
DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2006.1.a06Publicado:
01-01-2006Número:
Vol. 9 Núm. 19 (2006)Sección:
Artículos de investigación científica y tecnológicaAspectos de la estructura y del patrón de riqueza de la vegetación del transecto del Tatamá
Aspects of the structure and richness pattern of the Tatama transect vegetation
Palabras clave:
Estructura, vegetación, riqueza, Tatamá, cordillera occidental, Colombia (es).Descargas
Referencias
Arellano, II. 2001. Estudio de la vegetación del sistema andino de la serranía del Perijá. Tesis de pregrado. Departamento de Biología. Universidad Nacional de Colombia. Bogotá. 160 p.
Cantillo, E.E. 2001. Caracterización estructural, ordenación y dinámica de la vegetación en la zona de captación de aguas de la microcuenca El Tigre, municipio de Norcasia, Caldas. Tesis de Maestría. Universidad Nacional de Colombia. 178 p.
Cantillo, E.E., K. Rodríguez y A. Avella. 2005. La vegetación leñosa de la Reserva Forestal Cárpatos, Guasca, Cundinamarca. Centro de Investigaciones y Desarrollo Científico. Universidad Distrital Francisco José de Caldas. Fondo de Publicaciones. 109 pp. Bogotá.
Cantillo, E.E. y J.O. Rangel-Ch. (En imprenta). Aspectos de la estructura y del patrón de riqueza de la vegetación del transecto Sumapaz. En: T. van der Hammen & A. Cleef (eds). La Cordillera oriental (Colombia). Transecto Sumapaz. Estudios de Ecosistemás Tropandinos-Ecoandes.
Cantillo, E.E., J.O. Rangel-Ch & II. Arellano. (En imprenta). Aspectos de la estructura y del patrón de riqueza de la vegetación de la Serranía del Perijá. Instituto de Ciencias Naturales. Universidad Nacional de Colombia. Bogotá.
Duivenvoorden, J.F. & J.M. Lips. 1993. Ecología del paisaje del medio Caquetá. Estudios de la Amazonia Colombiana. Tropenbos. Colombia. Bogotá. 301 p.
Florez, A. 1987. Geomorfología estructural y dinámica morfogenética del transecto Sumapaz (Cordillera oriental). Análisis Geográficos 12. Bogotá. 50 p.
Flórez, A. 2005. Aspectos geomorfológicos del área del transecto Tatamá. En: T. van der Hammen, J.O. Rangel-Ch & A. Cleef (eds.). La cordillera occidental colombiana. Transecto Tatamá. Estudios de Ecosistemas Tropandinos-Ecoandes 6; 797-812. J. Cramer, Berlín-Stuttgart.
Lincoln R.J., G.A. Boxshall & P.F. Clark. 1986. Diccionario de ecología, evolución y taxonomía. Fondo de Cultura Económica. México.
Mori, 5. & B. Boom. 1987. The Lecythidaceae of the lowland neotropical forest: La Fumeé Montain, French Guiana. Mem. New York Bot, Garden 44: 9-29.
Moreno, C.E. 2001. Métodos para medir la biodiversidad. Manuales y Tesis SEA. Vol 1. CYTED, ORCYT/UNESCO & SEA. Zaragoza. 84 p.
Patirio, R.D. 2003. Estudio de la flora y la vegetación del bosque en la Estación de Primates Colosó, Sucre. Tesis de Maestría. Instituto de Ciencias Naturales. Universidad Nacional de Colombia. Bogotá. 114 pp.
Rangel-Ch, J.O. y G. Lozano-C. 1986. Un perfil de vegetación entre La Plata (Huila) y el Volcán Puracé. Caldasia 14 (68-70): 533-547.
Rangel-Ch., J.0., J. Aguirre-C & R. Sánchez-M. 1990. Aspectos estructurales, dinámicos y fisionómicos de la vegetación. En: J. Aguirre & J.O. Rangel-CH. (eds). Biota y ecosistemas de Gorgona. Fondo FEN Colombia. Bogotá. Pp. 127-151.
Rangel-Ch, J.0.. 1991. Vegetación y ambiente en tres gradientes montañosos de Colombia. Tesis de Doctor, Universidad de Ámsterdam. Amsterdam. 349 pp.
