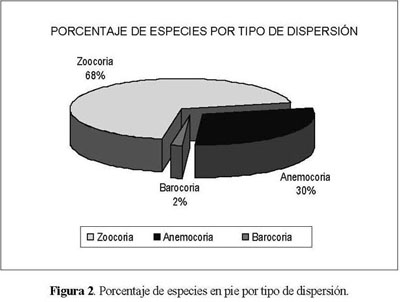
En total, para la vegetación en pie de las tres parcelas se encontraron 61 especies, representadas por 53 géneros y 36 familias. Las familias más representativas fueron: Lauraceae (11%),Melastomataceae (10%), Compositae (7%) y Myrsinaceae (5%) (Anexo 1). La zoocoria se presentó como el síndrome de dispersión dominante con 68%, seguida de laanemocoriaconel30%ylabarocoriaconel2% de representatividad (Figura 2).
El número de familias en las parcelas aumenta en forma proporcional con respecto a la disminución en el desarrollo de las etapas séricas (Figura 3), ya que mientras la parcela de bosque clímax contiene el 54% de los registros, la parcela de borde con claro contiene el 70% de las familias.
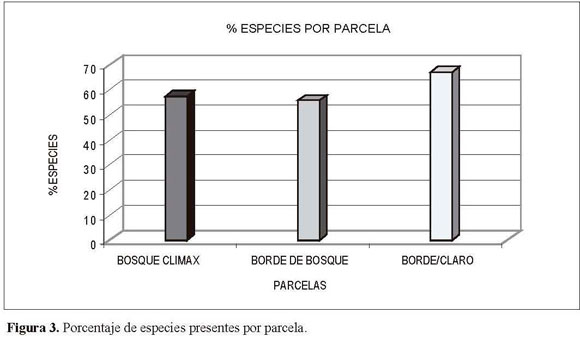
La parcela de bosque clímax registró 35 especies de 28 géneros y 20 familias; por su parte, en la parcela de borde se encontraron 34 especies de 30 géneros y 23 familias, mientras que en la parcela de borde de bosque con claro se hallaron 41 especies de 37 géneros y 26 familias.
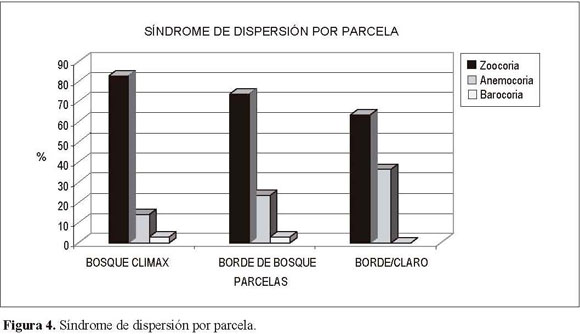
La relación de las tres parcelas por tipo de dispersión (Figura 4) mostró la zoocoria como el síndromedominanteparalastresparcelas,variandoentre el 60% (borde con claro) y 80% (bosque clímax); como segunda estrategia se presenta la anemocoria con una fluctuación entree l35% (bordeconclaro) y 15% (bosque clímax), seguida por la barocoria, la cual no es muy frecuente y presenta un valor bajo (<3%) endonde participa.
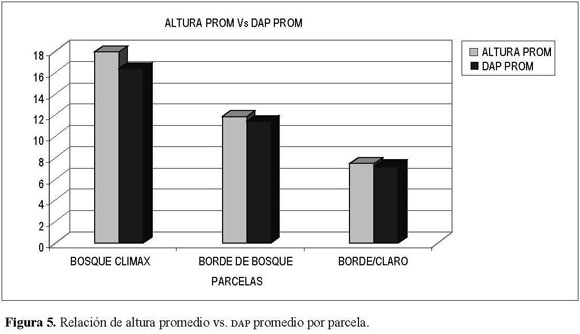
Entre el DAP promedio por parcela y la altura promedio por parcela (Figura 5) se presenta una relación directamente proporcional, al presentar un incremento en susregistros a medidaque lacomunidad se torna más madura.
BANCO DE SEMILLAS GERMINABLE (BSG)
Premuestreo
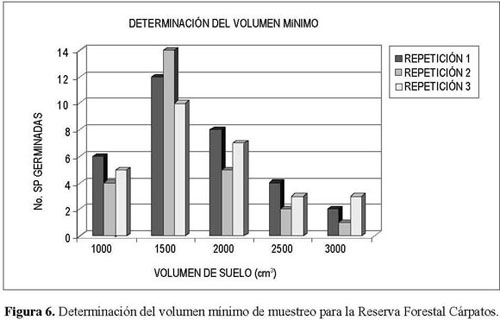
Los mayores registros de número de especies se presentaron en las muestras de 1.500 cm3, por lo que éste se definió como el volumen mínimo para la reserva (Figura 6).
Muestreos
En total a partir de los tres muestreos realizados a lo largo de un año germinaron 13.150 semillas pertenecientes a 55 especies de 50 géneros y 27 familias; del total de especies sólo ocho, que equivalen al 2.34% del total de los individuos, se identificaron hasta familia; la relación detallada de las especies se encuentra en el anexo 2. La densidad total del BSG fue de1.529 semillas/m2.
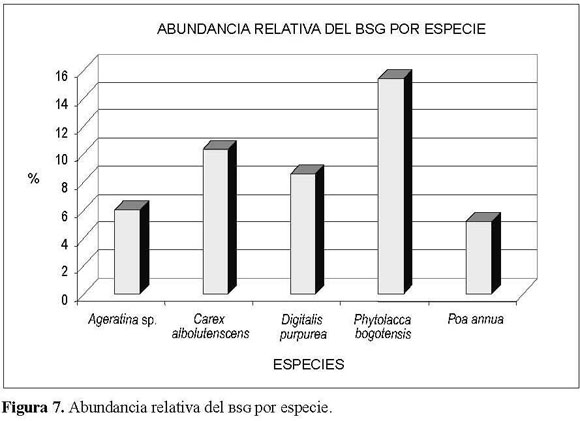
Las especies más abundantes para el banco de semillas de acuerdo con la figura 7 fueron en su or-den Phytolacca bogotensis, Carex albolutenscens, Digitalis purpurea, Borreria sp. y Ageratina sp., con valores entre el 15% y el 6% de la abundancia relativa total, es decir, entre 2.024 y 789 semillas germinadas.
Para el caso de las especies arbóreas, las más representativas fueron Cestrum mutisi, Buddleja bullata, Solanum inopinum, Axinaea macrophylla y Miconia denticulata, entre otras; estas especies individualmente presentan un porcentaje poco significativo, pues todas están por debajo de l2.2% del total de la abundancia relativa.
