DOI:
https://doi.org/10.14483/2256201X.3265Published:
2000-01-01Issue:
Vol. 6 No. 13 (2000)Section:
Scientific articleBases Ecológicas Para La Silvicultura Del Bosque Natural Estudio De Caso Del Catival (Prioretum Copaiferae)
Downloads
References
ARROYO, M.T.K. y SQUEO, F. 1990. Relationship between plant breeding systems and pollination. En: Breeding Systems and Pollination. New York, Academic Press. pp: 205-227. Tabls., gr fs.,maps.
CORPORACION NACIONAL DE INVESTIGACION Y FOMENTO FORESTAL. Archivo Técnico T-5. Project CONIFPIZANO S.A.; Investigación para el Manejo del Catival.
CORPORACION NACIONAL DE INVESTIGACION Y FOMENTO FORESTAL. 1993. Estado del conocimiento sobre la Asociación Catival en Colombia. Informe de Avance Contrato CONIF-COLCIENCIAS. Santafé de Bogotá, D.C. Documento en borrador.
CORPORACION NACIONAL DE INVESTIGACION Y FOMENTO FORESTAL: PIZANO. 1995a. Estudio comparativo: caracterización de la asociación catival en estado climácico y 21 arios después de la intervención con énfasis en su biodiversidad. La Gira y Clavellino, Bajo Atrato, Chocó, Colombia. Santafé de Bogotá, 180 p. Ilus., tabls., gráfs.
CORPORACION NACIONAL DE INVESTIGACION Y FOMENTO FORESTAL. 1995b. Análisis de los proyectos BAL-RN-0689, BAL-RN-07-9 y BAL-RN-08-91 que enmarcan la investigación de la dinámica y manejo de la regeneración natural en la asociación catival. Informe del contrato CONIFPIZANÓ S.A. Tabs., gráfs.
ECHAVARRIA, J. y VARON, T. 1988. Estudio dendrológico de la asociación catival en La Balsa, Chocó. Tesis Ing. For., Medellín, Colombia. Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia. 548 p.
ESCOBAR, J. y VASQUEZ F. 1987. Caracterización de tipos de cativales. Tesis Ing. For., Medellín, Colombia. Facultad de Agronomía, Universidad Nacional de Colombia. 201 p.-
FINEGAN, B. 1993. Curso Intensivo Internacional de Silvicultura y Manejo de Bosques Naturales Tropicales. 1993. I. Tema: bases ecológicas para la silvicultura. Turrialba, Costa Rica, Catie. 225 p. Ilus., tabls., gr fs.
INSTITUTO COLOMBIANO DE HIPROLOGIA, METEOROLOGIA Y ADECUACION DE TIERRAS (HIMAT). 1986. Hojas de Registros diarios . de la Estación climatológica Grado II "La Balsa", Riosucio -Chocó. Archivo Técnico de la Estación Silvicultura' "La Balsa".
LINARES, RICARDO. 1988. Estudio preliminar de la asociación catival en Colombia. Bogotá, Conif y Pizano S.A. 68 p. (Conif, Serie de Documentación No.17).
LINARES, RICARDO. 1995. Bases ecológicas para la silvicultura y el aprovechamiento de los bosques naturales en Colombia. En: Talleres regionales sobre aprovechamiento de los bosques naturales; Florencia (Caquetá), Leticia (Amazonas), Bucaramanga (Santander) y Cali (Valle), 1995. Proyecto Concesiones y Permisos Forestales PPD 11/93 Organizacion internacional de Maderas Tropicales - Ministerio del Medio Ambiente República de Colombia.
LINARES, RICARDO. 1998. Conservación, utilización y manejo sostenible del Catival (Prioretum copatferae). Resultados de 16 arios de investigaciones. Convenio Conif - Pizano S.A. Santafé de Bogotá, Conif. Documento en edición para publicación. 118 p.
LINARES, RICARDO Y MARTINEZ HIGUERA, HUGO. 1991. La regeneración natural temprana del bosque de cativo en el Chocó, Colombia. Bogotá, Conif. Serie Técnica No. 30.
LINARES, RICARDO et al: 1997. Bases ecológicas para la silvicultura del catival (Prioretum copaiferae). Informe Preliminar del Proyecto sobre Publicación del Catival. Documento de avance para discusión presentado por la Corporación Nacional de Investigación y Fomento Forestal -- Conif. Santafé de Bogotá, Conif, Contrato Pizano S.A. -- Conif 48 p.
MONTERO, M. y CORDOBA G. 1996. Estudio de la dinámica poblacional de Prioria copaifera (Grisebach) en un bosque de la llanura aluvial del río Atrato. Santafé de Bogotá , Universidad Nacional de Colombia. 150 p. Ilus., tabls., gráfs., maps. Tesis (Biól.)
OROZCO, R.H. 1995a. Contribución al estudio de los efectos del aprovechamiento forestal sobre la comunidad de aves de la consociación Priorietum copaiferae (Catival), en el Chocó colombiano. Santafé de Bogotá, D.C., Pontificia Universidad Javeriana. Facultad de Ciencias. Departamento de Biología. Ilus., tabls., gráfs., maps. Tesis (Biól.).
OROZCO, R. H. 1995b. Visión general de la Avifauna asociada a los Cativales del área de aprovechamiento forestal "La Balsa" (Riosucio-Chocó). Pizano S.A. Dirección de Recursos Naturales. 38p.
RIOS, SANDRA. 1996. Estudio de la arquitectura de la comunidad Prioria copaifera Grisebach, (Caesalpinaceae), en un bosque inundable de la región del bajo Atrato, Chocó, Colombia.
RODRIGUEZ, J. VICENTE., HERNANDEZ, J. Y DÁVILA, A.. 1995. Manejo de Vida Silvestre. Pizano S.A. Región de Domingodó, Chocó. 100p.
ROLLET, B. 1974. L'architecture des forest denses humides sempervirentes de plaine. Nogent sur Mame, Francia. Centre Technique Forestier Tropical. 298 p.
URDANETA, M. F. 1995. Diferencias en la composición y estructura de la comunidad de aves en época seca de tres agroecosistemas y un bosque primario asociados a un fragmento de catival maduro en el Chocó Colombiano. Pontificia Universidad Javeriana. Facultad de Ciencias, Departamento de Biología. Santafé de Bogotá. 173p.
WHITMORE, T. 1984. Tropical rain forest of de Far East. Oxford, Reino Unido: Clarendon Press. 314 p.
SCHULZ, J.P. 1960. Ecological studies of rain forest in Northern Suriname. Amsterdam, - NoordHollandsche Vitgevers Maatschaooyj. 267 p.
How to Cite
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Download Citation
Artículos Cientificos
Colombia Forestal, 2000-09-01 vol:6 nro:13 pág:5-37
Bases ecológicas para la silvicultura del bosque natural estudio de caso del catival (Prioretum copaiferae)
Ricardo Linares Prieto
Introducción
Para la producción forestal se requiere tener la capacidad de identificar y resolver los problemas de la conservación de los ecosistemas, evento en el cual es fundamental conocer y aplicar las bases ecológicas y así seleccionar métodos sostenibles de aprovechamiehto y manejo del bosque. La identificación y la caracterización de los aspectos ecológicos que rigen a una comunidad forestal permiten resolver varios de los próblemas existentes para alcanzar su utilización sostenible. Así como los ecosistemas, comunidades, poblaciones y especies vegetales son diversas, y poseen sus propias características, su eventual aprovechamiento requiere la solución de sendos problemas derivados de su intervención. Cada sistema de producción forestal funcionará dentro de un ambiente físico, químico y biológico heterogéneo, con variaciones sustanciales dentro de cada ecosistema, lo que exige estudios específicos, no recetas o fórmulas genéricas para el aprovechamiento y la silvicultura del bosque natural, como comúnmente está sucediendo.
Las Bases Ecológicas para la Silvicultura son los elementos mínimos para proceder y acceder al aprovechamiento y la producción forestal en términos de sostenibilidad o, al menos, de racionalidad. Su tratamiento se hace no tanto en el ámbito de la biología por sí sola, sino como una herramienta para hacer un puente o enlace entre ella y la silvicultura, con enfoque eminentemente práctico. Constituyen un tema amplio y aún relativamente nuevo, pero que día tras día adquiere mayor vigencia en el marco del Desarrollo Forestal Sostenible.
En Latinoamérica uno de los más connotados investigadores de este tema ha sido el Dr. Bryan Finegan quien ha realizado importantes aportes teóricos y prácticos. En Colombia algunos resultados se han generado mediante la sistematización y el análisis de resultados de investigaciones de campo efectuados por el autor durante el desarrollo de experiencias de silvicultura y manejo forestal en ecosistemas como el Catival, el Guandal, el Naidizal, el Manglar en el Pacífico, y algunos bosques del Caquetá y el Putumayo.
Aquí se presenta un primer desarrollo para Colombia de las Bases Ecológicas para la Silvicultura mediante el estudio de caso de la Consociación Catival (Prioretum copaiferae); se han retomado los resultados de las investigaciones ecológicas y silviculturales realizadas durante más de 16 arios por la Corporación Nacional de Investigación y Fomento Forestal —Conif- y la empresa Pizano S.A., a las que se han adicionado los logros de otras investigaciones bioiógicas auspiciadas por estas mismas dos entidades en asocio con Colciencias y algunas Universidades, especialmente mediante la financiación de tesis de grado. Se anota que este artículo constituye un resumen de uno de los capítulos de la publicación que se halla en proceso de edición y preparación para impresión, realizada por el autor con Conif, en el marco del Convenio con Pizano S.A. (Linares, 1998).
Se espera que este documento sea un aporte, de los muchos requeridos, para que los Ingenieros Forestales, los Biólogos, los Tecnólogos Fores: tales, los Técnicos en Recursos Naturales y demás profesionales relacionados, así como las instituciones, las comunidades y en general los diferentes actores del aprovechamiento y manejo de los bosques tropicales inicien o continuen el camino requerido para alcanzar la meta de conjugar la producción con la conservación forestal.
Síntéticamente las bases ecológicas para la silvicultura son las siguientes:
- Los factores ambientales del ecosistema forestal, especificados según condiciones y recursos, de acuerdo con los efectos que ejercen sobre las especies vegetales y la forma en que estas los utilizan.
- Los gradientes ambientales y las estrategias biológicas de los organismos.
- La ecología de las poblaciones del bosque.
- Los gremios de las especies forestales y su biología.
- Estructura de las poblaciones, prioritariamente de las especies objeto de la producción.
- Competencia interespecífica e intraespecífica, densidades de las poblaciones, crecimientos de las especies.
- Estrategias y hábitos de reproducción.. Mutualismos, agentes y sistemas de polinización floral y diseminación de semillas.
- Organización del bosque, estructura total (horizontal y vertical), composición florística, asociaciones y consociaciones vegetales involucradas.
- Procesos dinámicos y sucesionales de las especies (leñosas y herbáceas), el potencial cualitativo y cuantitativo de su regeneración natural y su tabla de vida.
1. Las condiciones y los recursos del ecosistema catival
Las condiciones son factores abióticos del ambiente que no son empleados directamente para realizar procesos fisiológicos ni para formar tejidos de los organismos y que varían en el tiempo y el espacio (como temperatura, pH, humedad relativa y humedad del suelo); según sus gradientes, las plantas y los animales adoptan. estrategias. Los recursos son factores que sí emplean los organismos, bien sea para formar biomasa o para realizar procesos vitales (agua, suelo, aire, nutrientes, materia orgánica y radiación solar, entre otros).
1.1 La temperatura
Las áreas del catival corresponden a un régimen isomegatérmico: temperaturas altas (media mensual > 24 °C) que se mantiene relativamente constante a lo largo del ario, con fluctuaciones máximas de 5 °C. Según registrós de 15 arios de las estaciones climatológicas de Riosucio, La Balsa, Unguía, Dorningodó y Murindó (Chocó), la temperatura media anual de las zonas con bosques de cativo es de 26,5 °C. En estos se presentan dos temperaturas: la primera, en el dosel superior; allí las hojas, como órganos de asimilación, mantienen casi siempre una temperatura mayor a la ambiental; otra, en el nivel del suelo, donde la temperatura es más baja que la ambiental pues los renuevos de las -plantas y el suelo requieren mantener un ambiente fresco que garantice la producción fotosintética y un nivel de humedad favorable para su funcionamiento. En los grandes claros del catival, bien sean originados por procesos naturales (por ejemplo por un ciclón) o antrópicos (causados por el aprovechamiento forestal o la agricultura migratoria), hay una abrupta elevación de la temperatura, que causa efectos negativos sobre la regeneración natural, como la disminución de la tasa fotosintética y en casos extremos, la muerte.