Rangel-Ch., J.O. y A. Garzón. 1994. Aspectos de la estructura, de la diversidad y de la dinámica de la vegetación del Parque Regional Natural Ucumari. En: J.O. Rangel-Ch. (ed.). Ucumarí: Un caso típico de la diversidad biótica andina: 59:84. Publicaciones de la CARDER. Pereira.
Rangel-Ch, J.0., y A. Velásquez. 1997. Métodos de estudio de la vegetación. En J.O. Rangel-Ch, P. Lowy y M. Aguilar. Colombia Diversidad Biótica II. P: 59-87. Instituto de Ciencias Naturales. Universidad Nacional de Colombia. Bogotá.
Rangel-Ch., J.0., A.M.Cleef & S. Salamanca. 2003. The ecuatorial interandean and subandean forest of the Parque Los Nevados Transect, Cordillera Central, Colombia. En: T. van der Hammen & A. Dos Santos (eds). La Cordillera Central Colombiana. Estudios de Ecosistemas Tropandinos 5: 143-204. J. Cramer, Berlín-Stuttgart.
Rangel-Ch, J.0.. 2005. Aspectos de la estructura y de la riqueza de especies en la vegetación de los transectos del Tatamá. En: T. Van der Hammen, J. O. Rangel-Ch & A. Cleef (eds). La cordillera occidental colombiana. Transecto Tatamá. Estudios de Ecosistemás Tropandinos-Ecoandes 6; 797-812. J. Cramer, Berlín-Stuttgart.
Rangel-Ch, J.0., A. M. Cleef, S. Salamanca y C. Ariza. 2005a. La vegetación de los bosques y selvas del Tatámá. En: T. Van der Hammen, J.O. Rangel-Ch & A. Cleef (eds). La cordillera occidental Colombiana. Transecto Tatamá. Estudios de Ecosistemás Tropandinos-Ecoandes 6; 469-643. J. Cramer, Berlín-Stuttgart.
Rangel-Ch, J.0., M. Aguilar-P. y A. Flórez. 2005b. Clima en el área del transecto Tatamá. En: T. Van der Hammen, J.O. Rangel & A. Cleef (eds). La cordillera occidental colombiana. Transecto Tatamá. Estudios de Ecosistemás Tropandinos-Ecoandes 6; 47-103. J. Cramer, Berlín-Stuttgart.
Ruiz-B, E. 2005. Climosecuencia altitudinal de los suelos del transecto Tatamá. En: T. Van der Hammen, J.O. Rangel & A. Cleef (eds). La cordillera occidental colombiana. Transecto Tatamá. Estudios de Ecosistemás Tropandinos-Ecoandes 6; 155-223. J. Cramer, Berlín-Stuttgart.
Sturges H.G. 1926. The Choice of class interval. Joumal of the American Statistical Association. 21:65-66
UNESCO/PNUMA/FAO. 1980. Ecosistemas de los Bosques Tropicales. UNESCO/CIFCA. Madrid. Pp 126-162.
Van der Hammen, T. 2005. Introduction: The study of the Tatamá transect. En: T. van der Hammen, J. O. Rangel-Ch & A. Cleef (eds). La cordillera occidental colombiana. Transecto Tatamá. Estudios de Ecosistemas Tropandinos-Ecoandes 6; 797-812. J. Cramer, Berlín-Stuttgart.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Artículos de investigación científicos y tecnológicos
Colombia Forestal, 2006-12-00 vol:9 nro:19 pág:88-125
Aspectos de la estructura y del patrón de riqueza de la vegetación del transecto del Tatamá
Édgar Ernesto Cantillo H.
ecantillo@udistrital.edu.co
Universidad Distrital Francisco José de Caldas.
J.Orlando Rangel Ch.