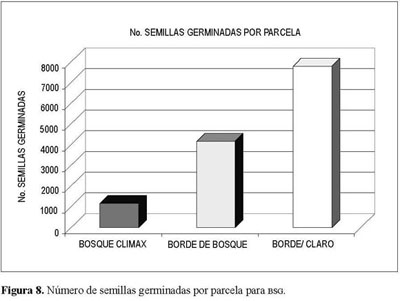
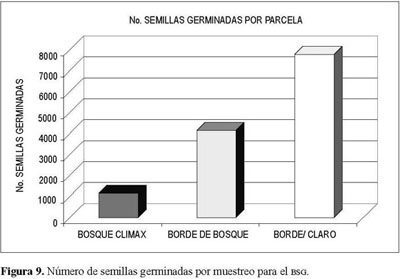
Los total es de semillas germinadas por parcela presentan valores disímiles, que aumentan conforme a una mayor heterogeneidad presente en la cobertura vegetal (Figura 8). Este comportamiento parece indicar que entre más desarrollo sucesional del ecosistema, mayores la posibilidad de aporte tanto del número como de la variabilidad de especies de semillas ya sea por lluvia o por estrategia de dispersión desdelugares aledaños.
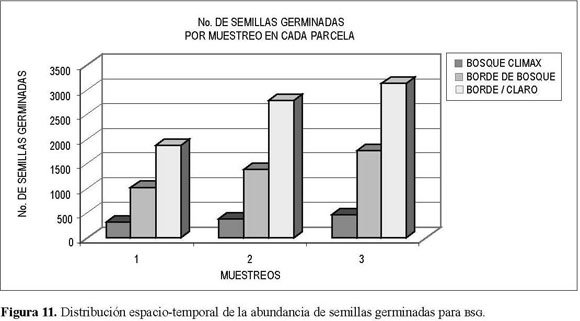
El comportamiento de muestreos varió, se registraron los menores valores para el muestreo I (noviembre-diciembre), con 3.214 semillas germinadas, hasta 5.378 semillas en el muestreo III (junio-julio, Figura 9).
No obstante la variabilidad del aporte de semillas entre los muestreos, el incremento en el número de semillas germinadas en las parcelas fue en igual proporción. Al situar cronológicamente los muestreos y compararlos con el régimen de precipitación de la reserva, se encontró una relación directamente proporcional entre la precipitación y el número de semillas inventariadas, teniendo en cuenta que el primer muestreo fue realizado a finales de noviembre, el segundo en marzo y el tercero en julio de (Figura 10 y 11).
En el anexo 3 se indican de manera resumida los valores de los índices de Margalef & Shannon hallados por muestreo y por parcela; a partir de ellos se pueden deducir los modelos de riqueza y abundancia del BSG de acuerdo a lo sugerido por Magurran (1989). Singularmente, los mayores registros tanto en riqueza y diversidad lo obtuvieron las parcelas de borde, debido posiblemente a la heterogeneidad de la composición de las coberturas boscosas en estos lugares.
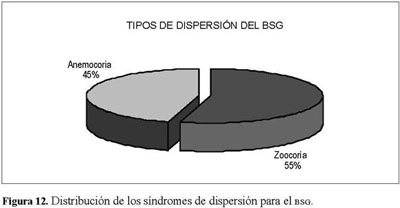
La tipificación del BSG presentó a la zoocoria como estrategia dominante de dispersión con un 55% del total de las especies, mientras que el restante 45% lo constituyó la anemocoria (Figura 12). No se presentó ninguna especie con estrategia barocoria.
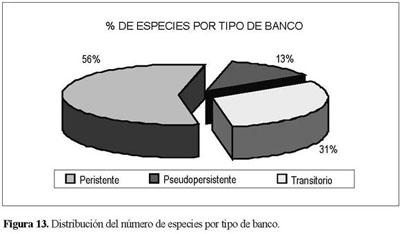
La distribución del tipo de semillas para el BSG registró el 84% por parte de especies que presentan semillas dicotiledóneas, frente a un 16% que constituyen las especies con semilla monocotiledóneas. Con respecto al tipo de semilla la relación fue de 3:1 (9.500 semillas dicotiledóneas frente a sólo 3.500 semillas de un monocotiledón). El tipo de banco para la reserva presenta una tendencia permanente al contener más de la mitad del total de las especies presentes del ensayo; los tipos de banco transitorio y pseudopersistente presentaron valores menores, aunque contaron con una participación significativa al tener el 31% y el 13%, respectivamente, del total de las especiesregistradas(Figura 13).
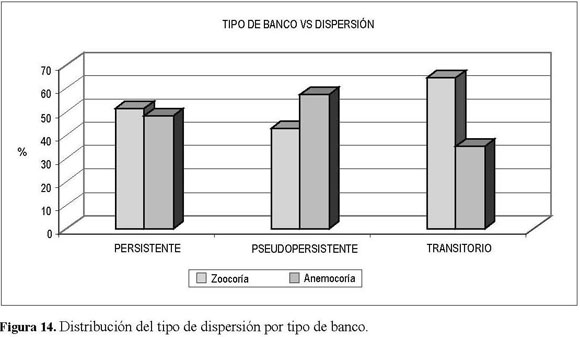
La zoocoria se presenta como la estrategia dominante en el aporte y lluvia de semillas (Figura 14), tanto de manera espacial (local y regional) como temporal (banco persistente y transitorio).
El análisis multifactorial del comportamiento espacio-temporal del BSG mostró que no hubo diferencias significativas entre parcelas y muestreos, pero sí existió diferencia entre la composición muestral de cada una de las parcelas (f=0.0005;g.l.=2;P=0.05).
Aunque la variación para cadaparcela fue diferente en la composición y número de especies, todas mostraron la misma tendencia de incremento por muestreo; además todas presentaron los mayores registros de emergencia de plántulas en los puntos donde la cobertura era más heterogénea (Figura 15). Por medio de la prueba de diferencia significativa se estableció que las parcelas que se relacionan con el borde presentan mayor asociación que las del bosque maduro.
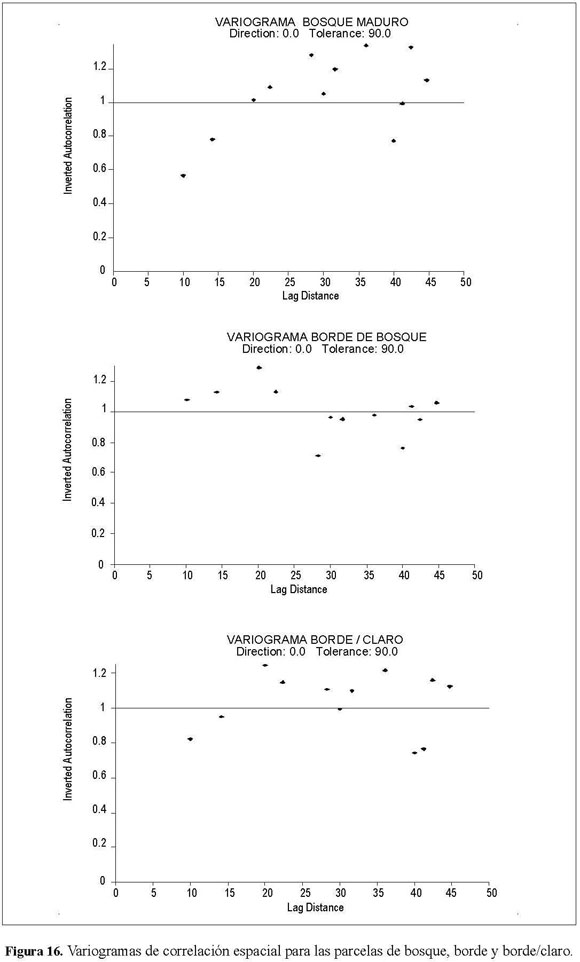
El comportamiento entre las unidades muestrales de las parcelas fue diferente, tanto de manera espacial como temporal; así, al generar los variogramas de correlación espacial se presentan diferencias en la estructura espacial de los puntos muestrales para cada parcela (Figura 16).