Según los estudios de fenología realizados por Conif (1982 a 1994) en la zona de "La Balsa", (Riosucio, Chocó) se deduce que hay una aparente coincidencia entre la iniciación de la formación de botones florales (octubre-noviembrediciembre) y la disminución de la temperatura durante los mismos meses; como se observa en los registros de las estaciones climatológicas "La Balsa", "La Honda" y "Riosucio".
1.2. La Humedad
Las zonas de catival se caracterizan por presentar un régimen monomodal de precipitaciones. El balance hídrico indica que en los cativales del río León y del norte del Atrato hay entre 1 y 2 meses ecosecos (que de un sector a otro se ubican entre enero y marzo); es decir, que durante la mayor parte del ario se presenta suficiente disponibilidad de agua para las plantas y que hay un período corto de déficit, indispensable para los procesos fenológicos de reproducción. Entre marzo y diciembre las precipitaciones superan a la evapotranspiración, en tanto que entre enero y febrero sucede lo contrario (déficit en la saturación del suelo); hacia la cuenca media del río Atrato no se presentan meses ecosecos. Debido a estas razones en el catival predominan los árboles perennifolios, con mezcla de caducifolios, y la vegetación pertenece a la Hydrophytia, selva húmeda tropical semidecidua.
Este bosque depende de los regímenes hídricos y tolera condiciones extremas de humedad con prolongadas y fuertes inundaciones aluviales; el régimen de lluvias es determinante para que su vegetación presente variaciones en la fisionomía y la estructura. Por la mayor precipitación, en las zonas de vida muy húmedo y pluvial tropical, el catival debería ser más exhuberante y desarrollado (Tossi, 1976); pero el estudio de Caracterización de Cativales (Escobar y Vásquez, 1987) no demostró diferencias significativas en tal sentido; y si las hay, son sutiles.
1.3. El pH y la Humedad del suelo
Los severos regímenes hídricos y la marcada presencia de texturas arcillosas en los horizontes minerales del catival originan un drenaje pobre o lento; lo que implica que haya bajas tasas de difusión del oxígeno y el CO2 alrededor de los sistemas radiculares y de los organismos de la microflora y la microfauna del suelo. La abundancia de materia orgánica en proceso de descomposición, principalmente vegetal, y la abundancia de lluvias que lavan permanente los suelos, han dado lugar a una acidez que origina la alta solubilidad de cationes potencialmente tóxicos de Aluminio, Hierro y Manganeso, inclusive. Estas circunstancias son más patentes en los cativales Ao y Al (ver tipos de catival en Linares, 1988). La dominancia de Priora copaifera se debe precisamente a que es una de las especies mejor adaptadas a tales condiciones anaeróbicas y a la limitada disponibilidad de nutrientes; en los cativales A2 y A3 (ibíd.) dichas condiciones son menos severas, de modo que el cativo pierde su dominancia, aunque la mejor calidad de sitio le permite alcanzar mejor desarrollo individual, al aumentar la tasa de metabolismo y de producción de biomasa por disponer de más nutrientes y mejorar las condiciones físicas del suelo.
1.4. La Radiación Solar
Considerada como recurso, la radiación es vital para el funcionamiento del Prioretum y tiene mucho que ver para la toma de decisiones sobre su silvicultura y manejo. Las áreas de catival están sometidas a menos de 1.500 horas anuales de brillo solar, con tendencia a disminuir hacia el centro y el sur del Chocó (Himat, 1986). La densa y casi permanente nubosidad presente en, la zona Atrato-León es una barrera para la utilización de la luz solar.
El bosque de cativo recibe la luz solar en dos, formas: directa en una menor proporción y difusa en la mayor parte. Su funcionamiento se basa más que todo en la luz de difusión que se origina por la interceptación que de la luz solar hacen hojas, ramas y tallos sobre todo de árboles de los estratos superior e intermedio. Aquí Prioría copaifera juega un papel determinante habida cuenta que es la especie de mayor abundancia y expansión horizontal en el dosel del catival (Linares, 1988).
Según Baumgartner y Brunig (1980) citados por Finegan, (1993), una fracción de la radiación solar difusa es absorbida por las hojas vegetales y utilizada como recurso, y se le llama radiación solar fotosintéticamente activa (RAFA); el resto de la luz es transmitida dentro del bosque. La intensidad de RAFA disminuye paulatinamente desde el 100%, a 58 m de altura del dosel superior, hasta el 25%, a 47 m. Este 25% se mantiene hasta los 30 m en el dosel intermedio denso y después hay una disminución logarítmica hasta llegar al 5%, a los 20 m y al 2% a los 10 m; por último, es menos del 1% para la altura entre el piso y 5 m. Para el catival el modelo puede ser similar, con algunas variaciones; este modelo de distribución y uso de la radiación solar origina que todas las especies, adopten estrategias según sus requerimientos en tal sentido.
Beadle et al. (1985) citados por Finegan (ibíd.), establecen que las variaciones de radiación pueden ser sistemáticas o no sistemáticas; las no sistemáticas son impredecibles, como aquellas originadas aleatoriamente por el crecimiento de otros árboles y plantas competidores, así como las causadas por los gradientes introducidos por la apertura del dosel a través del aprovechamiento maderero, el manejo silvicultural o la caída natural de árboles. De esto se derivan las estrategias biológicas adoptadas por las especies del catival, según los gradientes de iluminación dentro del bosque; como el caso de las especies que para su óptimo crecimiento y desarrollo requieren un hábitat abierto y un alto grado de iluminación solar (heliofitismo) o las que requieren o soportan sombra (esciofitismo), con una gama de rangos intermedios. Conocer y aplicar las estrategias de las especies es clave para el manejo del ecosistema y que se analizan en el ítem sobre los Gremios de Especies. Por estas circunstancias, las técnicas y prácticas silviculturales son flexibles y varían según el gremio de cada especie involucrada.
En las operaciones de cosecha se abre el dosel abruptamente y en consecuencia, las especies herbáceas y sufrútices del sotobosque, las plántulas de la regeneración natural temprana y los fustales remanentes con hojas de sombra se ven sometidos a un intempestivo y elevado incremento en la exposición solar, ocasionando graves pérdidas de población vegetal por mortalidad, así como retraso en su crecimiento y desarrollo, por el estrés fisiológico inducido. El mayor efecto de la apertura de claros en el bosque radica en que la intensidad lumínica, y con ello la radiación, se incrementa abrupta y excesivamente. Además, esto redunda en cambios en el microclima ya que se elevan la temperatura del suelo y la evapotranspiración; por esto, el tamaño de los claros ocasionados en el aprovechamiento y manejo forestal debe ser regulado.
1.5. Las Moléculas Inorgánicas
1.5.1. El Carbono
En cuanto al CO2, la disponibilidad ambiental de este recurso supera a la demanda por las plantas y hasta donde se conoce, no presenta limitaciones para el manejo del recurso.
1.5.2. El Agua
Las especies arbóreas de mayor vitalidad en el catival están genéticamente adaptadas a condiciones extremas de disponibilidad de agua y utilizan sólo una parte de ella para la fotosíntesis; el excedente es transpirado a través de su abundante estructura de estomas foliares, lo que le sirve de termoregulador de los tejidos. Si la tasa de transpiración disminuyera drásticamente, por restricciones en la disponibilidad natural de agua en el suelo del catival, habría un colapso y la planta moriría por estrés o retardaría su crecimiento y desarrollo. Este aspecto es importante para el manejo, debido a los eventuales efectos negativos por la apertura de vías de transporte fluvial y por posibles daños causados en las ramas y la copa de árboles circunvecinos por los árboles talados que producen la drástica pérdida de área foliar.
1.5.3. Los nutrientes
Las mayores variaciones al ciclaje de nutrientes se realizan por la quema del suelo y la extracción y remoción de grandes cantidades de biomasa en el aprovechamiento forestal. Con esto se desequilibra el funcionamiento del suelo y la disponibilidad de nutrientes para la regeneración natural, los árboles remanentes y las plantas no maderables. El aprovechamiento y el manejo silvicultural causan, en diversas ocasiones, la abundante disposición de residuos por encima de los niveles naturales en los primeros arios postintervención; este fenómeno acelera la tasa de crecimiento de los árboles, tal vez a causa del incremento de nutrientes en el suelo. En el caso que se elimina el bosque natural a tala rasa para proceder a realizar la agricultura migratoria, la situación también puede ser dramática ya que el desbalance negativo de nutrientes en el suelo es irreversible.
Parece que los cambios en el contenido de nutrientes del suelo no son tan determinantes en las variaciones fitosociológicas del catival y los cambios florísticos y estructurales pueden deberse más a las variaciones en las propiedades físicas de suelo, según los levantamientos de tipos de catival reportados por Arteaga y Gómez (1997).
En cuanto al Nitrógeno, su mineralización en el catival sin disturbación es gradual y está equilibrada con las necesidades vegetales. Pero el incremento de la radiación por la áobreexposición del suelo y la consecuente pérdida de humedad aumentan la tasa de mineralización del nitrógeno orgánico acumulado en el mulch, aumentando su disponibilidad por un corto plazo aunque ésta puede disminuir en el mediano y largo plazo.
Cuando se hacen drenajes en el bosque, la rápida disminución del contenido de agua se traduce en pérdida de la hidromorfia, que incrementa la tasa de oxidación de la hojarasca y demás tipos de materia orgánica del horizonte superior. Los cambios en el régimen de drenaje pueden introducir modificaciones a la composición florística y la estructura. También se origina una disturbación en el funcionamiento y la nutrición de la población forestal, poniendo en riesgo la productividad del ecosistema de catival y el potencial cualitativo y cuantitativo de la regeneración natural necesaria para el manejo sostenible.
2. Ecología de las poblaciones
La dinámica de las poblaciones de las especies vegetales implica fenómenos complejos, producto de un largo proceso de evolución natural que se manifiesta en la biostasis a través de una cierta forma de organización, funcionamiento e interrelación de sus componentes; los fáctores exógenos, como el aprovechamiento, pueden alterar radicalmente ese modelo natural. Conocer estos aspectos y entender en qué grado y de qué manera la cosecha y el manejo del bosque les afectan, son primordiales para conciliar la conservación con el aprovechamiento forestal sostenible.
2.1. El Balance de la Población
Estudiar el ciclo de vida de las especies, al menos de las vegetales aprovechables resulta clave para facilitar el manejo de sus poblaciones. Esto es difícil en las especies arbóreas cuyo ciclo de vida es largo. Para el catival este nivel de información aún es deficiente; el ciclo de vida de sus especies importantes corresponde a una de estas dos categorías: 1°) De Reproducción Reiterada Episódica: en especies cuyos episodios de reproducción (floración, fructificación y germinación) muestran una alta correlación con la estación más seca; ej. Tabebuía rosea y Ochroma lagopus y Prioria copaifera; parece ser la categoría más común; 2°) De Reproducción Reiterada Continua, ej. en algunos árboles como los Ficus y algunas plantas parásitas.