jorangelc@unal.edu.co
Universidad Nacional de Colombia
Resumen
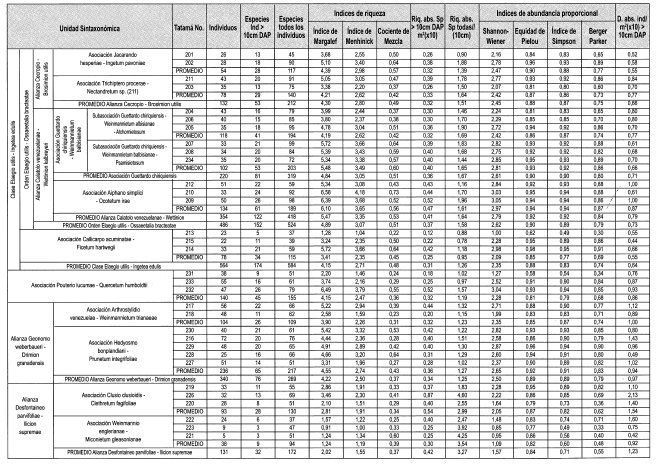
A lo largo de dos transectos con variación altitudinal entre 550 y 3.700 m en la vertiente occidental y entre 1.150 y 3.700 m en la oriental de la cordillera occidental, se caracterizaron los aspectos de la estructura de la vegetación boscosa. En la vertiente occidental de la región de vida Tropical, la más húmeda de la cordillera, la vegetación definida por la alianza Cecropio-Brosimion utilis presenta en el estrato arbóreo superior individuos emergentes mayores a 30 m, mayor dominancia de los estratos arbóreo inferior (64%), arbustivo (55%) y herbáceo (53%) y mayor altura promedio del dosel (23 m). Las especies con valores mayores de importancia son Elaeagia utilis, Jacaranda hesperia, Inga pavonia y Cecropia sp., mientras que las familias más representativas fueron Rubiaceae, Moraceae y Leguminosae. En la región de vida Subandina, la alianza Calatolo venezuelanae-Wettinion kalbreyeri, contiene mayor dominancia en el estrato arbóreo superior (23%). Las especies más importantes son Alchornea sp., Cecropia sp. y Wettinia kalbreyeri, con familias representativas como Lauraceae, Rubiaceae y Moraceae. Contiene el mayor número de especies (418) géneros (229) y familias (81) en el transecto. La región de vida andina está representada por la alianza Geonomo weberbaueri-Drimion granadensis, la cual registra mayor dominancia en el estrato subarbóreo (52%). Contiene especies importantes como Clusia multiflora, Prunus integrifolia y Brunellia sp., destacándose las familias Melastomataceae, Clusiaceae y Cunoniaceae. En la franja altoandina, la alianza Desfontaineo parvifoliae-Ilicion supremae presenta mayor promedio en el número de individuos (97) y en el área basal promedio (9,6 m2) en elementos con DAP > 10 cm en 0,1 ha, y la más baja altura promedio del dosel en el transecto (10 m), se destacan Clusia clusioides, Miconia gleasoniana, Scheffiera bejucosa e Ilex aif pernervata, como especies importantes, y Melastomataceae, Aquifoliaceae y Araliaceae como familias representativas.
A escala regional, los valores de densidad absoluta en individuos con DAP > 10 cm fueron mayores en la región de vida altoandina en la vertiente occidental (1,18) y andina en la oriental (1,13). En las dos vertientes, la riqueza absoluta para individuos con DAP > 10 cm fue mayor en la región de vida subandina (0,46 y 0,35) mientras que para todos los éstrátos lo fue en la región de vida altoandina (3,19 y 3,40).
Palabras claves:
Estructura, vegetación, riqueza, Tatamá, cordillera occidental, Colombia
Abstract
Along two transects with variation altitudinal between 550 and 3.700 m on the West flank and 1.150 and 3.700 m on the East flank of the Colombian Western Cordillera, the aspects of the structure of the forest vegetation were characterized. In the West flank of the Tropi cal life-region, the most humid in the cordi llera, the vegetation defined by the alliance Cecropio-Brosimion utilis, presents in the uppermost arboreal superior stratum greater emergent individuals to 40 m, bigger dominancia of the lower arboreal stratum (64%), shrubs (55%) and herbaceous (53%) and bigger height average of the canopy (23 m). The species with bigger values of importance are Elaeagia utilis, Jacaranda hesperia, Inga pavonia and Cecropia sp., while the most representative families were Rubiaceae, Moraceae and Leguminosae. In subandean life-region, the alliance Calatolo venezuelanae-Wettinion kalbreyeri, contains bigger dominancia in the lower arboreal stratum (23%). The most important species are Akhornea sp., Cecropia sp. and Wettinia kalbreyeri, with representative families as Lauraceae, Rubiaceae and Moraceae. It contains the biggest number of species (418) genera (229) and families (81) in the arboreal strata. The Andean life-region is represented by the alliance Geonomo weberbaueri-Drimion granadensis, which registers bigger dominancia in the stratum sub-arboreal (52%). It contains important species as Clusia multiflora, Prunus integrifolia and Brunellia sp., standing out the families Melastomataceae, Clusiaceae and Cunoniaceae. In upper-andean life-region,the alliance Desfontaineo parvifoliae-Ilicion supremae presents bigger average in the number of individuals (97) and in the area basal average (9,6 m2) in elements with DAP>10 cm in 0.1 ha, and the lowest height average in the canopy in the transect (10 m); Clusia clusioides, Miconia gleasoniana, Scheffiera bejucosa and Ilex aff. Pernervata, as important species, and Melastomataceae, Aquifoliaceae and Araliaceae like representative families.