Igualmente, los mapas de superficie efectuados a partir de los datos de abundancia obtenidos en el BSG presentaron un comportamiento disímil entre las parcelas, variando así el número, la disponibilidad y la dispersión de las plántulas emergentes para cada una de ellas (Figura 17).
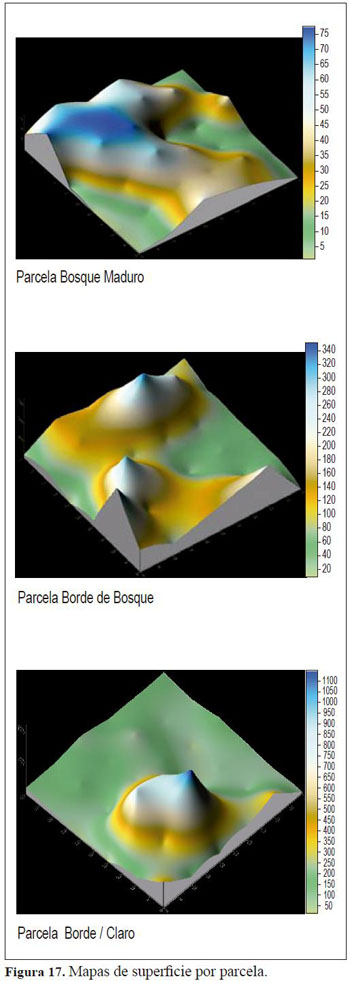
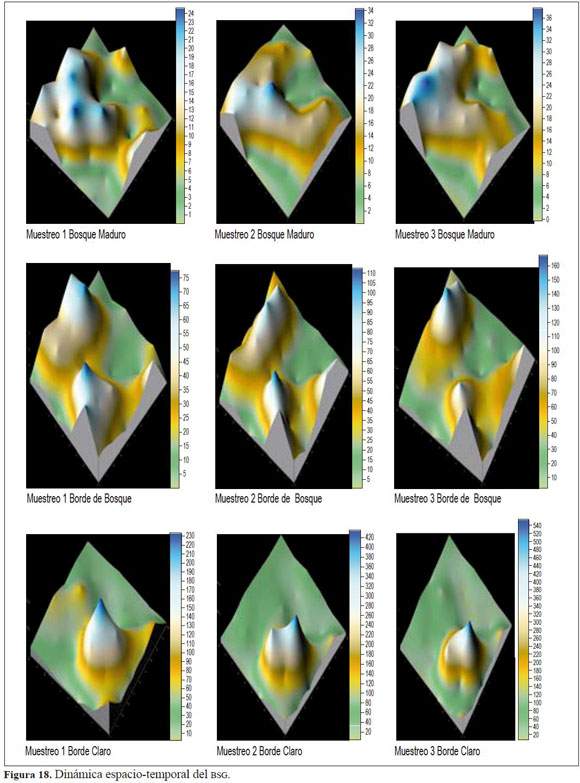
La variación en la composición de los registros fue disímil con respecto a los muestreos por lo que estructuralmente existe una dinámica espacio-temporal en la germinación de semillas (Figura 18), a pesar de que las muestras fueron tomadas en el mismo lugar (parcela) y tiempo (muestreo)
.
FENOLOGÍA Y DISPERSIÓN
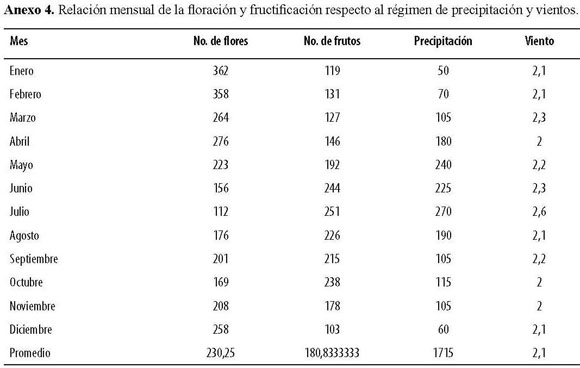
Los periodos de fenofases para la reserva parecen tener una relación estrecha con el régimen local de precipitación (Anexo 4); los máximos de floración se presentaron durante la estación seca (diciembre a marzo), mientras que los registros de fructificación correspondieron al periodo de la estación de lluvias más prolongada (entre los meses de junio a agosto), independientemente entre meses la floración y fructificación guardaron una relación inversamente proporcional en cuanto a la sincronía de sus producciones (W =0.028; X2 =0.333;n =12).
La duración de las fenofases entre especies también varió, pues mientras especies de comunidades maduras(Hedyosmon bomplandianum, Clusia ducu y Ocotea callophylla, entre otras) presentaron ciclos cortos y específicos de reproducción, las especies que corresponden a estados sucesionales iniciales como Phytolacca bogotensis correspondieron a periodos de floración y fructificación constantes durante el ciclo anual.
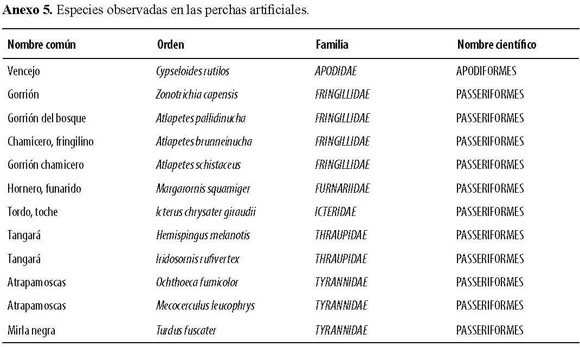
Las perchas artificiales fueron exitosas ya que durante todo el tiempo del ensayo contaron con visitas en mayor o menor proporción. El mayor registro de número de perchas visitadas ocurrió durante la época seca, donde los recursos son más limitados; el número de semillas, número de especiesdispersadas yvisitasa lasperchas presentaron sus máximos registros durante la época de lluvias. Se identificaron doce especies que fueron dispersadasdirectamentedesdelasperchas,entrelasque se destacan Rubus floribundum, Arcytophyllum sp. y Phytolacca bogotensis; de la misma manera, se identificarontrece especies de aves (Anexo 5).