Sobre el balance de población para el catival, se registran dos investigaciones: la primera, de Linares y Martínez (1991), halló que la regeneración natural de algunas especies arbóreas presenta una alta tasa anual de reclutamiento, con valores significativamente mayores en el catival A2, considerándola satisfactoria para asegurar la reposición "in situ" de la población bajo aprovechamiento. La época de mayor tasa de natalidad de Prioria copatfera coincide con la finalización de las fenofases de floración y fructificación de las especies arbóreas y con el advenimiento de la etapa lluviosa; la fase de mayor mortalidad de brinzales R (renuevos), y de mayor crecimiento longitudinal de los sobrevivientes, coincide con la época de mayor inundación én el bosque, particularmente en el catival Al. Se demostró que la selección natural es intensa y que de la abundante natalidad (hasta 12.700 renuevos por ha/ario) sólo un 2% llega a la fase de fustales jóvenes (alrededor de 10 cm. de diámetro), equivalente a entre 30 y 50 árboles por hectárea. El segundo trabajo, de Montero y Córdoba (1996), indica que para una población de Prioría copatfera en Catival A2 la cosecha/ario es de 30.490 semillas/ha con "natalidad neta" del 1.5% (aproximadamente 450 plántulas sobrevivientes y establecidas); de las pérdidas, el 18% (casi 5.500) fueron abortos; aunque hubo una tendencia de mayor abortaje en los árboles de mayor número de semillas producidas, fenómeno que aún no se sabe si está controlado por factores endógenos de la especie o por condiciones ambientales; esto hace pensar que la tasa de natalidad no depende tanto de la cantidad de semillas de cativo producidas en el bosque (léase cantidad de árboles semilleros), sino de la calidad del sitio para su germinación.
Estos aspectos confirmarían la necesidad de causar la menor disturbación a las condiciones del catival en el aprovechamiento, con precauciones como conservar en la medida de lo posible la humedad del suelo y su contenido de mulch y gradualizar la exposición solar con el fin de que se aumenten las posibilidades de germinación y el establecimiento de regeneración natural temprana, aunque haya una disminución de la cantidad de árboles portagranos a raíz de la cosecha de individuos maduros. La mayor tasa de mortalidad de la especie se presenta en la etapa de brinzales y las mayores causas son la competencia por condiciones y factores del sitio de agrupamiento de la semilla; la especie sufre un autoraleo fuerte, propio de la competencia intraespecífica, cuando los arbolitos se hallan en el cambio de categoría de tamaño de brinzales R (menor de 0.30 cm. de altura a la de brinzal U1 (entre 0.30 y 1.50 m de altura).
Según Tamayo(1991) la mayor depredación de semillas se debe a los insectos: Eubulus sp. (Curculionidae), Ectomielois ceratonie (Pyralidae) y un micro-himenóptero no determinado cuyo mayor ataque ocurre en la predispersión (antes de caer al suelo) lo que parece es la causa principal de la pérdida de viabilidad de las semillas. El consumo de semillas por roedores es poco significativa. Se producen cerca de 30.000 semillas/ha entre diciembre (final de la época lluviosa) y mayo (inicio de la época lluviosa). Estudios fenológicos de Conif durante 12 arios consecutivos indican que la cuantía Brinzales de cativo (Prioria copaifera) de una misma cohorte. Ríosucio, Chocó, Colombia. de producción de semillas varía de ario en año. Montero y Córdoba (1995) indican que el volumen de la cosecha de semilla del cativo es independiente del tamaño del árbol (fuste y copa) y alcanza su tamaño reproductivo a partir de un D.A.P. de 40 cm y altura de 30 m.
La germinación y el desarrollo de los brinzales son más propicios al inicio del "verano" mientras que en la época más seca (febrero a marzo) las condiciones del suelo no son propicias para tales fines. La rápida tasa de crecimiento de los renuevos de cativo y su sobrevivencia a la época de máximas lluvias e inundaciones se debe al alto contenido de reservas nutritivas en su semilla, alcanzando tasas máximas de crecimiento de hasta 1/2 m/mes en alfura (Linares y Martínez, 1991). Es decir,- que si bien la profundidad del agua es una condición que opera como gradiente ambiental para el establecimiento de la esikcle, las pérdidas deben asociarse también a otras condiciones y factores del ecosistema, no sólo al nivel de la inundación. El grado dé inundación es una limitante importante para la regeneración del cativo cuando hay disturbio antrópico del suelo, como en el caso de las huellas de tractores o del arrastre de trozas, que aumentan el grado, velocidad y temporalidad del encharcamiento afectando el patrón de natalidad de la especie. Una alta mortalidad natural de'cada cohorte de cativo se debe a la competencia intraespecífica. propia de las especies gregarias; competencia que, a la vez, está regulada con la producción gradual y no sincrónica de las semillas y súele ser incrementada por los daños mencionados.
 "Brinzales de cativo (Prioria copaifera) de una misma cohorte. Ríosucio, Choco, Colombia"
"Brinzales de cativo (Prioria copaifera) de una misma cohorte. Ríosucio, Choco, Colombia"
 "Distribución de la iluminación solar en el perfil del bosque de cativo (Priora copaifera)."
"Distribución de la iluminación solar en el perfil del bosque de cativo (Priora copaifera)."
En la práctica, ésto significa que en épocas de fructificación y germinación tal vez no se debería aprovechar cativo; o que, al menos, se podría hacer en meses posteriores a la consolidación de la población de brinzales de cada nueva cosecha. Entonces, resulta recomendable minimizar el impacto de las maquinas sobre el suelo y conservar a toda costa los árboles portagranos seleccionados.
2.2. Los Gremios de las Especies Forestales
2.2.1. Heliófitas Efímeras
Son especies de crecimiento rápido y vida corta (10 a 15 arios), claves en la colonización de sitios abiertos; tienen producción precoz y de gran cantidad de semillas, muchas de las cuales se mantienen vivas en el "banco de semillas" del suelo. Su alta capacidad fotosintética les permite un crecimiento rápido pues asignan alta cantidad de recursos a la producción de follaje, frutos y semillas. Como producto de esta estrategia biológica tiene madera liviana, suave y poco resistente. Su ciclo de aprovechamiento y manejo es corto y más eficiente en sitios abiertos más o menos extensos. Silviculturalmente, la mayoría son especies con limitaciones industriales o comerciales; excepto algunas como Ochroma lagopus (Balso).
2.2.2 Heliófitas Durables
Son especies intolerantes a la sombra, de crecimiento rápido y ciclo de vida más bien largo. De dimensiones mayores y vida más larga que las HE; facultadas para poblar más sitios que éstas, tanto en sitios abiertos claros extensos como pequeños, pero no tan rápido como las mismas; se apropian de dichos sitios después que las efímeras desaparecen, estableciéndose por un período de tiempo mucho más largo. Su capacidad fotosintética es intermedia, aunque en condiciones de sombra se torna baja. Su patrón de asignación de recursos le otorga una capacidad de crecimiento diamétrico de hasta 2 y 3 cm/ario. El patrón de reproducción puede ser precoz en condiciones favorables (sucesión secundaria), empezando a veces a los 5 años de edad, en tanto que en condiciones desfavorables (sucesión primaria) la reproducción queda suprimida hasta que el árbol establezca su copa en el dosel superior.
En este gremio es donde más comúnmente ocurre dispersión anemófila de semillas (sin ser la más eficaz). Con semillas de tamaño variable (pequeñas a grandes), generalmente con pocas reservas y con cotiledones fotosintéticos, de vida corta y que no se les halla fácilmente en el banco de semillas del suelo; pero sus plántulas son capaces de sobrevivir un ario, a veces más, bajo la sombra y responder favorablemente cuando haya la posibilidad de un claro en el bosque. Su madera va de -medianamente liviana a moderadamente pesada (peso específico anhídro de 0,30 a 0,45); la combinación de rápido crecimiento y de madera de aceptables características tecnológicas hace que las HD sean muy interesantes comercial e industrialmente. Varias de las especies de mayor utilización y valor económico están en este grupo, tales como: Cedro (Cedrela odorata), Caoba (Swieténia macrophylla), Peinemono (Apeiba aspera y A. Membranaceae), Arenilla (Qualea spp.), Nogal o Moho (Cordia alliodora), Sajo (Campnosperma panamensis) y Melina (Gmelina arborea), entre otras.
2.2.3. Esciófitas Parciales (EP)
Especies tolerantes a la sombra en las etapas tempranas de su desarrollo, requieren elevadas tasas de iluminación para pasar de las etapas intermedias a la madurez. Soportan la sombra pero no la requieren y aumentan el crecimiento si se abre el dosel del bosque, permitiendo mayor ingreso de luz solar. Su aparato fotosintético se satura a bajos niveles de radiación, aunque tienen la capacidad de incrementar su crecimiento al aumentar la radiación, adaptándose a cambios de RAFA del 1% en el sotobosque y hasta el 20% en un claro nuevo. El ciclo de vida es centenario (100 o más arios), pues debido a su patrón de asignación de recursos los destina principalmente a conformar estructuras más sólidas y duraderas que las de las heliófitas, sobre todo madera y corteza. Las especies de este gremio presentan maderas que van de pesadas a muy pesadas (peso específico de 0,45 a 0,90).
Tienen semillas medianas a grandes, por lo cual la dispersión se hace por corrientes de agua o por animales, principalmente mamíferos. En el patrón de germinación los cotiledones sirven para almacenar grandes reservas alimenticias y permanecen con la plántula durante sus primeras etapas de vida para soportar condiciones adversas, como la sombra o el suelo inundad() (caso típico de Prioria copaifera). Muchas especies tienen germinación epigeal (Ecopaifera, Virola sebtfera, Otoba gracilipes, Brosimun spp.) y con la notable disponibilidad de reservas alcanzan crecimientos impresionantes en la etapa de brinzales (ej. Ecopaifera y Carapa guianensis); en algunas los cotiledones permanecen dentro de la testa, probablemente como protección contra los depredadores. Muchas EP tienen semillas apetecidas por la fauna silvestre, especialmente los roedores, que llegan a consumir cantidades significativas (C. guianensis), lo que sirve como mecanismo de dispersión pues, en épocas de abundancia, cierta fauna como la guagua (Agouti paca), transportan y entierran estas semillas, algunas escapan del consumo, logrando germinar y así ayudan a la repoblación del catival. Para la silvicultura es fundamental saber que las semillas de las EP toleran pocos ambientes de alta temperatura y suelo seco; y a menos que estén enterradas, se ven limitadas para colonizar los claros grandes o sitios expuestos a plena exposición.
2.2.4. Esciófitas Totales (ET)
Especies que requieren sombra y no tienen la capacidad de aumentar significativamente su crecimiento con la apertura del dosel. Muchos de los aspectos de biología y ecología de las EP son válidos también para las esciófitas totales. El número de especies ET es menor que el de EP. En el catival es típico el Caimito (Pouteria caimito). En otra zonas el Ahumado (Minquartia guianensis). Este gremio no es muy amplio.
2.3. Organización y Dinámica Sucesional del Catival
En este sentido es necesario .diagnosticar tres indicadores: la composición florística, la estructura y la dinámica sucesional. La complejidad y naturaleza del catival exigen también su comprensión sistémica y el conocimiento de su riqueza y diversidad. Estos aspectos permiten disponer de adecuados elementos de juicio para proponer mejoras al sistema de aprovechamiento y para justificar y tomar decisiones de manejo silvicultura'. Es una de las más importantes bases ecológicas para su silvicultura. Su estudio es valioso en las sucesiones postaprovechamiento pues el futuro de la conservación, la utilización y el manejo del Prioretum, como el de muchos bosques tropicales, recae en los Cativales Residuales del Aprovechamiento. Un trabajo de Conif sobre fitosociología del Catival Clímax y 21 años postaprovechamiento (1995), así como otros estudios de campo de dicha entidad permitieron obtener la siguiente información al respecto.
2.3.1 Organización del Catival Climax A2
- Vegetación de D.A.P. > 10 cm
Se halló una relativa riqueza florística de especies arbóreas, en contraposición con la característica, monotípica de este bosque en torno al Cativo (Prioria copatfera). Se registran 235 individuos, pertenecientes a 42 especies de 25 familias; predominan Lecythidaceae con Eschweilera garagarae; Gustavia nana; Gustavia superba; Lecythis turiana y Lecythis sp. y Meliaceae con Carapa guianensis, Trichilia martiana y Cedrela sp. Hay pocas especies arbóreas, cada una con gran número de individuos; se destaca Prioria copaifera (100 ind./ ha, seguida del Roble (Tabebuia rosea) y Güino (Carapa guianensis) con 13 ind. cada una. Siete especies agrupan el 90.7% del área basal y 35 sólo el 9.3%; P copaifera es la primera con el 76.97% (31.38 m2/ha) del área basal del bosque. Según el Indice de Valor de Importancia Simplificado -IVIS-, cativo es la especie de mayor peso ecológico (119/300) seguida de Lecythis tuyrana y otras 4 especies que suman el 78.28% del IVIS y dominan el bosque no intervenido.