On regional scale, the values of absolute density in individuals with DAP>10 cm was greater in the region of upper-andean life-region in the West flank (1,18) and Andean in the East flank one (1,13). In the two flanks, the absolute riches for individuals with DAP>10 cm was bigger in the sub-andean life-region (0.46 and 0,35) whereas for all the strata, it was it in the region of upper-andean life-region (3,19 and 3,40). The values of riches of species in the sub-arboreal stratum of the sub-andean life-region of the West flank (29), they are the highest registered in the Andean region of Colombia.
Key words:
Structure, vegetation, richness, Tatamá, cordillera occidental, Colombia
INTRODUCCIÓN
La caracterización de la vegetación puede realizarse sobre la base de definir espectros florísticos derivados de inventarios de campo detallados (Rangel, 2005) o mediante metodologías que combinen atributos diferenciables de manera rápida y fácil. Estas clasificaciones toman como base las pautas fisionómicas, es decir, el aspecto de la estructura de las plantas (Rangel et al., 1990; Rangel, 1991; Rangel y Garzón, 1994; Cantillo, 2001; Arellano, 2001; Patirio, 2003; Cantillo et al., 2005; Cantillo y Rangel, en imprenta; Cantillo et al., en imprenta). En esta contribución se presentan los aspectos que definen las características estructurales de la vegetación boscosa en los transectos del Tatamá, macizo que forma parte de la cadena montañosa de la cordillera occidental. La caracterización florística de sus comunidades vegetales fue realizada por Rangel et al. (2005a). Rangel (2005) analizó la variación de parámetros estructurales como cobertura y densidad. En este estudio, se complementa esta aproximación con análisis detallados de altura, diámetro a la altura del pecho (DAP), área basal, densidad e índices ecológicos para especies y familias (IVI, IPF, IVIF), relacionándolos con las condiciones climáticas definidas por Rangel et al. (2005b) y las características edáficas más sobresalientes que prevalecen en la región (Ruiz, 2005).
ÁREA DE ESTUDIO
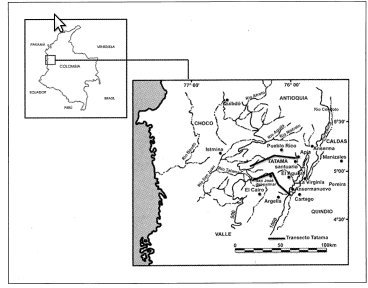
Los transectos efectuados en el macizo del Tatamá se localizan entre las latitudes norte de 4°43' a 5°14' y longitudes Oeste de 75° a 76°27' (Figura 1), entre 500 y 4.100 m en la vertiente occidental y entre 1.100 y 3.900 m en la vertiente oriental, comprendiendo territorio de los departamentos de Chocó, Risaralda y Valle del Cauca (Rangel, 2005).
Según Flórez (1987; 2005), la región del Tatamá se localiza entre las depresiones tectónicas del Cauca, al Este, y del Atrato-San Juan, al Oeste. Entre las unidades geológicas que caracterizan esta región, las del grupo Dagua y el grupo Diabásico se encuentran profundamente alteradas, falladas y diaclasadas, y solamente los gruesos bancos de areniscas y conglomerados del macizo de Tatamá presentan una menor alteración. El relieve muestra pendientes abruptas (mayores de 20°) y bloques levantados, además de los plutones que produjeron relieves altos y escarpados. El clima húmedo actual, el bosque de escaso anclaje vertical, los suelos impermeables en la mayor parte de la vertiente occidental y en partes altas de la vertiente oriental, con horizontes argílicos endurecidos, hacen que la mayor parte de esta área sea potencialmente inestable. Los suelos están representados por el 85% de Inceptisoles, 6% de Histosoles y Molisoles y 3% de Entisoles. La mayor parte de los perfiles estudiados son suelos poco evolucionados, perfil AC y ABC, ricos en materia orgánica, colores oscuros en los horizontes superiores y friables, químicamente muy ácidos, de complejo de cambio muy desaturado.