DISCUSIÓN DE RESULTADOS
COMPOSICIÓN DE LA VEGETACIÓN
Especies como Myrsine coriacea, Miconia squamulosa, Drymis granadensis, Clusia multiflora, Weinmannia pinnata y Myrsine guianesis, pertenecientes a comunidades maduras, fueron también registradas por Cantillo et al. (2004) en el área de estudio y por Jaimes & Rivera (1990) y Montenegro (2000) en estudios realizados a la misma altitud; de la misma manera, especies asociadas con las primeras etapas de desarrollo, tales como Ageratina sp., Carex sp., Poa sp., Ruagea sp., Arcytophyllum sp. y Chusquea scandens forman parte de la vegetación de los ecosistemas altoandinos registrados por Posada & Cárdenas (1999), Montenegro (2000), Olaya (2000), Pedraza (2000), Acosta (2004) y Mahecha (2004), entre otros autores.
La menor variabilidad de hábitos y estratos se presentó en la parcela de bosque maduro, donde cerca del 95% del total de las especies correspondió a los estratos superiores, mientras que una pequeña minoría se refirió a hierbas (Ageratina sp.) y helechos (Cyathea sp.). Por su parte, la parcela que tuvo mayor número de hábitos correspondió al borde de bosque, al contar con ocho de los nueve tipos de vida registrados: arbóreo(68%) y arbustivo(18%), que fueron los más representativos. La distribución de abundancia, tanto para la vegetación como para el BNG, fue mayor en número de especies y semillas que la registrada por Montenegro (2000), Olaya (2000) y Acosta (2004). Esta distribución parece ajustarse al modelo normal, ya que, de acuerdo a Magurran (1989), en ninguna comunidad todas las especies pueden ser igualmente abundantes. La vegetación en pie tuvo a la zoocoria como el síndrome de dispersión dominante, propia de vegetación de hábitats húmedos, concordando con lo publicado por Howe & Smallwood (1982), que sitúan a esta estrategia en hábitats húmedos, mientras que las estrategias de dispersión abiótica como la anemocoria se presentan en agrupamientos marcados geográficay temporalmente de carácterseco.
BANCO DE SEMILLAS GERMINABLE
Volumen mínimo y densidad del banco
La determinación del volumen mínimo presentó un valor menor (1500 cm3) al determinado por Jaimes & Rivera (1990), que correspondía a 2000 cm3 y que se constituía como la unidad muestral ideal para estudios de BSG de los bosques altoandinos. Por medio del hallazgo de un nuevo volumen mínimo se puede afirmar que los ecosistemas alto andinos no presentan un patrón especifico, existiendo un alto grado de heterogeneidad en sus componentes (unidad muestral, densidad, abundancia y diversidad), como lo han demostrado estudios anteriores entre los que se destacan los de Posada & Cárdenas (1999), Montenegro (2000) y Acosta (2004). La densidad total del banco de semillas germinable fue de 1529 semillas/m2, valor menor a los determinados en varios estudios en ecosistemas altoandinos, como es el caso de Jaimes & Rivera (1990) con 1.813 semillas/m2 y el de Montenegro (2000) con 3.954 semillas/m2; sin embargo, el valor resultó ser mayor que el encontrado porAcosta (2004) en la reserva forestal de Cogua, el cual fue de 1.397 semillas/m2. Este valor está en el intervalo de 1.000 a 6.000 semillas /m2 que Guevara y Gómez-Pompa (1972) atribuyen a bosques secundarios parael trópico.
Dinámica de la emergencia de plántulas en el BSG
Phytolacca bogotensis, Digitalis purpurea, Ageratina sp., Carex sp., Cestrum mutisi, Buddleja bullata y Solanum inopinum fueron las especies primordiales que conforman el BSG en la reserva, especies que se cuentan dentro de los estudios de Posada y Cárdenas (1999), Montenegro (2000), Olaya (2000) y Acosta(2004). Las familias de mayor relevancia en cuanto a abundancia fueron: Cyperaceae, Phytolaccaceae y Compositae, mientras que las familias que registraron mayor número de especies fueron Compositae, Rubiaceae y Melastomataceae. De igual manera que en la vegetación en pie, la participación de los estratos arbóreo y arbustivo fue importante al contener cerca del 40% del total; sin embargo el hábito hierba fue el más dominante para el banco al contener el 43% de las especies. Este comportamiento concuerda con lo publicado por Conell (1978), que afirma que el disturbio promueve el dominio y la dispersión de especies pertenecientes a las etapas sucesionales tempranas. Los registros más altos de la abundancia de semillas emergentes coincidieron con las fluctuaciones temporales de la precipitación; esta característica puede estar relacionada con el even-to de que después del periodo lluvioso, la mayoría de semillas ya han sido dispersadas y se encuentran en el suelo prestas a germinar, coincidiendo con lo afirmado por Bonilla & Zuloaga (1994), que registran los máximos de fructificación en épocas secas y los de emergencia durante el periodo de lluvias. La diversidad y riqueza fueron mayores en las parcelas donde hubo mayor heterogeneidad de especies, de la misma manera ocurrió con los muestreos; esta relación de variación espacio-temporal fue también identificada en las parcelas realizadas por Montenegro (2000). El tipo de semilla dominantefueladicotiledónea,coincidiendoconloque afirman McGraw & Varek (1989): que las semillas monocotiledóneas son en proporción menos abundantes que las dicotiledóneas. Esta tendencia concuerda con lo expuesto por Rice (1989), quien afirma que cualquier grupo de hierbas dicotiledóneas son más dadas a formar bancos de semillas que los pastos.