- Vegetación de D.A.P. a 10 cm
Se presentaron 32 familias botánicas con 57 especies, considerada una alta diversidad. Las de mayor presencia eran Rubiaceae (Condominea; Gonzalogumia y Pentagonia), Annonaceae (Guatteria sp., Guatteria tanduzii, Malmea anomala; Oxandra sp . y Rollinia mucosa),Mimosaceae (gén. Inga, especies semialata; sapindoides, edulís y psittacorum); Piperaceae es menos importante pero bien representada (gén. Piper, especies novogranatense, multiplinervium; reticulatum y umbraculum). Se reportaron 494 individuos de 57 especies. La regeneración de brinzales y latizales era relativamente escasa.
- Vegetación de D.A.P. < 1cm
De alta diversidad en gran cantidad de taxas y variedad de formas botánicas (lianas, hierbas, bejucos, plantas del piso, arbolitos, arbustos y palmas). Se reportan 19.914 ind./ha de 80 especies; predominan "Baba de zaino" (Dieffenbachia longispatha) con 9.212 id., es muy agresiva y se distribuye extensamente sobre el piso formando grupos dominantes; Heliconia brachiantha (3.134 id.) que ocupa grandes áreas abiertas del catival y posee hábito gregario; la "Pita" (Aechmea magdalenense) con 1.952 id., su comportamiento es similar al de Dieffenbachia e inhibe el desarrollo de otro tipo de vegetación. Abundan los helechos, como las Dryopteridoidea "H. Hurtado" (Tectaria incisa) con 128 ind. y "H. Mena" (Tectaria draconoptera Cope!) con 124 ind. y la Polipidiaceae "H. Arroyo" (Pteris propingua Agardn.) con 50 ind. También abundan las S elaginellas (550) como la Oreja de Mula (Philodendron podophyllum). Abunda la regeneración de E copatfera (2.028 brinzales/ ha.), localizada en sitios con significativa apertura de claros, Castilloa elastica (con 548) y Bactris paula Bailey (con 88).
En la vegetación mayor de 1 cm de diámetro se reportaron 80 especies de 37 familias, dominando Rubiaceae; Annonaceae; Moraceae; Lecythidaceae; Piperaceae; Mimosaceae; Arecaceae; Apocynaceae; Chrysobalanaceae; Meliaceae y Papilionaceae. E copaifera tiene área basal de 41.56 m 2/ha., equivalente al 75.55% del total de la del bosque y es la especie de mayor peso ecológico con un 44.90% del IVI total, ratificándose la tendencia del ecosistema a la homogeneidad específica.
El cociente de mezcla es 1:9. El Indice de Shannon-Weaver (H) es de 5.243; equivalente a una alta diversidad del ecosistema. Para la Riqueza Florística mediante el méto'clb AREA-ESPECIE se hallaron 85 especies en 2.500 m 2 ., incluyendo la vegetación baja; y a partir de 1.700 m2. la curva incluye al 92% de las especies, indicativo de una notable riqueza florística.
Verticalmente hay 3 estratos, dos superiores continuos y uno inferior separado de los dos primeros. En el estrato más alto se agrupan las especies dominantes, con árboles de hasta 40 m. de altura y copas de diámetro >15 m. fuertemente condensadas, con alta dominancia de Prioria copaifera y Lechytis tuyrana (D.A.P. >100 cm.), fustes en su mayoría rectos y con buenas características morfológicas, esporádicamente cubiertos por lianas de hasta de 35 cm. de diámetro, como el "Bejuco Chocho" (Entada sp.) y el "Bejuco Don Guandú" (tincaría racemosa). En el piso subsiguiente hay escasos árboles intermedios de altura <30 m, sin formar una gruesa faja árboles con vegetación asociada hasta los 20 m. de altura, de pocas raíces fúlcreas o contrafuertes, hecho curioso, que en cambio es una característica común de la vegetación cliófila intermedia y baja. El piso inferior es de 15 m. de altura, dominad() por Arecaceae. El sotobosque está muy poblado, principalmente por Aechmea magdalenae; Dieffenbachia longispatha y Heliconia brachiantha. Abunda la regeneración natural arbórea. Las hierbas y bejucos representan a 73 especies.
2.3.2. Organización del Catival Intermedio
2.3.2.1. Catival Intermedio - 21 años Postaprovechamiento
- Vegetación con D.A.P. > = 10 cm
Se registraban 382 individuos de 86 especies y 39 familias; la mejor representada es Annonaceae (Guatteria sp.; Guatteria tanduzii; Oxandra panamensis; Oxandra sp; Rollinia mucosa y Unonopsís sp.). seguida de Moraceae (Brosimun alicastrum.; Castilloa elástica y C. tunu; Clarisia racemosa; Pseudolmedia laevigata. y Sorocea aff. Hensi , Meliaceae (Carapa guianensis.; Cedrela sp.; THchilia martiana; T poeppizii. y T quadrijuga) , Mimosaceae (Inga psittacorum; 1. edulis , 1. Sapindoides e I. Semialata, Cynometra martius y Pentaciethra macroloba); de gran importancia las Papilionaceae como Dipteryx panamensis; Dussia sp.; Myroxylum balsamum; Pterocarpus officinalis y Uribea tamarindoides. Las 10 especies más abundantes son Prioria copaifera (con 18 individuos), «Vela» (Rubiaceae N:N:) con 18, Swartzia pinnata y Pouteria caimito, con 15 cada una, C. guianensis, con 13, y C. elastica con 12, Pentaclethra macroloba con 11, y Aspidosperma sp., Brosimun alicastrum y Triplarís americana, cada una con 10 ind.. Estas 10 especies representan el 34.5% de la abundancia, mientras que las otra 76 componen el 65.45%, a veces con menos de 10 ind./ha.
Según su importancia hay tres grupos de especies: el de alto peso ecológico conformado por Anacardium excelsum (22,9% del IVIS), y P. copatfera (12.7%) con alta abundancia y dominancia. El de peso medio formado por Dypterix panamensis (7.4%) con. árboles pero de gran porte, Carapa guianensis (7.3%), C. elástica. (6.7%) y Neea delicatula (6.1%). El tercero, de peso bajo, formado por Pentaclethra macroloba; Pouteria caimito; Aspidosperma sp; Chrysophyllum sp; una Rubiaceae (N.N.) y L. tuyrana (5 al 6%).
- Vegetación con D.A.P. > 1cm. y < 10 cm.
Este grupo tenía palmas, árboles juveniles y arbustos de 93 especies pertenecientes a 39 familias (alta diversidad). Se destacan Annonaceae (Guatteri a chocoensis; G. tanduzii; Malmea anomala; Oxandra panamensis; Oxandra sp; Rollinia mucosa y Unonopsis gracilis), Mimosaceae (Inga psittacorum; 1. edulis; 1. saffordiana; 1. sapindoides, 1. semialata y macroloba), Moraceae (Brosimum alicastrum; B. guianense; Castilloa elástica; C. tunu; Pseudolmedía. laevigata y Sorocea affinis), Lecythidaceae (Eschweilera garagarae; Crías caultflora; Gustavia nana; G. superbaL.,tuyrana), Meliaceae (C. guianensis; Cedrela sp; Trichillia martiana; T poeppigii y T quadriyuga). Las Palmas participan de modo importante (Bactris maraja; B. paula; Crysophila kalbreyeri; Geonoma oxicarpa y Socratea exorrhiza).
 Bosque de cativo en proceso de reconstrucción a los 16 años postaprovechamiento. Riosucio, Chocó. Colombia.
Bosque de cativo en proceso de reconstrucción a los 16 años postaprovechamiento. Riosucio, Chocó. Colombia.
 Apertura de grandes claros para la agricultura migratoria practicada por colonos en el catival. La Balsa, Riosucio, Chocó, Colombia.
Apertura de grandes claros para la agricultura migratoria practicada por colonos en el catival. La Balsa, Riosucio, Chocó, Colombia.
Fueron hallados 1.726 ind. /ha, de 93 especies correspondientes a 39 familias. Por su abundancia se destacan: Sloanea sp. (7.88% del total), Bactris maraja (6.3%), Clavija mezzii (5.3%), Castilloa tunu 80 (4.6%), Gustavia superba (4.2%) y P.' copaifera (3.6%); 28 conforman el 72.5% del total de la población, mientras que las 65 especies restantes conforman el 27.5%. Por do. minancia sobresalen Gustavia superba (7.4% del área basal total), Sloanea sp. (7%) y C. tunu (5.5%). El 86% de las especies tienen pocos individuos, cada uno de escasa área basal.
- Vegetación con D.A.P. < 1 cm
- Para desarrollar un manejo forestal sostenible es importante considerar que, así como el bosque tropical es ampliamente diverso y rico en ecosistemas, comunidades y especies forestales, también su conocimiento biológico debe ser amplio y específico.
- Las bases ecológicas aún son poco reconocidas y aplicadas, pero las exigencias de la utilización de las selvas tropicales conducen a que no pueden seguir siendo soslayadas y al contrario, constituyen una situación álgida a resolver para alcanzar su ordenación, manejo y aprovechamiento sostenible.
- Las bases formuladas teóricas deberán siempre ser contrastadas con la realidad para dilucidar si hay o no suficiente conocimiento de nuestros ecosistemas forestales productores, si son o no aplicadas en la normatividad y en los procesos del aprovechamiento de los bosques naturales.
- El aprovechamiento y manejo sostenible de los bosques tropicales exige incluir y aplicar las bases ecológicas en un enfoque de conservación (mas no de preservación) de su condición y su salud y vitalidad.
- El mantenimiento de la salud y el funcionamiento ecosistémico de los bosques, observando las bases ecológicas durante su aprovechamiento y manejo, permite que la silvicultura sea exitosa.
- El estudio y la implementación técnica y normativa de las bases ecológicas debe ser gradual, ofreciendo a los usuarios y a las autoridades ambientales la informaciórla educación y sensibilización para que el proceso sea exitoso y no se convierta en una nueva talanquera para el desarrollo del potencial forestal nacional.
- Muchos de los estudios pertinentes, por ser de mediano y largo plazo y de alto costo y pueden ser ejecutados, por ejemplo, mediante cooperación entre Conif, los Institutos de Investigación del Sistema Nacional Ambiental, algunas Universidades, y empresas forestales, entre otros.
- La aplicación de los resultados de los anteriores estudios deben permitir un ajuste gradual de los métodos de cosecha, la planificación de aprovechamiento y los planes silvícolas.
- El estudio y la implementación de los mutualismos en el manejo forestal deberá orientarse en función de incentivar el establecimiento de la regeneración e inducir la produción de semillas y renuevos mediante árboles portagranos.
- Es necesario realizar y disponer de los estudios directos de la fauna más importante asociada con los bosques objeto del manejo y aprovechamiento, principalmente de aquellos grupos taxonómicos (en mamíferos, aves e insectos) referenciados en este trabajo; con énfasis en su autoecología con respecto a la fenología de las principales especies vegetales.
- Se recomienda estudiar y mejorar la descripción de los órganos de la reproducción de, por lo menos, las especies arbóreas más importantes para la silvicultura y el aprovechamiento.
- Se recomienda tomar medidas de prevención y mitigación del impacto de las actividades de aprovechamiento y manejo forestal sobre la fauna silvestre, especialmente con respecto a aquella involucrada en la ecología de las especies arbóreas de mayor importancia socioeconómica.