Al ser el clima más húmedo en la vertiente occidental y en las partes altas de la vertiente oriental, no se presenta por tanto un régimen ústico, como si es común en altitudes menores de 1.200 msnm de la vertiente oriental (Ruiz, 2005). Estas condiciones de alta humedad en los suelos han favorecido la conservación de los minerales amorfos y, por tanto, la domi- . nancia de Andosoles en los suelos del transecto. En la vertiente occidental, las secuencias observadas en el transecto en orden ascendente de altitud fueron: hasta 1.000 m, Typic Distropept-Andic Humitropept; entre 1.000 y 3.500 m los subgrupos Entic Dystrondept, Hydric Dystrondept y Typic Dystrondept; en alturas mayores a los 3.500 m, donde las áreas son de menor pendiente, se encuentran Typic Cryandept, Terric y Lithic Cryosaprist; En la vertiente oriental, entre 1.100 y 1.275 m, se encuentran los subgrupos Entic Haplustoll Udorthentic Haplustoll; a 1.600 m, Lithic Humitropept; entre 1.830 y 3.300 m, al igual que en la vertiente occidental, dominan los subgrupos Typic Dystrondept, Entic Dystrondept e Hydric Dystrondept; a alturas mayores a 3.300 m, se encuentran Hydric Cryandept y Typic Cryandept (IGAC 1987; Ruiz 2005). El clima, ampliamente descrito en Rangel et al. (2005), muestra que en la vertiente occidental, los valores de precipitación y humedad relativa son mayores que los de la vertiente oriental donde, por el contrario, las expresiones de evaporación, brillo solar y temperatura son más altas. En la vertiente occidental, los análisis de precipitación promedio por regiones de vida son: en la tropical, varían entre 4.267,1 mm en La Unión y 6.146,5 mm en Santa Cecilia, con promedio de 5003,2 mm, en la subandina oscila entre 2.192,5 mm en Pueblo Rico y 3.732,6 mm en San José del Palmar, con promedio de 2962,55 mm, en la andina, el monto anual es de 5.395,3 mm en El Sinaí. En la vertiente oriental, la región de vida subandina tiene una variación entre 1.444,3 mm en Santiago Gutiérrez y 2.913,3 mm en Campoalegre, y en la andina, el promedio anual es de 2.180,7 mm en El Porvenir. Predominan los regimenes de distribución bimodal-tetraestacional desde las colinas bajas del Andén Pacífico hasta la región subandina.
METODOLOGÍA
Para el análisis de la cobertura, número de individuos y número de especies, se tomaron los datos referenciados en Rangel (2005).
La estratificación de la cobertura se delimitó a partir de las categorías propuestas por Rangel y Lozano (1986). La distribución de la cobertura relativa fue estimada para todos los estratos. La estimación de la altura media del dosel superior se adaptó de la propuesta de Duivenvoorden y Lips (1993), a partir del promedio de alturas de los árboles más altos que alcanzan una cobertura total de 60%.
La estructura horizontal se determinó para los estratos arbóreos a partir de los valores de abundancia, frecuencia, área basal y cobertura con el fin de estimar el índice de valor de importancia de Cottam, IVI, y el índice de predominio fisionómico, IPF (Rangel y Velásquez, 1997). Para determinar la distribución de las variables altura (talla), diámetro y cobertura, se siguió el modelo de Sturges (1926). El patrón de distribución de diámetro y altura se definió para los estratos arbóreos. En la distribución de diámetros de las especies de bosques tropicales se utilizan intervalos de 10 cm por clase diamétrica, puesto que el número de árboles desciende casi geométricamente al aumentar las clases, esto es, entre clase y clase se reduce a la mitad; además, deben excluirse en áreas grandes, diámetros menores de 1,Q,cm de DAP ya que el número de especies es elevado (UNESCO, 1980).
Con base en los resultados de la distribución de altura, se incluye un modelo de estratificación adaptado de Lincoln et al. (1986), que considera conceptos como dominante o emergente: uno o varios individuos que ejercen una influencia considerable sobre una comunidad debido a su altura y cobertura; codominante: grupo de individuos con menor altura y cobertura que los dominantes, pero que ejercen gran influencia sobre los estratos más bajos en una comunidad vegetal (semidominancia); dominado: individuos inhibidos o reprimidos por los estratos superiores; suprimido: generalmente el estrato con mayor número de individuos leñosos de tallas bajas o en estados iniciales de desarrollo.