Persistencia y tipo de dispersión del BSG
El banco de semillas presentó un aporte constante durante todo el año, por lo que, de acuerdo con lo publicado por Garwood (1989), se puede afirmar que la tasa de renovación de las semillas en el banco es igual o menor que la tasa de mortalidad, tendencia que permite la persistencia temporal del banco de semillas. Los resultados arrojados en el presente trabajo concuerdan con lo determinado por Montenegro (2000), al encontrar que el BSG está compuesto principalmente por semillas persistentes y pseudopersistentes, y muy pocas de tipo transitorio, ya que, de acuerdo con Thompson et al. (1998), los lugares con presencia de disturbios presentan bancos con tendencia a la persistencia temporal. La reserva muestra una tendencia a presentar un BSG constanteconlamayoríadelasespecies y el 89% del total de la abundancia del banco; los tipos de banco transitorio y pseudopersistente tuvieron una participación significativa en especies mas no en número de emergencias. Una posible causa de la persistencia puede deberse a que elbancopresentaenlamayoríadesucomposición a especies herbáceas, concordando con lo hallado con Montenegro (2000) yAcosta (2004). Este tipo de hábito tiende a la persistencia, ya que germinan por pulsos, característica que les ayuda a perdurar tras disturbios (Bertiller 1991, Grime 1989). Otras variables que permiten relacionar la persistencia delbancoserelacionanconelgrado dedisturbio y el tipo de dispersión; teniendo en cuenta lo citado por Thompson et al. (1989) y lo identificado por Montenegro (2000), se puede confirmar que áreas que han sufrido procesos de perturbación, como la reserva, revelan una tendencia a presentar alta capacidad de persistencia en las semillas y por ende en su banco. El banco de semillas persistente presentó abundancias similares para la zoocoria y la anemocoria, el banco pseudopersistente presentó un marcado predominio de la anemocoria sobre la zoocoria y, finalmente, en el banco transitorio la zoocoria se presentó como la estrategia más relevante frente a la anemocoria. La capacidad de persistencia y el tipo de dispersión pueden dar a entender porquéparael ensayolosmayoresregistros corresponden a emergentes anemócoros, si se tiene en cuenta que el máximo de las abundancias se presentó en las parcelas asociadas con el borde; estorelacionaloafirmadoporSchupp et al. (1989) y corroborado por Montenegro(2000),queindican que las semillas anemócoras se ven favorecidas en etapas tempranas de la sucesión o por medio de la llegada de éstas, producto de un flujo de aire intensificado en un claro recién creado, mientras las zoócoras arriban y dominan en etapas más evolucionadas de la sucesión (Brown 1992).
En el banco de semillas predominó levemente el número de especies de dispersión zoócora sobre la anemócora; estos resultados concuerdan con estudios realizados en los trópicos por Swaing & Hall (1983) en bosques de Ghana y por Young et al. (1987, citado en Garwood 1989), en un bosque de Costa Rica, donde para ambos casos el banco fue dominado por la zoocoria, debido posiblemente a que estos bosques no habían sufrido disturbios desde años atrás al estudio. Esta característica se presenta de igual manera para la Reserva Forestal Cárpatos, la cual viene fluctuando desde un proceso de sucesión sin intervenciones a disturbios periódicos. Este, pues, es otro indicador de que la reserva se encuentra en una etapa de desarrollo sucesional asociado a la madurez de la comunidad, ya que, de acuerdo a lo referido por Howe & Smallwood (1986), Jaimes & Rivera (1990), Brown (1992), Vargas (1998) y Montenegro (2000), en las primeras etapas de la sucesión existe un predominio por parte de las especies anemócoras que aprovechan el disturbio, mientras que las especies zoócoras empiezan a llegar posteriormente, sobre todo por medio de las aves.
Vegetación y BSG
Se registraron para el total del estudio 83 especies; 61 especies para la vegetación en pie y 55 especies registradas en el BNG.Estos valor es superan significativamente los registros de Jaimes & Rivera (1990), Posada & Cárdenas (1999), Montenegro (2000), Olaya (2000), Acosta (2004) y Cardona (2004), que sonlosestudiosmásrelevantesrealizadosdebancos para los ecosistemas altoandinos. La vegetación en pie y el banco desemillas comparten 34 especies; la vegetaciónpresentóbancoenel44%desusespecies que representa el 75% del total de la abundancia del banco, contrario a lo hallado por Montenegro (2000), que afirma que la composición de su banco revela la conformación de la vegetación en estados sucesionales anteriores al estudio. Dicho comportamiento del banco puede deberse, de acuerdo a Pickett & McDonnell (1989) y Rico-Gray & García-Franco (1992),aqueelnúmerode especies encomúnentre el banco y la vegetación es bajo en áreas muy perturbadas (Gibson & Brown 1991), mientras que en bosques perturbados con presencia de comunidades maduras esta cifra aumenta (Fenner 1995). En otras palabras, los bancos de semillas pueden presentar una imagen dual, como lo afirman Grandin y Rydin (1998) y Falinska (1999), que califican al banco de semillas como la memoria ecológica sucesional del lugar, o afirman que éste puede considerarse como un espectro de la vegetación actual en diferentes estados sucesionales. Para el presente estudio en particular aplican ambos postulados, pues el banco también presenta especies exclusivas; sin embargo, la composición del banco presenta en su mayoría un reflejo del dinamismo y evolución de la vegetación actual.
Dinámica espacio-temporal del BSG
Las parcelas que presentaron las mayores oscilaciones con respecto a la normalidad en cuanto a la abundancia, son también las que poseen mayor diversidad y riqueza; para este caso esta tendencia serelaciona hacia las parcelas de borde ydisminuye de manera gradual hacia la comunidad vegetal madura, concordando con lo concluido por Acosta (2004). La diversidad y riqueza del banco fue mayor en las parcelas donde existió más heterogeneidad de especies, de la misma manera que ocurrió con los muestreos. Esta relación de variación espacio-temporal fue también identificada en las parcelas realizadas por Montenegro(2000) y Acosta (2004); el BSG tuvo mayor dinámica en registros de abundancia a nivel espacial en aquellos puntos de las parcelas donde las coberturas vegetales son más heterogéneas, dominadas en su mayoría por vegetación correspondiente a los estados primarios del desarrollo sucesional.
FENOLOGÍA Y TIPO DE DISPERSIÓN
Fenofases de producción
Los patrones fenológicos de la floración y fructificación parecen estar influenciados directamente por los factores bióticos y abióticos (Hilty 1980), por lo que se puede afirmar que existe una correlación entre la estacionalidad de las fenofases de producción y el régimen de precipitación; así lo corroboran estudios realizados en el trópico como los efectuados por Smythe (1970) y Fleming et al. (1987) y otros realizados en ecosistemas altoandinos como los de Morales (2002) y Sánchez (2003). La floración, aunque es constante durante los doce meses del año, parece presentar un patrón respecto a su máxima producción con la época de menos precipitación (Carabias & Guevara, 1985, Morales 2002); así lo citan autores en diversos trabajos para el trópico tanto en tierras bajas(Croat 1975,Mori & Kallunki 1976 y Marquis 1988) como en altas (Bonilla & Zuloaga 1994). La fenofase de producción de flores puede estar asociada a la época seca para aumentar el porcentaje de éxito de polinización (Fagua 2002, Sánchez 2003), de esta manera,aumentan las probabilidades de generación y producción de frutos durante la temporada de lluvias (Williams et al. 2002).
Fenofases de fructificación y síndromes de dispersión
De acuerdo a Cuatrecasas (1934) y Howe & Smallwood (1982) el comportamiento esperado en los bosques húmedos es que la zoocoria predomine como estrategia sobre la anemocoria. Este dictamen concuerda con lo determinado en este estudio, dado que la zoocoria se presentó como la estrategia dominante para la vegetación en pie y el banco de semillas, presentando fructificación con registros superiores al 65% del total de los individuos durante todo el año. Este porcentaje es de esperarse para la zoocoria si se tiene en cuenta que los vertebrados y en especial las aves se reconocen como dispersores de semillas de más del 50% de las especies de árboles y arbustos tropicales(Fleminget al. 1987, Fonseca 2001).