Este grupo es el más diverso; constituido por bejucos, hierbas, herbáceas, rastreras, brinzales, arbustos, aráceas y líquenes y musgos. Se registraron 15.924 ind./ha de 118 especies; las más abundantes eran: C. elastica (4.906 ind. /ha.) y "Selaginella" Selaginella arthritica Aiston (2.214) de población localizada pero muy abundante. Entre la regeneración natural temprana arbórea sobresalen: Quararibea asterolepsis (574 ind./ha), Dypterix panamensis (274), Gustavia superba (168), Inga semialata (180), Oxandra sp. (306), Pouteria. caimito (192), Hirtella racemosa (200) y Posoqueria latifolia (288). Es notable el papel ecológico de las palmas: en el estrato bajo se destacan Astrocaryum malibo (374 palmas/ha), Geonoma oxicarpa (326) y Chamaedorea sp. (250), distribuidas uniformemente y frecuentes en el piso; de menor presencia son Socratea exorrhiza (62), Crysophila kalbreyeri (20) y Bactris maraja (56). También son importantes los bejucos: «Bejuco tortugo» Hippocratea sp. (586 ind./há.), «B. hombresolo» Paullinía sp. (106) y Cortada sp,. (escaso pero con estructuras de hasta 35 cm. de diámetro. Igualmente los helechos Hurtado Tectaria incisa (400 ind/ha), Clavellino Aspienium delitescens (346), Moreno Diplazium gradifolum (466); Tectaria draconoptera «H. mena» (132), Pterís propingua «Helecho arroyo» (42) y Adiantum tetraphyllum «Helecho porrancho» (44).
Según Area Basal, dominan Anacardium excelsum (20.8%) con pocos individuos corpulentos remanentes de la explotación) y Dipteryx panamensis (6.3%), C. guianensis (3.7%) e Hirtella racemosa (4.2%). Por peso ecológico sobresalen: A. excelsum (10.5% del total del TVE), Ecopaifera, abundante y de alta AB (5.8%), Sloanea sp. (3.6%), D. panamensis (3.4%) y Bactris maraja (2.7%). El cociente de mezcla es 1:17. A pesar de su alta diversidad este catival está evolucionando a la homogeneidad, con características de ecosistema secundario tardío. El índice de Shannon-Weaver (H) es de 5.707, indicador de alta diversidad florística (1.208 individuos de 118 especies). El análisis de la Riqueza Florística, mediante la curva especie-área en 2.500 m 2., señaló que 1.700 m2, es el área mínima de expresión biológica del ecosistema pues allí se reúne el 93% del total de las especies muestreadas, exceptuando briófitos.
Es importante la cantidad de pequeños claros en evolución; producidos por la caída, natural o antrópica, de plantas de gran tamaño, con troncos y ramas aún en descomposición. La disposición de las frondas de las palmas les perfflite actuar como canales recolectores de los detritos (hojarasca). Los árboles presentan raíces fúlcreas y a veces, contrafuertes. Las lianas proliferan sobre el piso presencia típica de aguijones bien desarrollados. Se reportan especies herbáceas indicadoras de ambientes húmedos, como Seloginelba onceps y Piper multiplinervium.
En el catival A2, a los 21 arios de aprovechado, las especies más importantes del bosque clímax ya han recuperado buena parte de su estatus fitosociológico y que están ocupando posiciones estratégicas dentro de la organización del ecosistema y que, a la vez, otras están en el proceso de ganar espacios típicos para su establecimiento. También se infiere que en esta edad postaprovechamiento el bosque está finalizando su etapa más dinámica de reconstrucción y que se puede estar iniciando su proceso de homeostasis. La condición de este catival de 21 arios no es adecuada para realizar una cosecha de madera de Cativo, Sande, Virola o Güino; su condición para iniciar tratamientos o manejo silvicultura' tampoco es adecuada pues requeriría una intensiva aplicación de capital, trabajo y tecnología. Ya se ha pasado el momento de hacer manejo silvicultura' para obtener un catival orientado a. la producción; la etapa de desarrollo del bosque para hacerlo debe ser de una edad anterior a 21 arios postaprovechamiento. Ahora, si el enfoque de un análisis es para sólo conservación del ecosistema, la conclusión es que al cabo de este tiempo la reconstrucción del bosque ha sido bastante favorable y que la incidencia del aprovechamiento en el largo plazo es mucho menos negativa que recién que éste se ha llevado a cabo.
2.3.2.2. Catival Intervenido - 16 Años Postaprovechamiento
Conif (1993) realizó una investigación de campo durante 6 arios (1982-1987) en un catival tipo A2 que había sido aprovechado en 1977 por una empresa productora de tríplex; el estudio se hizo general, no por grupos de tamaño y sólo para árboles, con estos resultados:
- Riqueza y Diversidad Florística
Se registran 68 especies de 26 familias; Caesalpinaceae y Fabaceae con 5 y 4 géneros cada una, seguidas de Annonaceae, Lecythidaceae, Mimosaceae y Moraceae (3 gén. cada una). Sólo 21 especies se encontraban simultáneamente en las categorías de tamaño brinzal, latiz,aly fustal. Dominan Caracolí, Cativo, Guasco, Pacó y Dormilón. Se hallaron 53 especies en el estado de brinzal, 52 en latizal y 27 en fustal; la participación de las palmas fue casi nula.
El 88% del peso ecológico lo suman 9 especies. Se destaca el cativo (44% del total), abundante y con árboles de buen porte; están poco representadas güino (5%) y virola; el caracolí alcanza alto peso ecológico con muy pocos individuos pero de grandes dimensiones (esta especie no es aprovechada). Por familias, Caesalpinaceae lleva el 48% del peso ecológico, mientras que Fabaceae, Lecythidaceae y Anarcadiaceae suman el 25%; otras 22 familias alcanzan solo el 27%. Por abundancia, Caesalpinaceae reunen el 39% de la regeneración, seguida de Lecythidaceae (13%), Fabaceae (12%) y Mimosaceae (9%). Tendencia similar ocurre para la dominancia. El Area Basal total es de 35.8 m2/ha, superior al rango óptimo promedio (19-23 m2/ha) establecido para bosques tropicales. La variación de la regeneración (densidad y composición) y de la estructura diamétrica durante varios años no es significativa, como se aprecia en la Tabla 1.
Más del 70% de la población se encuentra en la clase 60 cm y parte de ella es remanente del aprovechamiento industrial. La distribución de valores de AB en las clases diamétricas de 10 a 60 cm es muy pobre (menor a 3 m2/ha).
En cuanto a la distribución del volumen, este estado sucesional sigue un patrón semejante al relacionado para el área basa', como se aprecia en la Tabla 2.
Para D.A.P. > 10 cm. el Indice de Similaridad Florística de Sorensen es del 27 y el 54%, que indica una relativa heterogeneidad en composición florística; y que es inferior al del Clímax; el Cociente de Mezcla (C.M.) es de 1:3,5 (mezcla intensa de especies); para brinzales el cociente es 1:12 (mayor homogeneidad) y para latizales de 1:3.
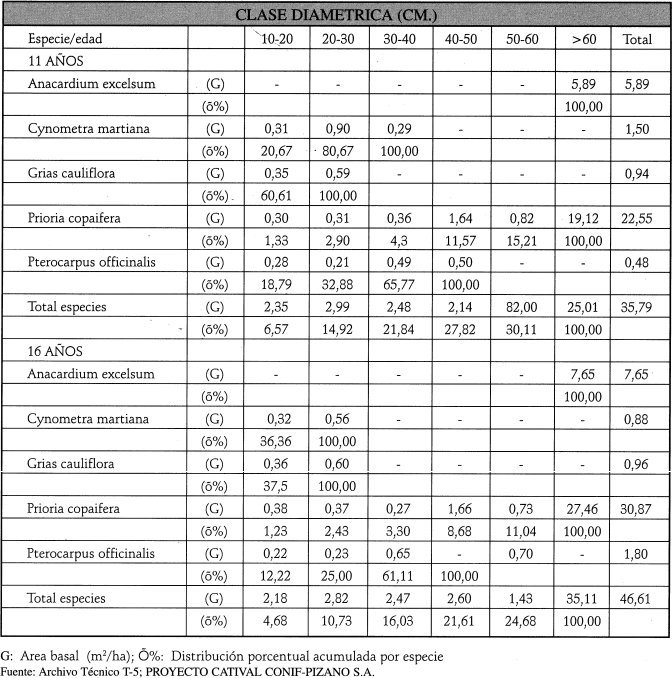
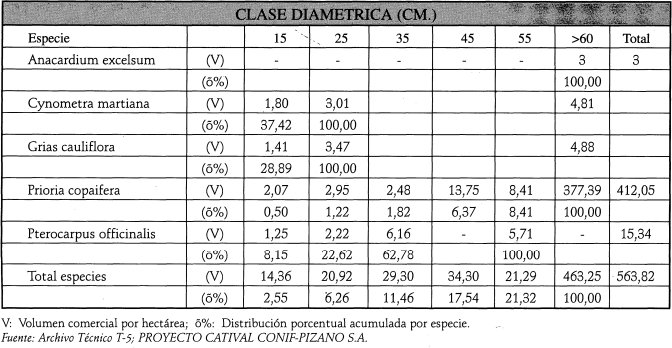
A los 16 años postaprovechamiento, en el catival A2 hay un hecho de gran importancia silvicultural consistente en que el peso ecológico (IVIS) del cativo se incrementa al pasar de una edad sucesional a otra, situación que es muy favorable para planificar los trabajos del manejo forestal. También se destaca la dominancia total de especies heliófitas durables y esciófitas parciales, hecho que indica que el bosque ya se encuentra en un estado de sucesión avanzado. La abundancia de la regeneración de brinzales y latizales de las especies comerciales garantiza el abastecimiento continuo de plántulas del futuro vuelo, las cuales deben ser estimuladas y asistidas para promover su desarrollo y disminuir las tasas naturales de mortalidad. Casi todas las especies de mayor interés para el aprovechamiento y el manejo pertenecen a las familias dominantes en esta etapa sucesional del catival (por ej. Caesalpiniaceae). A los 16 arios de reconstrucción hay una hetereogeneización en la composición florística y la estructura diamétrica tiende a ser similar a la del catival no intervenido.
En esta edad, todos los indicadores ecológicos del catival A2 señalan que podría ser una edad o etapa tardía para iniciar un eventual manejo silvicultural en bosques residuales del aprovechamiento, según los mismos elementos analizados para el caso de 21 años postcosecha
2.3.2.3 Catival Intervenido - 10 Años Postaprovechamiento
La población de brinzales es de 10.700 ind./ha aprox., similar que en catival O arios; sin embargo, Prioría copaifera abarca el 14.3%, que es el doble que en dicha edad. Caracolí es la especie predominante (30.4%), casi igual que en O arios. Los géneros Gustavia y Grias (Pacós) también son importantes (8.7%), similar que en O arios; en contraste aparece una nueva e importante regeneración de Lecythidaceae (especialmente Eschweilera) que conforma el 11.5% del total de brinzales. En esta fase la participación de heliófitas efímeras y heliófitas tardías (Cecropia spp.) es menor y conforma sólo el 0.003% de la comunidad vegetal arbórea. Los latizales se han reducido a la mitad, respecto al año O. El Cativo tiene poca presencia pues conforma sólo el 0.8%.
Fueron registrados 242 fustales/há y cativo es la especie más abundante (20.7%), una buena parte remanente del bosque primario, con D.A.P. promedio de 42 cm.; le siguen las heliófitas Trema micrantha (19%)y Cecropia spp. (9.2%), establecidas en grandes claros originados por la cosecha maderera; después vienen especies del clímax como Gustavía y Crías (9.8%), higa spp. (8.4%), Anacardium (3.3%) y Eschweilera (2.4%).
En este tamaño Prioria copaifera tiene el mayor peso ecológico (IVIS 73.9); seguida de Anacardium (58.9) y Lecythis tuyrana de alto peso ecológico por sus árboles maduros réffianentes del aprovechamiento). De 51 árb./ha. de cativo hallados, 37 tenían menos de 50 cm de D.A.P. y todas las especies presentan distribución diamétrica típica o normal.
En esta edad la población de heliófitas efímeras ha empezado a descender y ha perdido rápida y gradualmente su importancia ecológica, en comparación con la sucesión de 5 y O arios y el dosel se está cerrando nuevamente; igualmente, el bosque ha terminado la parte más activa de su recontrucción dinámica. Se han empezado a dar las condiciones y los recursos para el establecimiento y el mejor desarrollo de las especies del gremio de las esciófitas parciales, como el caso del Cativo, de alto interés pata el manejo. Aquí se presentaría una condición silvicultura' mínima para iniciar la aplicación de tratamientos.