Se determinaron los índices de riqueza (Margalef, Menhinick), de heterogeneidad (Simpson, Berger-Parker) y de equidad (Shannon-Wiener) para cada grupo florístico (Moreno, 2001). La riqueza absoluta para cada grupo se definió como el promedio de la relación (No. de especies/área de muestreo) x10, de cada levantamiento; además, se definió el Índice de Valor de Importancia para Familias (Mori y Boom, 1987) y las familias y especies dominantes por estrato para cada comunidad.
RESULTADOS
VARIACIÓN GLOBAL DE LA VEGETACIÓN
Como complemento a la contribución de Rangel (2005), en la cual se analiza la variación de parámetros estructurales como cobertura, densidad y riqueza, se presenta a continuación la descripción de índices y otros parámetros como altura, diámetro a la altura del pecho (DAP), área basal, riqueza absoluta y los patrones de distribución.
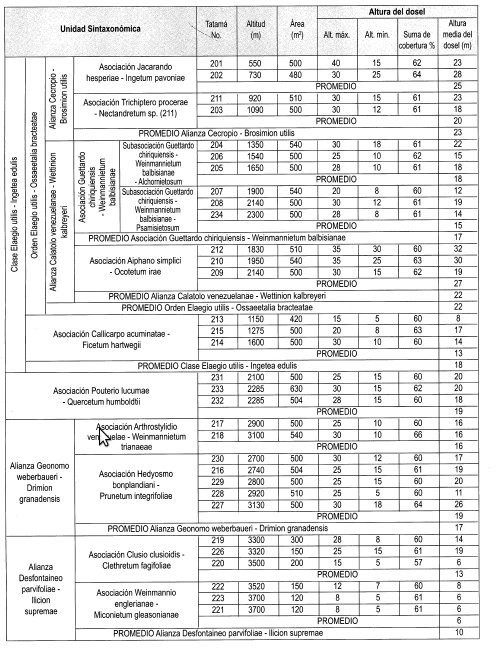
Altura del dosel (Tabla 1, Figura 4)
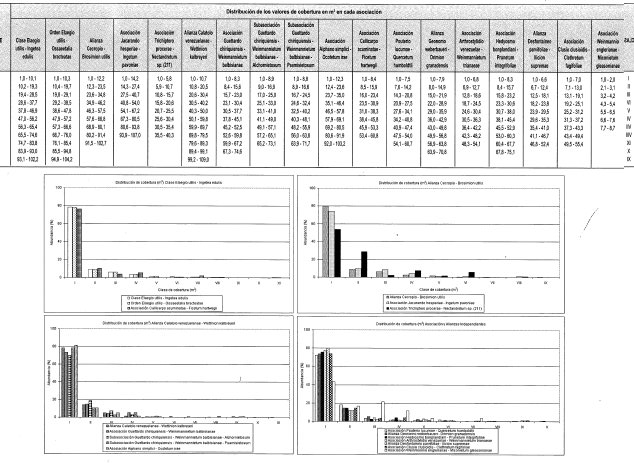
Vertiente oriental. La altura promedio del dosel varía entre 6 m (TAT 220, 221) a 3.500 m y 3.700 m respectivamente, y 32 m (TAT 212) a 1.830 m.
Vertiente occidental. Con valores superiores más bajos, alcanzan entre 6 m (TAT 223) a 3.700 m y 28 m (TAT 202) a 730 m.
Densidad absoluta individuos con DAP >10cm (Tabla 5)
Vertiente oriental. Varía entre 0,42 (TAT 221) a 3.700 m y 2,13 (TAT 226) a 3.320 m.
Vertiente occidental. La fluctuación es menos extrema que en el flanco oriental al variar entre 0,49 (TAT 228) a 2.920 m y 1,6 (TAT 222) a 3.520 m.
Área basal individuos con DAP > 10 cm (Tabla 2)
Vertiente oriental. El valor calculado varía entre 0,6 m2 (TAT 221) a 3.700 m y 35,3 m 2 (Lev 226) a 3.320 m.
Vertiente occidental. La variación es más estrecha 1,8 m2 (TAT 203) a 1.090 m y 9,6 m2 (Lev 227) a 3.130 m.