Existió estacionalidad de la producción de frutos con respecto al régimen de precipitación, comportamiento contrario al observado por Bonilla & Zuloaga (1994) en un bosque altoandino; sin embargo, otros estudios llevados a cabo en el trópico (Howe & Smallwood 1982 y Wikander 1984) y otros realizados en bosques alto andinos (Montenegro 2000, Morales 2002 y Sánchez 2003) concuerdan con la tendencia de estacionalidad de las fenofases. La periodicidad de fenofases entre especies varía; así, especies pertenecientes a comunidades maduras presentan ciclos cortos y específicos de reproducción. Las especies que corresponden a estados sucesionales iniciales presentaron fenofases de producción más largas; esta variabilidad periódica es apoyada por Marquis (1988), quien afirma que generalmente las especies pertenecientes a estados sucesionales tempranos presentan ciclos fenofásicos más extensos que las que se encuentran en comunidades maduras.
Incidencia de la avifauna en la dinámica de dispersión
Durante los doce meses del año se contó con registros de heces y visitas en todos los ensayos (cercanía, medianía, lejanía). Este patrón permite identificar que las aves son las responsables directas del éxito del establecimiento de algunas especies de plantas emergentes en los claros, como lo describen ensus estudios Martínez & Álvarez (1986) y Murray (1987). Para determinar la efectividad de la dispersión a diferentes escalas y distancias del bosque es preciso evaluar aspectos cuantitativos relacionados con las visitas y el número de semillas encontrados (Jordano & Schupp 2000). Para este estudio se presentaron los mayores registros en todos los aspectos (especies de semillas, número de semillas, número de visitas y especies de aves) para las perchas que se encontraban en la cercanía del bosque y disminuyó gradualmente su dinámica a medida que aumentaba la distancia del borde. El comportamiento de la dinámica de las perchas frente a la distancia concuerda con lo publicado por Howe & Primack (1975), que sugiere que la distancia a la cual son trasportadas las semillas depende de los patrones de movimiento y forrajero de las aves, así como la distancia y dirección de los vuelos depende de la distribución de frutos, semillas y sus parentales (Howe&Primack1975, Herrera&Jordano1981); en otras palabras, las probabilidades de dispersión por parte de las aves aumenta proporcionalmente a la disponibilidad y proximidad del recurso que por lo general se asocia con el bosque. La totalidad de las variables presentaron una estrecha relación con el régimen de precipitación, mostrando una estacionalidad definida en la cual los patrones fenológicos de las plantas tienen lugar cuando las condiciones climáticas son favorables para la polinización de las flores y dispersión de los frutos por parte de los frugívoros (van Schaik et al. 1993), razón por la cual la temporada de mayor producción de frutos (julio), que coincide con elpico de precipitación, se convierte también en la época de mayor dispersión de semillas y variabilidad de especies dispersoras y dispersadas, reafirmándose lo sugerido por Poulin et al. (1999) cuando establece que la sincronía en la producción de frutos entre individuos y especies permite atraer en conjunto a un mayor número de dispersorespotenciales.
La mayor dispersión regional ocurrió en la época seca, contrario a lo esperado, dado que este periodo es el demenor oferta de frutos maduros;la respuesta ante esta tendencia es que durante la estación seca y ante la escasez de otros recursos alimenticios como insectos, las aves tienen que cambiar la constitución de su dieta pasando a ser frugívoros temporales (Janzen 1983, Levey 1988, Poulin et al. 1999,Sánchez2003). Asímismo, durante la escasez del recurso las aves tienen que recorrer distancias más largas para conseguir su sustento. La mayoría de las especies y familias registradas en las perchas pertenecen a estados de desarrollo inicial o intermedio, como es el caso de Arcytophyllum sp., Phytolacca bogotensis, Rubus floribundum, Miconia squamulosa y Ocotea sp., entre otras; entre las familias se registró a Melastomataceae, Rubiaceae, Lauraceae y Rosaceae. Esta tendencia concuerda con la estrategia de continuidad en la fructificación que presentan algunas especies por la competencia para atraer el mayor número de dispersores potenciales (McKey 1975), en la que existe una gran oferta de frutos pequeños con semillas pequeñas y maduraciónsincrónica (Morales2002). Este patrón atrae a diversos frugívoros generalistas que se concentran en aquellas especies de mayores cosechas (Murray1988), que resultan ser aquellas asociadas a las etapas iníciales de la sucesión (Foster & Janson 1985), como ocurre para este estudio.
CONCLUSIONES
Puede afirmarse que las parcelas relacionadas con el borde, sobre todo la que contiene un claro, presentan un comportamiento más dinámico, definido por la heterogeneidad en sus coberturas, la cual permite mayor distribución y participación de diferentes especies, hábitos y estrategias de dispersión, indicando que en lugares con vegetación sérica intermedia la probabilidad de encontrar más diversidad y número de semillases mayor.
La relación entre la zoocoria y la anemocoria es inversamente proporcional en cuanto a su participación presencial en la Reserva Forestal Cárpatos; deestamanera, en comunidades que se encuentran en etapas iníciales de desarrollo sucesional la anemocoria juega un papel importante y a medida que se avanza hacia las comunidades maduras su participación va disminuyendo gradualmente, permitiendo a la zoocoria primar como la estrategia de dispersión más importante para la reserva.
La determinación del volumen mínimo indicó que no existe un patrón específico para los componentes que integran el banco de semillas de los bosques altoandinos.
El banco de semillas de la reserva presenta una tendencia a ser constante espacio-temporalmente, al poseer una buena cantidad de semillas latentes distribuidas en todos los estados serales y en cualquier época del año, independientemente de las fenofases en que se encuentren las especies de la vegetación en pie. De igual manera, es importante resaltar la alta tasa de renovación de semillas, al presentarse el banco transitorio en el segundolugar de relevancia.
El BSG tuvo mayor dinámica en registros de abundancia a nivel espacial en los puntos donde las coberturas vegetales se presentan másheterogéneas y a nivel temporal en los periodos que coinciden con las épocas de mayor precipitación, debido a que la mayoría de semillas ya han sido dispersadas y se encuentran en el suelo dispuestas a germinar.
Los patrones de fructificación, frente al tipo de dispersión, presentaron estacionalidad respecto al régimen de precipitación. Las especies zoócoras presentaron una estrecha relación con las temporadas de mayor humedad, mientras que las especies anemócoras parecen asociarse al periodo anterior a las época de lluvias, cuando el ambiente se torna más seco y la brisa, aunque constante, es suave, caso contrario a la barocoria que parece responder mejor a la intensidad de lluvia y viento.
El ensayo de las perchas puede considerarse exitoso, dado que durante los doce meses del año se contó en mayor o menor medida con registros de heces y visitas en todos los ensayos (cercanía, medianía y lejanía del bosque).