2.3.2.4. Catival Intervenido - 5 Arios Postaprovechamiento
El Coeficiente de Mezcla es de 1:32 (tendencia a una relativa homogeneidad florística); la abundancia de P. copaifera y de Cecropia spp. y Trema micrantha es menor que a los 3 y O arios postaprovechamiento. En la Riqueza Florística se destacan Cecropia peltata (9.4% del IVIS) de alta abundancia y distribución irregular; Inga spp. (6,3%), Schweilera spp. (5.7%), ambas con alto número de individuos y Prioria copatfera (5,3%) con alta Area Basal (AB). Menos importantes son Castilloa elastica, Ficus insípida, Spondias mombin yLecythis tuyrana, cada una con el 3 a 4%. El 62.4% del IVIS lo representan 17 especies y el 37.6% otras 38
Algunos indicadores de la estructura y la diversidad del catival a los 5 arios se anotan a continuación, con base en estudio de campo realizado para tal fin, empleando tres parcelas de muestreo en el Bajo Atrato, Riosucio — Chocó. Algunas características fitosociológicas del catival A, 5 años postaprovechamiento*

Se registraron 3.363 brinzales/ha de Gustavia y Crías (18.8%), Anacardium excelsum (12.6%), Lecythis tuyrana (6.9%) y Priora copatfera (5.9%). También hay pocos brinzales (30% menos que en otras edades) tal vez por la proliferación de Heliconias que impiden su establecimiento y desarrollo. Se hallaron 1.796 latizales/ha, similar que bosque cero arios; predominando Cecropia membranaceae (25.6% del IVI), Gustavia y Grias (8.7%), Prioria copaifera (7.5 ()/0) que a esta edad tiene poco peso ecológico, seguida de Lecythis tuyrana y Anacardium excelsum. Se encontaron 160 fustales/ha, de Cecropia\membranaceae (27.6%), Prioria copaifera (16.7%), Cynometra longifolia (9%), Gustavia superba, Crías cauliflora y Anacardium excelsum.
 Arbol de sande (Brosimun utile) talado y troceado. Esta actividad origina claros en el bosque, que dinamizan la regeneración natural.
Arbol de sande (Brosimun utile) talado y troceado. Esta actividad origina claros en el bosque, que dinamizan la regeneración natural.
 Bosque de Catival (Prioretum copaiferae) en quema por colonos para cambio de uso del suelo. Aquí la reconstrucción natural es casi irreversible. La Balsa, Riosucio, Chocó, Colombia.
Bosque de Catival (Prioretum copaiferae) en quema por colonos para cambio de uso del suelo. Aquí la reconstrucción natural es casi irreversible. La Balsa, Riosucio, Chocó, Colombia.
A los 5 años postaprovechamiento, la cantidad y la calidad de la regeneración natural deseable del Catival son menores que a los 10, 16y 21 arios. Había una población de árboles en su mayoría de especies de valor económico actual restringido. La composición e importancia de brinzales, latizales y fustales de especies deseables como Cativo, Güino, Virola y Sande indican una indeseable condición silvicultura' para iniciar un proceso de manejo. Sucesionalmente, a los 5 arios postaprovechamiento el catival se halla en una etapa muy dinámica de reconstrucción (fase de sucesión secundaria temprana).
2.3.2.5. Catival Intervenido - 3 Años Postaprovechamiento
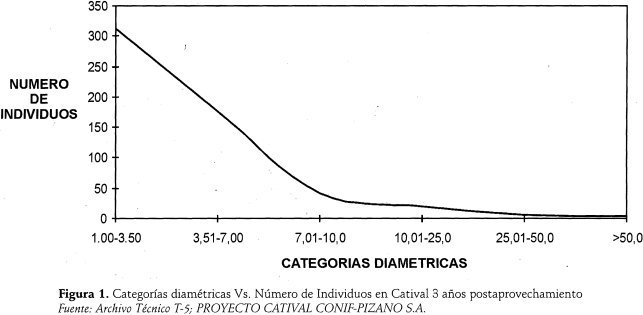
Su Coeficiente de Mezcla (CM) es de 1:13 (diámetro >1 centímetro de diámetro); similar que en 5 arios, por la abundancia de especies heliófitas efímeras y de cativos residuales. La Riqueza Florística mostró 43 especies de 25 familias, relativamente baja comparada con la del catival clímax. Dominan AnaCardiaceae, Moraceae, Lecythidaceae y Caesalpinaceae. Las familias mejor representadas eran Anacardiaceae, Caesalpinaceae y Lecythidadeae. El análisis conjunto para todas las categorias de tamaño señala a 10 especies como las más importantes, hallándose 1.552 ind./ha. (D.A.P. > 1 cm.) que abarcan el 72,6 % del IVI total. Las especies de mayor peso ecológico son Anacardium excelsum (20)%, con pocos fustes residuales de gran tamaño), inga edulís (11,2%), Cecropia peltata (10,2%) y Schweilera sp, (9,8). Un poco menos importantes eran Pterocarpus officinalís, Grias cauliflora, Cynometra martiana y Prioria copaifera Spondias mombin y Trichilia cuadrijuga. Todas estas especies son dominantes en la estructura del catival climax pero, para la etapa de 5 años, ocupan posiciones sociológicas secundarias. Otras 33 especies suman el 27,5%. Prioria copaifera era la especie de mayor abundancia de brinzales y latizales, localizados sobre todo en claros. Como lo indica la Figural, la estructura diamétrica hallada indicó que existía una relación inversa (a menor diámetro mayor número de individuos y viceversa); la curva en "J " invertida mostraba que aún había un relativo equilibrio en el sitio.
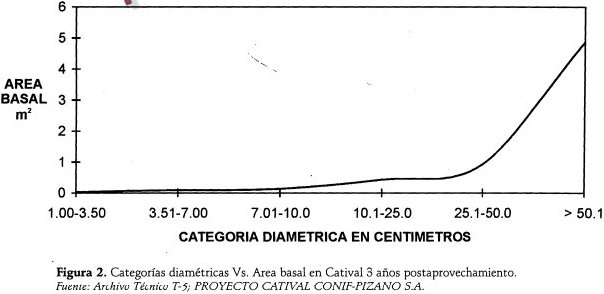
Igualmente, y de acuerdo con la Figura 2, la relación Categorías Diamétricas vs. Area Basal muestra la tendencia de una relación directa entre el diametro y la área basal, es decir que a mayor diámetro mayor área basal, cuya curva en forma de "J" invertida señala también un equilibrio estructural relativo.
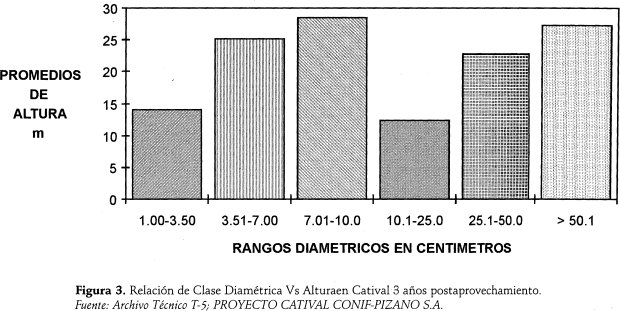
Según la Figura 3, la relación Categorías Diamétricas vs. Alturas muestra dos picos, correspondientes a dos máximas alturas promedios de 28 y 27 metros repectivamente; comportamiento debido, posiblemente, a la incidencia de los árboles remanentes y de los que se elongan tras la apertura de claros.
La cosecha selectiva deja una serie de árboles de tamaños superiores e inferiores a los límites diamétricos de corta que, junto con la nueva regeneración de los claros generados en el catival, origina una mezcla de arreglos estructurales con diferentes microfases sucesion- ales dentro del ecosistema. En los claros provocados por el aprovechamiento se desarrollan especies que determinan cierta dominancia transitoria como la especies heliófitas efímeras (tipo Cecropia peltata), pero que paulatinamente disminuyen hasta casi desaparecer (ver sucesión 10 arios postaprovechamiento). Las poblaciones de brinzal, latizal y fustal joven de especies arbóreas deseables son consideradas insuficientes en términos de su potencial de reserva como reemplazo de las poblaciones de mayor tamaño; además, se considera que están aún compitiendo por lograr establecerse y superar la etapa crítica de sobrevivencia, debido a que el sitio está influenciado por fuertes factores abióticos limit'antes como los períodos secos fuertes y prolongados, sumado a las quemas causadas por el hombre, junto con los periodos de lluvias y posterior inundación, prolongado hasta 9 meses al ario; a lo que se le suman los desequilibrios introducidos por las operaciones de cosecha.
Por todas estas razones la iniciación de un manejo silvicultura intensivo en la etapa 3 arios postaprovechamiento no es aún factible ni razonable técnica o biológicamente.
2.3.2.6. Catival A2 recién intervenido por aprovechamiento — año 0
A los 6 u 8 meses postaprávechamiento la Riqueza Florística muestra 22 especies de 14 familias (que se estima pobre). El C.M. es 1:5.5 (fuerte tendencia a la hetereogeneidad). Estructuralmente hay 12 especies que según su I.V.11 son las más importantes: Cecropia peltata con el 27.4% del IVI, alto valor debido a su abundante población, Grias cauliflora (9.9%), Tiplaris cumingiana (8.9%) y Spondias mombin (7.2%). De mediana importancia son Nectandra reticulata (6.9%), Prioria copaífera (5.7%) y Pterocarpus officinales (5,5%) Tapirira meryantha (4.4%), Aspidosperma marcgravianum (4.2%) y Rheedia edulis (3.4%). De baja importancia ecológica son Clavija mezzii y Trichilia quadrijuga.
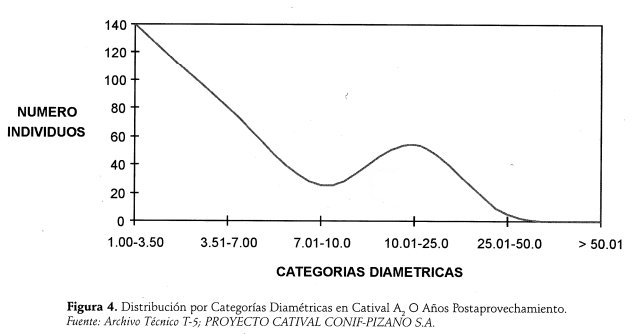
La relación Categorías diamétricas vs. Número de árboles muestra una curva con tendencia de "J" invertida, con un pico en el rango 7 a 25 centímetros, debido a la afluencia de un mayor número de árboles en dicha categoria de tamaño, remanentes del aprovechamiento (ver Figura 4.)
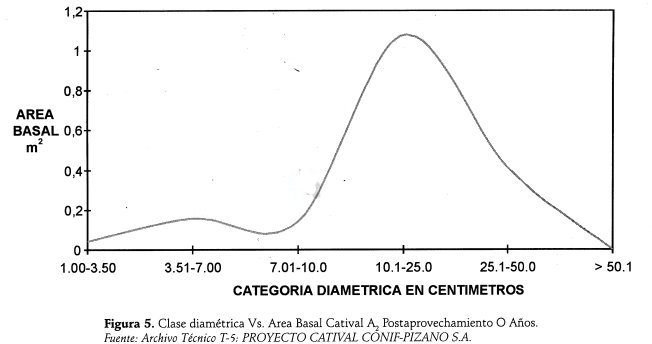
La curva Categorías Vegetales Vs Areas Basales, que se observa en la Figura 5, presenta concentracion de áreas basales en el rango de 7.0 a 50 cm., debido a la afluencia de árboles remanentes del aprovechamiento; esto es una buena serial pues significa que los árboles de este grupo serán los sustitutos de los ya cosechados y/o de los sobremaduros en decandencia.