Riqueza absoluta (Tabla 5)
Vertiente oriental. En todos los estratos varía entre 0,78 (TAT 215) a 1.275 m y 4,6 (TAT 226) a 3.320 m. En individuos con DAP > 10 cm, la variación es de 0,12 (TAT 213) a 1.150 m y de 0,87 (TAT 226) a 3.320 m.
Vertiente occidental. Para todo el conjunto fluctúan los valores entre 0,9 (TAT 201) a 550 m y 3,92 (TAT 223) a 3.700 m. Individuo s con DAP > 10 cm, varían entre 0,25 (TAT 223) a 3.700 m y 0,42 (TAT 230) a 2.700 m.
DISTRIBUCIÓN DE LA VEGETACIÓN SEGÚN REGIONES DE VIDA Y UNIDADES SINTAXONÓMICAS
REGIÓN DE VIDA TROPICAL
La Alianza Cecropio-Brosimion utilis representa esta región de vida en la vertiente occidental. La vegetación se establece entre 550 m y 1.090 m.
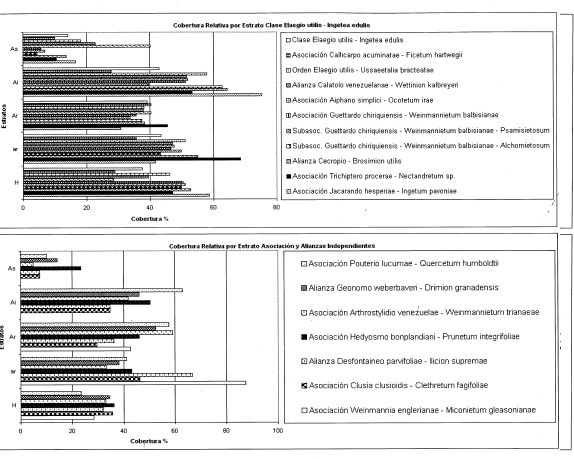
Cobertura (Figura 2)
Arbóreo superior.
El valor máximo fue 24% (TAT 202) a 730 m. El valor promedio fue 14%.
Arbóreo inferior.
Varió entre 51% (TAT 203) a 1.090 m y 73% (TAT 201) a 550 m. El valor promedio fue 64%.
Arbolitos o subarbóreo.
Fluctuó entre 26% (TAT 202) a 730 m y 48°/.9 (TAT 203) a 1.090 m. El promedio de cobertura fue 38%.
Arbustivo.
Los valores extremos fueron 24% (TAT 201) a 550 m y 76% (TAT 203) a 1.090 m, con promedio para el estrato de 55%.
Herbáceo.
Varía entre 40% (TAT 211) a 920 m y 61% (TAT 201) a 550 m. El promedio fue 53%.
El diagrama estructural (Figura 2) muestra el estrato Arbóreo inferior como dominante al cubrir el 64%; los estratos Arbustivo y Herbáceo superan el 53% de la cobertura.
Distribución de la cobertura (m2)(Figura 3)
Los valores de cobertura se distribuyen en nueve clases, cuyos límites oscilan entre 1,0 y 102,7 m2. La clase I (1,0-12,2 m2) contiene al 79% de los individuos y la clase 11 (12,3-23,5 m2) el 9%.
Altura del dosel (Tabla 1, Figura 4)
La altura promedio del dosel es 23 m, con variación entre 18 m (TAT 203) a 1.090 m y 28 m (TAT 202) a 730 m.
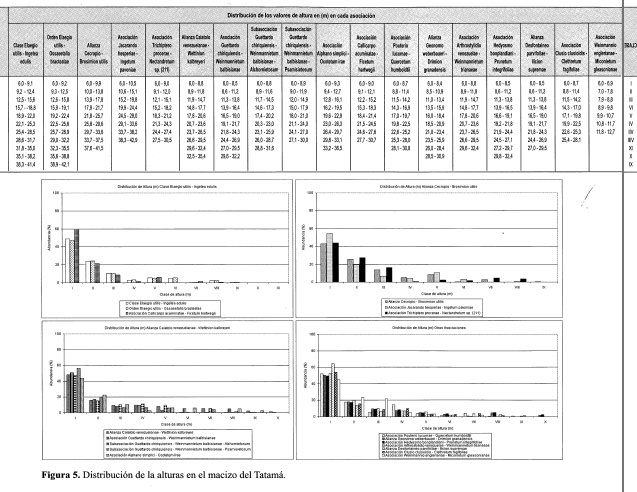
Distribución de las alturas (tallas) (Figura 5)
Se distribuyen en nueve clases con valores entre 6,0 y 40 m. La clase I (6,0-12,1 m) agrupa al 68% de los individuos y la clase II (12,218,3 m) el 16%; juntas reúnen el 84% de los individuos totales. Al agrupar clases se diferencian cuatro estratos: el suprimido, menor a 12 m, contiene el 60% de los individuos, el dominado entre 12 y 25 m que retiene el 30%, el codominante entre 25 y 37 m con el 8% y el dominante entre 37 y 40 m con el 2%.