Contrario a lo esperado, el mayor alcance en términos de dispersión (número de visitas y área de dispersión) se presentó durante la época de mayor escasez de recursos; esta tendencia se relaciona con el hecho de que algunas especies de aves presentan estrategias alimenticias estacionales y por que antela disminución de la oferta de frutos para su sustento las aves aumentan el radio de interacción, variando la dispersión de un fenómeno local a uno regional.
AGRADECIMIENTOS
Al centro de investigaciones de la Universidad Distrital Francisco José de Caldas, por su respaldo económico. A Corpoguavio por aprobar esta investigación en sus predios. A la familia Rodríguez Otálora por su incondicional apoyo durante la fase de campo. A Javier Burgos por sus valiosas apreciaciones.
REFERENCIAS BIBLIOGRÁFICAS
Acosta, M. 2004. Efecto sobre el banco de semillas germinable en un fragmento de bosque altoandino (páramo de Guerrero). Trabajo de grado. Universidad Nacional de Colombia. Pg. 89.
Bertiller, B. 1991. Seasonal Variation in the Seed Bank of a Patagonian Grassland in Relation to Grazing and Tography. Journal of Vegetation Science 2: 47-54.
Bonilla, A. & G. Zuloaga. 1994. Fenología de algunas especies de plantas del páramo "El Granizo", Monserrate (Cundinamarca, Colombia). Academia Colombiana de Ciencias Exactas, Físicas y Naturales. Bogotá. Colección Jorge Álvarez Lleras 6: 485-501.
Brown, D. 1992. Estimating the Composition of a Forest Seed Bank: a Comparison of the Seed Extraction and Seedlink Emergence Methods. Canadian Journal of Botany 70: 1603-1612.
Cantillo, E. K. Rodríguez & A. Avella. 2004. Caracterización florística, estructural, diversidad y ordenación de la vegetación en la Reserva Forestal Cárpatos, Guasca, Cundinamarca. Universidad Distrital Francisco José De Caldas. Bogotá. Pg. 150.
Carabias, J. & S. Guevara. 1985. Fenología de una selva tropical húmeda y en una comunidad derivada: Los Tuxtlas, Veracruz. En: Gómez-Pompa, A. & S. del Amo (eds.). Investigaciones sobre la regeneración de selvas altas en Veracruz, México. Volumen II. Págs. 27-78.
Cardona, A. 2004. Potencial de regeneración del banco de semillas germinable en dos tipos de bosque subandino: Implicaciones para la restauración (Reserva Biológica Cachalú, Encino, Santander). Trabajo de grado. Universidad Distrital francisco José de Caldas. Bogotá.
Conell, H. 1978. Diversity in Tropical Rain Forests and Coral Reefs. Science 1999: 1302-1310.
CORPOGUAVIO. 2000. Plan de Recuperación y Manejo Forestal, predio Cárpatos. Bogotá.
Croat, T. 1975. Phenological Behavior of Habit and Habitat Classes on Barro Colorado Island. Biotropical 7: 270-277.
Cuatrecasas, J. 1934. Observaciones geobotánicas en Colombia. Serie botánica 27. Trabajos del Museo Nacional de Ciencias Naturales. Madrid. Pg. 143.
Fagua, C. 2002. Estrategia de reproducción sexual de una población de E. grandiflora en el Parque Nacional Natural de Chingaza. Universidad Nacional de Colombia. Departamento de Biología. Bogotá.
Falinska, K. 1999. Seed Bank Dynamics in Abandoned Meadows during a Twenty Year Period in the Bialowieza National Park. Journal of Ecology 87: 461-475.
Fenner, M. 1995. Ecology of Seed Banks. En: Kigel, J. Seed Develoment and Germination. Inc. New York. Págs. 507-527.
Fleming, T. R. Bridtwisch & G. Whitesides. 1987. Patterns of Tropical Vertebrate Frugivore Diversity. Annual Review of Ecology and Systematics 18: 91-108.
Fonseca, T. 2001. Dinámica de la dispersión de semillas por aves en un pastizal con perchas artificiales en comunidades de vegetación altoandina. Tesis de pregrado. Pontificia Universidad Javeriana. Bogotá.
Foster, S. & C. Janson. 1985. The Relationships between Seed Size and Establishment Conditions in Tropical Woody Plants. Ecology 66: 773-780.
Garwood, N. 1989. Tropical Soil Seed Banks. Ecology of Soil Seed Bank. Academic Press. San Diego, California. Págs. 149-209.
Gibson, D. & K. Brown. 1991. The Effects of Grazing on Local Colonization and Extinction during Early Succession. Journal of Vegetation Science 2: 291-300.
Grandin, U. & H. Rydin. 1998. Atributes of the Seed Bank after a Century of Primary Succession on Islands in Lake Hjälmaren, Sweden. Journal of Ecology 86: 293-333.
Grime, J. 1989. Seed Bank in Ecological Perspective. En: Leck, M.A., V.T. Parkery R.L. Simpson (eds.). Ecology of Soil Seed Bank. Academic Press. San Diego, California. Págs. 15-27.
Guevara, S. & A. Gómez-Pompa. 1972. Seed from Surface Soils in a Tropical Region of Veracruz, Mexico. Journal of the Arnold Arboretum 53 (3): 312-335.
Herrera, C. & P. Jordano. 1981. Prunus mahaleb and Birds: The High Efficiency in the System of a Temperate Fruiting Tree. Ecological Monographs 51: 203-218.
Herrera, R. D. Ulloa, O. Valdés-Lafont, A.A. Priego & A. Valdés. (eds.) 1997. Ecotechnologies for the Sustainable Management of Tropical Diversity. Nature y Resources 33: 2-7.
Hilty, S. 1980. Flowering and Fruiting Periodicity in a Premontane Rainforest in Pacific Colombia. Biotropical 12: 292-306.
Howe, H. & R. Primack. 1975. Differential Seed Dispersal by Birds of the Tree Casearia nitida (Flacurtaceae). Biotropical 7 (4): 278-283.
Howe, H. & J. Smallwood. 1982. Ecology of Seed Dispersal. Annual Review of Ecology and Systematics 13: 201-208.
Jaimes, V. & D. Rivera. 1990. Banco de semillas y tendencias en la regeneración natural de un bosque altoandino en la región de Monserrate (Cundinamarca, Colombia). Trabajo de grado. Facultad de Biología. Universidad Nacional de Colombia. Bogotá. Pg. 56.
Janzen, D. 1983. Birds. En: Janzen, D. (ed). Costa Rica Natural History. University of Chicago Press. Págs. 502-549.
Jordano, P. & E. Schupp. 2000. Seed Dispersal Effectiveness: The Quantity Component and Patterns of Seed Rain for Prunus mahaleb. Ecological Monographs 70: 591-615.
Levey, D. 1988. Spatial and Temporal Variation in Costa Rican Fruit and Fruit-Eating Bird Abundance. Ecological Monographs 58: 251-269.