A los 0 años postaprovechamiento industrial el catival, A1 y A2, presentan, a pesar del impacto negativo de las operaciones de cosecha, un A los O años postaprovechamiento industrial el catival, A1 y A2, presentan, a pesar del impacto negativo de las operaciones de cosecha, un estado relativamente favorable púa su reconstrucción satisfactoria en un largo plazo; sin embargo, este estado postaprovechamiento podría ser mejor, y la recostrucción más rápida y positiva, si las operaciones de cosecha ocasionaran un impacto aún menor. Para un eventual manejo silvicultural orientado a las especies actualmente comerciales (Cativo, Sande, Virola, Güino y otras) es más favorable la condición año O del catival A 1 que la del A2 . Sin embargo, la estructura y la dinámica del ecosistema en esta fase no favorecen ni hacen aconsejable iniciar un plan silvicultural, sino que debe permitirse al bosque un avance de, por lo menos, 10 años en la reconstrucción.
Por último, es necesario enfatizar en que los elementos ecológicos tratados en esta sección, referentes a las características estructurales de los Cativales Intervenidos son de suma importancia para planificar, decidir y hacer la silvicultura de los bosques de Cativo que, como también ya se expresó, son el futuro de la conservación y el manejo sostenible de este recurso forestal.
2.4. Asociaciones y Consociaciones del Ecosistema Catival
En el catival clímax existen asociaciones y consociaciones vegetales; siempre dominadas por Prioria copaifera. En los llamados Cativales A1 predominan consociaciones, con preponderancia del cativo > 50%, y estos bosques se denominan Prioretum copaiferae. En los A2 y A3, son más comunes las asociaciones, con dominancia de P copaiferae pero < 50%, unida a otra especie también de alto peso ecológico, bien sea el Caracolí, el 011eto o el Choibá, entre otros, casos en los cuales se forman las asociaciones Prioríoanacardietum, Prioriolecythietum y Prioriodipteryetum, respectivamente.
La aplicación de estos dos conceptos se fundamenta en que en que cada forma de asociación o de consociación implica significativas diferencias ecológicas que sensibilizan los planes de silvicultura y aprovechamiento, al igual que los planes de conservación de la biodiversidad; como estos planes no son recetas para aplicar indiscriminadamente, se requiere flexibilizarlos y adecuarlos según cada caso y por lo tanto, tener un conocimiento cartográfico, biológico, físico y silvicultura específico para cada una de las formas de agrupación de las comunidades vegetales del catival.
2.5. Los Mutualismos
Los mutualismos constituyen uno de los aspectos biológicos menos estudiados y aplicados en el aprovechamiento y el manejo de bosque tropical, aún a pesar del papel clave que la polinización y la dispersión de semillas desempeñan en su reconstrucción y en la producción de la regeneración natural de las especies objeto del manejo y la cosecha forestal. Para la silvicultura se debería considerar prioritario abordar el estudio de los mutualismos de la relación plantas-animales, sin que ello signifique que sea el más importante.
Para el catival, se han tomado como base los conocimientos existentes sobre la ecología vegetal y animal de este ecosistema, generados por universidades, científicos, centros de investigación y otras fuentes, a los que se les ha integrando con los resultados de las investigaciones del proyecto Conif — Pizano S.A. Por las razones antes expuestas, la información es precaria y los resultados presentados son muy preliminares y se recomienda ahondar en ellos para llegar a disponer de bases más sólidas. No obstante las anteriores circunstancias, se considera que esto no es limitante para, desde ya, empezar a tenerlos en cuenta dentro de las propuestas y planes de manejo del bosque tropical. Aquí se abordan dos de los mutualismos más importantes: la polinización y la diseminación de semillas.
2.5.1 La Polinización
La gran mayoría de las especies tropicales son autoincompatibles, lo que genera problemas para el manejo forestal pues es posible que la cantidad de árboles portagranos que se dejan en el catival u otro bosque luego de aprovechado, y la distancia entre ellos, no sean suficientes para garantizar la suficiente polinización y el éxito en la producción de semillas para la regeneración natural. Los siguientes apuntes han sido obtenidos indirectamente, mediante la consulta de estudios, biológicos realizados en las zonas de catival y por deducción al comparar características de las flores de las especies algunas típicas del ecosistema y los resultados deben ser confirmados por estudios específicos.
- Polinización por Abejas (Hymenoptera: Apoidea)
Las abejas son los polinizadores más importantes del neotrópico (Finegan, 1993) sobresaliendo las familias Anthophoridae (abejas grandes y solitarias) y Apidae (abejas pequeñas a grandes, sociales); éstas prefieren las flores zigomórficas, como las de las familias Papilionoideae, Orchidaceae, Scrophulariaceae y Gesneriaceae, aunque también polinizan un rango muy amplio de otras estructuras. En el catival, varias especies de Papilionoide ocupan posición sociológica predominante y algunas son de interés comercial, bien sea actual o potencial; como Andira inermis, Myroxylon balsamum, Pterocarpus officinalis yP rohrii,Dipteryx panamensis y Dussia lehmanniii. Según Kevan y Baker (1983), las abejas pequeñas tienen tendencia al uso de flores actinomorfas como las de Cativo, Virola (Virola sebifera y V aff. surinamensís), 011eto (Lecythis tuyrana), Güino (Carapa guianensis), Ceiba bonga (Ceiba pentandra) y Caracolí (Anacardium excelsum); también podría inferirse una importante relación con las abejas grandes, que están entre los polinizadores más importantes de los árboles del dosel superior.
- Polinización por Escarabajos (Coleóptera)
Los escarabajos prefieren flores con forma típica de taza pues no ofrecen obstáculos para el ingreso del cucarrón; los adultos se alimentan del polen, de modo que las flores polinizadas por cucarrones pocas veces ofrecen néctar. Son de los polinizadores más antiguos y actualmente se les considera vectores importantes en árboles tropicales, especialmente de las familias Arecaeae y Moraceae, ampliamente representadas en el catival, y en plantas de Araceae y Cyclanthaceae, herbáceas de este ecosistema.
- Polinización por Mariposas (Lepidóptera)
Una proporción importante de mariposas diurnas o nocturnas son vectoras. Las diurnas muchas veces tienen atracción por los síndromes florales de colores encendidos y estructuras pequeñas, generalmente en forma de trompeta de, tubo muy estrecho (que impide el acceso de abejas) sostenida verticalmente; De Vries (1987), citado por Finegan (1993), halló que algunas Rubiaceae como Warsewiczia coccinea (especie típica del catival) y Cephaelis spp. (Ipecacuana) son polinizadas por mariposas diurnas. En los ecosistemas boscosos las mariposas nocturnas son más numerosas e importantes que las diurnas como polinizadoras; así, se distingue la familia Sphingidae por su nivel de especializa ción en la función polinizadora y por la cantidad de especies vegetales de las cuales es vectora; estas mariposas están muy relacionadas con el síndrome del tubo, típico de las Rubiaceae; Posoqueria latifolia, conocida como Huevo de Tigre en el catival, es polinizada por este grupo de mariposas. En el catival también existe Posoqueria pittieri (Jagua) que tiene el mismo síndrome floral e, igual, seguramente es polinizada por Sphingidae.
Las Sphingidae también están asociadas al síndrome de cepillo (flores tubulares, pequeñas y dispuestas en grupos densos con una sola flor central nectífera, cada uno de de los cuales se comporta como una sola unidad de polinización; los estigmas y los estambres están en posición muy salida respecto al resto de los órganos florales); la mariposa, al recoger el néctar de la flor central, también recoge el polen que se pega a su cuerpo. Es típico en especies de Mimosaceae como los Guamos (Inga edulis, 1. sapindoídes e I. Semialata) y el Dormilón (Pentaclethra macroloba) y algunas Tiliaceae como Luehea seemannii; que son dominantes en el catival.
- Polinización por Aves
Hay aves polinizadoras especializadas (alimentación a base de néctar floral) y oportunistas (el néctar es componente de una alimentición variada). Los colibríes (familia Trochilidae) son polinizadores especializados y conforman el grupo más importante de este tipo en el trópico americano (Stiles y Skutch, citados por Finegan, 1993); ellos prefieren el síndrome de flores tubulares (el néctar en el fondo del tubo y los órganos sexuales se ubica de modo que al tomarlo el ave, también recoge el polen y luego lo pasa de una flor a otra; las flores son de colores encendidos, comunmente rojo, anaranjado y amarillo; es más común entre las epífitas (Bromeliaceae), las familias Rubiaceae y Acanthaceae y el género Heliconia; de estas taxas vegetales, los géneros Warscewiczia y Cephaelis y las especies Heliconia brachianta y H. imbricata son típicas del catival, en el cual desempeñan importantísimo papel en el desarrollo de la sucesión luego de la apertura de claros por aprovechamiento, cambio de uso del suelo o fenómenos naturales. Las aves de la familia Icteridae son más de carácter oportunista.
Según Urdaneta (1995), existen en el Catival once (11) especies de la familia Trochilidae del grupo tráfico Nectarívoro- Insectívoro: Anthracothorax Chrysolampis mosquitus; Colibri delphinae; Damophila julie; Florisuga mellivora; Glaucis hirsuta; Heliothryx barrotí; Lepídopyga coeruleogularis; Phaethornis anthophilus yE longuemareus y Thalurania furcata; la riqueza de especies de este grupo aviario en el Prioretum determina amplias posibilidades de que estén involucradas en diversos mutualls.mos de polinización. El trabajo de Orozco (1995) permite concluir que el potencial de aves polinizadoras en el Catival equivale al :51% del total de las especies de avifauna.
- Polinización por Murciélagos (Chiroptera)
Los síndromes florales son de cepillo y de flores grandes acampanuladas; el murciélago re. , volotea como una esfíngida o como colibrí y se cubre el pecho con polen al meter la lengua para chupar néctar. Los arboles polinizados por murciélagos tienen inflorescencias sostenidas en pedúnculos largos y gruesos, que sobresalen completamente de la planta dándole mayor acceso al animal, como en A. excelsum, Pentaciethra macroloba, Prioria copaifera, Lecythis tuyrana, Parkia velutina y P. pendula. Según Rodríguez et al., (1995), en el catival dé Colombia existen, entre otros, dos grupos taxonómicos de murciélagos: la Subfamilia Phyllostomineae (Lonchorhina aurita, Phyllostomus discolor y E hastatus, Lonchophylla thomasi y L. mordax) y Subfamilia Glossophaginae (Glossophaga soricina, G. commissarisi, Lionycteris spurrelli, Anoura geoffroyi, Lichonycteris obscura). De hábitos marcadamente nectarívoros; y que son reportados por Einsenberg, (1989), como vectores de polinización en los ecosistemas boscosos desde Costa Rica hasta Panamá. En el catival, la Bonga o Lano (Pseudobombax septenatum) es polinizada por Chiroptera (Echavarría y Varón, 1988), probablemente del grupo Glossophaginae.
Lo anteriormente expuesto plantea serios e interesantes retos para la silvicultura; por ejemplo, determinar los agentes polinizadores de las especies objeto del manejo para protegerlos y facilitar la regeneración natural de éstas; también, establecer medidas de prevención y mitigación por eventuales daños a la fauna clave en los procesos de polinización, de modo que sea sustentable el funcionamiento del ecosistema y que, por la eventual desaparición y disminución de sus poblaciones, a la vez se ponga en peligro la conservación, regeneración y productividad de aquellas especies (maderables o no maderables) de interés para el silvicultor, en particular, y del manejo forestal en general.
2.5.2 Dispersión de Semillas
El síndrome ,de diseminación por animales con provisión de alimento puede ser el vector más común e importante en el catival; esta dispersión por animales está muy ligada con la frugivaria. Los mamíferos son vectores (mamocoria) de especies con frutos grandes a medianos, color anaranjado, amarillo, café o verde y protegidos por una cáscara, que por su tamaño son aptos para la capacidad visual y su morfología. Las aves son vectores (aviocoria) de especies con frutos pequeños, sin protección y de color rojo, negro, blanco, morado o de más de un color. Con base en la información de Linares, (1988), y de Rodríguez et al. (1995), se determinan algunas especies de mamíferos y aves potencialmente relacionados con frugivoria y dispersión de semillas en el Catival (Tabla 3).
En los bosques tropicales existen las ESPECIES VEGETALES CLAVES cuya función e importancia tienen que ver con que gracias a ellas muchas especies de fauna no sufren ni carecen de alimento pues su producción de frutos es predecible y casi permanente. Entonces, cuando las especies vegetales específicas a su hábitos no se hallan en cosecha, las claves desempeñan dicho papel. En el catival, por ejemplo, es el caso de Ficus paraensis, E insípida, E dendrocída y Ficus spp. De este modo se deriva la necesidad de considerar como como recursos estratégicos a estas y otras especies más del ecosistema forestal, para manetener el funcionamiento del bosque bajo manejo y así mismo, incrementar su productividad biológica y económica.
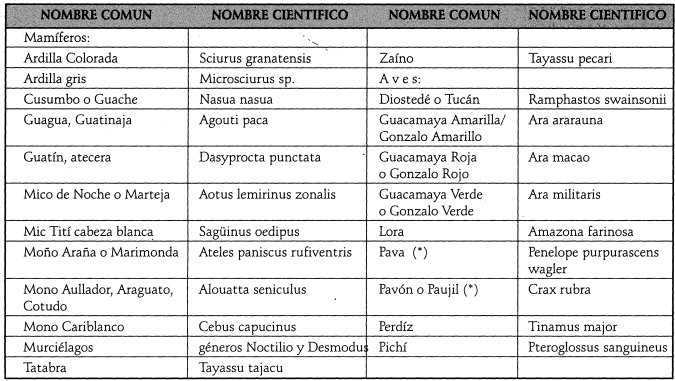
La calidad nutritiva de los frutos es vital para identificar vectores de diseminación de semillas; en este sentido, en el catival son comunes e importantes especies de Annonaceae, Meliaceae (géneros Cedrela y Guarea); Myristicaceae (gén. Virola y Otoba) Lauraceae y Lecythidaceae (gén. Couroupita, Lecythis y Eschweilera)que poseen frutos y semillas de ALTA CALIDAD NUTRITIVA, ricos en grasas y proteínas.
Es así como en algunos sistemas silviculturales "clásicos" se acostumbró a realizar refinamientos indiscriminados eliminando muchas especies arbóreas dentro del erróneo concepto de "indeseables", según su valor comercial, pero importantes por el valor estratégico o nutricional antes mencionado. La nueva perspectiva de la biología dentro del manejo forestal sostenible implicará gradualizar y reconsiderar la intensidad de eliminación de dichas especies para evitar alteraciones significativas a la salud del ecosistema.
- Dispersión por Mamíferos
Hay tres grupos involucrados: 1°) Rodentia (Roedores): con el ratón de espina, Heteromys sp., que es un animal granívoro habitante de un complejo sistema de túneles donde almacena semillas en grupos concentrados, enterrándolas o en fosos poco profundos, y como no llega a consumir todo lo almacenado, una parte germina y así se efectúa la diseminación. Igual sucede con el ratón semiespinoso, Proechimys semispinosus, que vive en túneles y troncos huecos caídos. 2°) Dasyproctidae tiene tres géneros frugívoros, pero en el catival sólo hay dos: Agouti, representado por Agouti paca, Guagua o Guartinaja, y cuyo papel dispersor es limitado pues no almacenador sino más bien escarificador y consumidor de frutos y semillas; Dasyprocta, con Dasyprocta punctata, localmente llamado Patecera, que es diurno y utiliza túneles sólo para proteger a las crías; su alimento principal son las semillas y las almacena en un patrón disperso, enterrándolas en huecos poco profundos. 3°) Tayassuidae: Tayassu tajacu (Tatabro o Saíno) y T pecarí (Puerco manao), ávidos consumidores de frutos y semillas, probablemente son más consumidores y depredadores que consumidores.
- Dispersión por Aves
Es típica en las semillas de Virola (;Mirysticace.ae;) y Protium (;Burseraceae); también consumidos y diseminados por Ramphastidae (familia de la que existen los llamados Tucanes y Pichís del catival. Las Psittacidae son principalmente consumidoras y destructoras de frutos y semillas, pero cumplen importante papel de dispersión (indirecta) en especies arbóreas de frutos medianos y pequeños, como Anacardium excelsum. Analizando el estudio de Orozco (1995) se infiere que aproximadamente. el 41% de las especies de aves del catival son potenciales diseminadoras de semillas, de modo que este grupo animal debe ser considerado debidamente para evitar regresiones graves en la recuperación de los cativales aprovechados.
Otros síndromes de dispersión en el catival y las especies asociadas son: a) Hidrocoria: el Cativo, pues la semilla posee forma y cámaras de flotación que le permiten sobreaguar y transportarse durante largas distancias hasta instalarse para proceder a la germinación en sitio favorable. b) Explosión del Fruto: la Ceiba Amarilla (Hura crepitans), el Dormilón (Pentaciethra macroloba) y el Camajón (Sterculia apetala). c) Anemocoria: especies de semilla alada, como Terminalia chiriquensis (Guayabillo), Tabebuia rosea (Roble), Cedrela sp. (Cedro), Pterocarpus officinalis y P.' rohrii (Bambudo). d) Epizoocoria: Cadillo (Sloanea sp. y Sloanea picapica) y Guácimo blanco (Trichospermun mexicanum).
Los anteriores aspectos señalan la importancia de mantener cierto grado de equilibrio en las poblaciones de la fauna silvestre en un bosque tropical bajo manejo forestal por su rol en el mantenimiento del funcionamiento y la productividad de bienes y servicios. Incluso, a la fauna se le debería considerar como un instrumento "natural" del manejo, pues le ahorra esfuerzos económicos y técnicos al silvicultor al evitarle inversiones y esfuerzos en viveros y reproducción ex situ de las especies deseables.
CONLUSIONES Y RECOMENDACIONES
BIBLIOGRAFIA
ARROYO, M.T.K. y SQUEO, F. 1990. Relationship between plant breeding systems and pollination. En: Breeding Systems and Pollination. New York, Academic Press. pp: 205-227. Tabls., gr fs.,maps.
CORPORACION NACIONAL DE INVESTIGACION Y FOMENTO FORESTAL. Archivo Técnico T-5. Project CONIFPIZANO S.A.; Investigación para el Manejo del Catival.
CORPORACION NACIONAL DE INVESTIGACION Y FOMENTO FORESTAL. 1993. Estado del conocimiento sobre la Asociación Catival en Colombia. Informe de Avance Contrato CONIF-COLCIENCIAS. Santafé de Bogotá, D.C. Documento en borrador.
CORPORACION NACIONAL DE INVESTIGACION Y FOMENTO FORESTAL: PIZANO. 1995a. Estudio comparativo: caracterización de la asociación catival en estado climácico y 21 arios después de la intervención con énfasis en su biodiversidad. La Gira y Clavellino, Bajo Atrato, Chocó, Colombia. Santafé de Bogotá, 180 p. Ilus., tabls., gráfs.
CORPORACION NACIONAL DE INVESTIGACION Y FOMENTO FORESTAL. 1995b. Análisis de los proyectos BAL-RN-06- 89, BAL-RN-07-9 y BAL-RN-08-91 que enmarcan la investigación de la dinámica y manejo de la regeneración natural en la asociación catival. Informe del contrato CONIFPIZANÓ S.A. Tabs., gráfs.
ECHAVARRIA, J. y VARON, T. 1988. Estudio dendrológico de la asociación catival en La Balsa, Chocó. Tesis Ing. For., Medellín, Colombia. Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia. 548 p.
ESCOBAR, J. y VASQUEZ F. 1987. Caracterización de tipos de cativales. Tesis Ing. For., Medellín, Colombia. Facultad de Agronomía, Universidad Nacional de Colombia. 201 p.-
FINEGAN, B. 1993. Curso Intensivo Internacional de Silvicultura y Manejo de Bosques Naturales Tropicales. 1993. I. Tema: bases ecológicas para la silvicultura. Turrialba, Costa Rica, Catie. 225 p. Ilus., tabls., gr fs.
INSTITUTO COLOMBIANO DE HIPROLOGIA, METEOROLOGIA Y ADECUACION DE TIERRAS (HIMAT). 1986. Hojas de Registros diarios .de la Estación climatológica Grado II "La Balsa", Riosucio — Chocó. Archivo Técnico de la Estación Silvicultura' "La Balsa".
LINARES, RICARDO. 1988. Estudio preliminar de la asociación catival en Colombia. Bogotá, Conif y Pizano S.A. 68 p. (Conif, Serie de Documentación No.17).
LINARES, RICARDO. 1995. Bases ecológicas para la silvicultura y el aprovechamiento de los bosques naturales en Colombia. En: Talleres regionales sobre aprovechamiento de los bosques naturales; Florencia (Caquetá), Leticia (Amazonas), Bucaramanga (Santander) y Cali (Valle), 1995. Proyecto Concesiones y Permisos Forestales PPD 11/93 Organizacion internacional de Maderas Tropicales - Ministerio del Medio Ambiente República de Colombia.
LINARES, RICARDO. 1998. Conservación, utilización y manejo sostenible del Catival (Prioretum copatferae). Resultados de 16 arios de investigaciones. Convenio Conif - Pizano S.A. Santafé de Bogotá, Conif. Documento en edición para publicación. 118 p.
LINARES, RICARDO Y MARTINEZ HIGUERA, HUGO. 1991. La regeneración natural temprana del bosque de cativo en el Chocó, Colombia. Bogotá, Conif. Serie Técnica No. 30.
LINARES, RICARDO et al: 1997. Bases ecológicas para la silvicultura del catival (Prioretum copaiferae). Informe Preliminar del Proyecto sobre Publicación del Catival. Documento de avance para discusión presentado por la Corporación Nacional de Investigación y Fomento Forestal — Conif. Santafé de Bogotá, Conif, Contrato Pizano S.A. — Conif 48 p.
MONTERO, M. y CORDOBA G. 1996. Estudio de la dinámica poblacional de Prioria copaifera (Grisebach) en un bosque de la llanura aluvial del río Atrato. Santafé de Bogotá , Universidad Nacional de Colombia. 150 p. Ilus., tabls., gráfs., maps. Tesis (Biól.)
OROZCO, R.H. 1995a. Contribución al estudio de los efectos del aprovechamiento forestal sobre la comunidad de aves de la consociación Priorietum copaiferae (Catival), en el Chocó colombiano. Santafé de Bogotá, D.C., Pontificia Universidad Javeriana. Facultad de Ciencias. Departamento de Biología. Ilus., tabls., gráfs., maps. Tesis (Biól.).
OROZCO, R. H. 1995b. Visión general de la Avifauna asociada a los Cativales del área de aprovechamiento forestal "La Balsa" (Riosucio-Chocó). Pizano S.A. Dirección de Recursos Naturales. 38p.
RIOS, SANDRA. 1996. Estudio de la arquitectura de la comunidad Prioria copaifera Grisebach, (Caesalpinaceae), en un bosque inundable de la región del bajo Atrato, Chocó, Colombia.
RODRIGUEZ, J. VICENTE., HERNANDEZ, J. Y DÁVILA, A.. 1995. Manejo de Vida Silvestre. Pizano S.A. Región de Domingodó, Chocó. 100p.
ROLLET, B. 1974. L'architecture des forest denses humides sempervirentes de plaine. Nogent sur Mame, Francia. Centre Technique Forestier Tropical. 298 p.
URDANETA, M. F. 1995. Diferencias en la composición y estructura de la comunidad de aves en época seca de tres agroecosistemas y un bosque primario asociados a un fragmento de catival maduro en el Chocó Colombiano. Pontificia Universidad Javeriana. Facultad de Ciencias, Departamento de Biología. Santafé de Bogotá. 173p.
WHITMORE, T. 1984. Tropical rain forest of de Far East. Oxford, Reino Unido: Clarendon Press. 314 p.
SCHULZ, J.P. 1960. Ecological studies of rain forest in Northern Suriname. Amsterdam, - NoordHollandsche Vitgevers Maatschaooyj. 267 p.
Creation date:




License
Colombia Forestal retains the patrimonial rights (copyright) of the published works, and favors and allows the reuse of the same under the Creative Commons Attribution-ShareAlike 4.0 International license, so they can be copied, used, disseminated, transmitted and exhibited publicly, provided that:
You acknowledge the credits of the work in the manner specified by the author or licensor (but not in a way that suggests that you have their support or that they endorse your use of their work).