Magurran, A.E. 1989. Diversidad ecológica y su medición. Ediciones Vedra. Barcelona. Pg. 200.
Mahecha, G. 2004. La vegetación del territorio CAR. CAR (Cundinamarca). Bogotá. Pg. 620.
Marquis, R. 1988. Phenological Variation in the Neotropical Understory Shrub Piper arienum: Causes and Consequences. Ecology 69: 1552-1565.
Martínez-Ramos, N. & E. Álvarez-Buylla. 1986. Seed Dispersal, Gap Dynamics and Tree Recruitment: the Case of Cecropia obtusifolia at Los Tuxtlas, Mexico. En: Estrada, A. & H. Fleming. Frugivory and Seed Dispersal. Junk Publishers. La Haya. Págs. 333-346.
Mc Graw, J. &M. Varek. 1989. The Role of Buried Viable Seeds in Artic and Alpin Communities. En: Leck, M., V. Parker y L. Simpson (eds.). Ecology of Soil Seed Bank. Academic Press. San Diego, California. Págs. 91-105.
Mckey, D. 1975. The Ecology of Coevolved Seed Dispersal Systems. En: Gilbert, L. & P. Raven (eds.). Coevolution of Animals and Plants. University of Texas Press. Págs. 159-191.
Montenegro, A. 2000. Estrategias de dispersión y regeneración por banco de semillas en dos comunidades de bosque altoandino en Colombia. Trabajo de grado. Facultad de Biología. Universidad Nacional. Bogotá. Pg. 132.
Morales, N. 2002. Ecología de la dispersión de semillas por aves de Brunellia colombiana en un bosque altoandino (Parque Nacional Natural Chingaza). Tesis. Universidad Nacional de Colombia. Pg. 85.
Mori, S. & J. Kallunki. 1976. Phenology and Floral Biology of Gustavia superba (Lecythidaceae) in Central Panama. Biotropical 8: 184-192.
Murray, G. 1987. Selection for Optimal Fruit Crop Size in Bird-Dispersed Plants. American Naturalist 129: 18-31.
Newstrom, L. G. Frankie, H. Baker y R. Collwell. 1994. Diversity of Long-term Flowering Patterns. En: Mc Dade, K., H. Bawa, H. Hespenheide & G. Hartshorn (eds.). Ecology and Natural History of a Neotropical Rainforest. Págs. 142-160.
Olaya, A. 2000. Efecto de borde sobre el banco de semillas germinable del bosque altoandino (Embalse de San Rafael, La Calera, Cundinamarca). Tesis de grado. Pontificia Universidad Javeriana. Bogotá.
Pedraza, L. 2000. Las plantas con flores de los alrededores de la laguna de Chisacá (Sumapaz, Colombia). Trabajo de grado en Biología. Universidad Nacional de Colombia. Facultad de Ciencias. Bogotá. Pg. 237.
Piquett, S. & M. McDonnell. 1989. Seed Bank Dynamics in Temperate Deciduous Forests. En: Allessio, M., V. Parker & R. Simpson (eds). Ecology of Soil Seed Banks. Academic Press. San Diego, California. Págs. 123-146.
Posada, C. & C. Cárdenas. 1999. Ecología de los bancos de semillas en una comunidad vegetal de páramo sometida a disturbios por quema y pastoreo (Parque Nacional Natural Chingaza). Trabajo de grado. Universidad Nacional de Colombia. Facultad de Biología. Bogotá. Pg. 190.
Poullin, B. J. Wright, G. Lefebvre &O. Calderón. 1999. Interspecific Synchrony and Asynchrony in the Fruiting Phenologies of Congereric Bird-Dispersed Plants in Panama. Journal of Tropical Ecology 15: 213-227.
Rice, J. 1989. Impacts of Seed Banks on Grass land Community Structure and Population Dynamics. En: Leck, M., V. Parker y R. Simpson (eds.).. Ecology of Soil Seed Bank. Academic Press. San Diego, California. Pg. 211-230.
Rico-Gray, V. & J. García-Franco. 1992. Vegetation and Soil Seed Bank of Successional Stages in Tropical Lowland Deciduous Forests. Journal of Vegetation Science 3: 617-624.
Rossi, R. P. Lavelle & E. Tondof. 1995. Statistic Soil Biology. Geostatical Analysis. European Journal of Soil Biology 31 (4): 173-181.
Saldarriaga, J.G. 1994. Recuperación de la selva de tierra firme en el alto río Negro, Amazonia Colombiana-Venezolana. Tropenbos. Pg. 201.
Sánchez, M. 2003. Fenología reproductiva y dispersión de semillas del arbusto altoandino Monina salicifolia. Tesis de grado. Universidad Nacional de Colombia. Bogotá. Pg. 65.
Schupp, E. H. Howe, C. Augspurger & D. Levey. 1989. Arrival and Survival in Tropical TreefallGaps. Ecology 70(3): 562-564.
Society for Ecological Restoration (SER). 2004. SER International Primer on Ecological Restoration. Society for Ecological Restoration. [disponible en: http://www.ser.org/content/ecological_restoration_primer.asp].
Simpson, R. M. Allessio & V. Parker. 1989. Seed Bank: General Concepts and Methodological Issues. En: Leck, M. , V. Parker y R. Simpson (eds. ). Ecology of Soil Seed Bank . Academic Press. San Diego, California. Págs. 3-8.
Smythe, N. 1970. Relationships between Fruiting Seasons and Seed Dispersal Methods in a Neotropical Forest. American Naturalist 104: 25-35.
Sorzano, C. 2001. Regeneración reproductiva en un sitio abandonado e invadido por Pteridium aquilinum Khun. Implicaciones para la restauración. Reserva Biológica Cachalú. Encino, Santander, Colombia. Trabajo de grado. Pontificia Universidad Javeriana. Facultad de Ciencias. Departamento de Biología. Bogotá.
Swaing, D. & J. Hall. 1983. Early Succession on Cleared ForestLandin Ghana. Journal of Ecology 71: 601-627.
Thompson, K. J. Bakker & R. Bekker. 1998. Ecological Correlates of Seed Persistence in Soil in the North-West European Flora. Journal of Ecology 86: 163-169.
van Schaik, C. 1993. The Phenology of Tropical Forest: Adaptative, Significance and Consequences for Primary Consumers. Annual Review of Ecology and Systematics 24: 353-377.
Vargas, O. 1998. Un modelo de sucesión/regeneración de los páramos después de las quemas. Caldasia 19 (1-2): 331-345.
Wikander, T. 1984. Mecanismos de dispersión de diásporas de una selva decidua en Venezuela. Biotropical 16 (4): 276-283.
Williams-Linera, G. & J. Meave. 2002. Patrones fenológicos. En: Guariguata, M. y G. Kattan (eds.). Ecología y conservación de bosques neotropicales. Libro universitario regional de Costa Rica. Págs. 407-431.





Licencia
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