Número de individuos estrato arbóreo
Se registraron en este grupo un total de 228 individuos mayores de 5 m en un área de 1.990 m2, de los cuales 132 tienen DAP > 10 cm.
Arbóreo superior
Se presenta un promedio de tres individuos por levantamiento, variando entre uno (TAT 203) a 1.090 m y cuatro (TAT 211) a 920 m.
Arbóreo inferior
Varía entre once individuos (TAT 201) a 550 m, y 18 (TAT 211) a 920 m. El promedio es de 16 individuos por levantamiento.
Arbolitos o subarbóreo
El número de individuos fluctúa entre 28 (TAT 202) a 730 m y 49 (TAT 211) a 920 m, con un promedio de 39 individuos por levantaliento.
Densidad absoluta individuos con DAP 210 cm (Tabla 5)
La variación es de 0,52 (TAT 201) a 550 m y 0,84 (TAT 211) a 920 m.
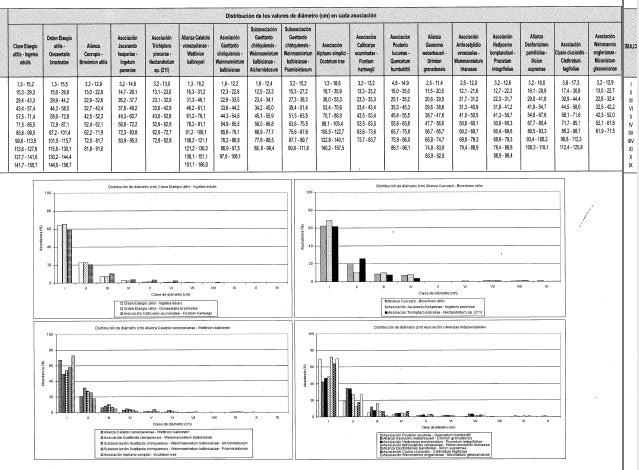
Distribución del DAP (Figura 6)
El DAP se distribuye en nueve clases con valores entre 3,2 y 91,6 cm. La clase I (3,2-12,9 cm) agrupa al 62% de los individuos y la clase II (13,0-22,8 cm) el 20%; juntas reúnen el 82% de los individuos totales.
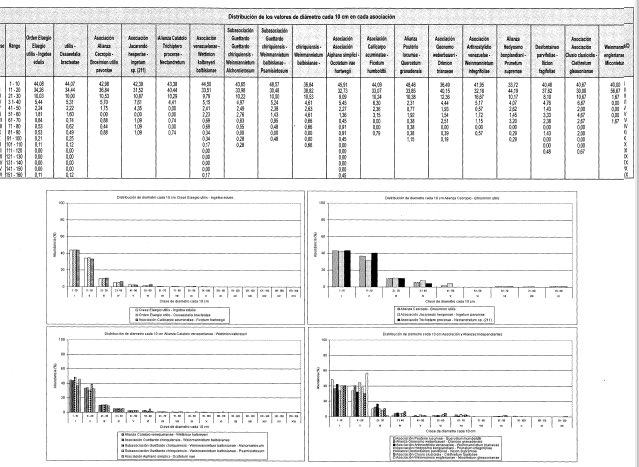
Distribución del DAP cada 10 cm en los estratos arbóreos (DAP > 10 cm) (Figura 7)
Los individuos se distribuyen en nueve clases con valores entre 1 y 90 cm. La clase I (1-10 cm) agrupa al 43% de los individuos y la clase II (11-20 cm) el 37%; juntas reúnen el 80% de los individuos totales.
Al comparar los dos tipos de distribución, se observa igualdad en el número de clases.
Área basal (Tabla 2)
Contiene un total de 7,3 m2 de área basal total, que varía entre 1 m2 (TAT 203) a 1.090 y 2,7 m2 (TAT 202) a 730. Se estima un promedio de 3,7 m2 por 0,1 ha.
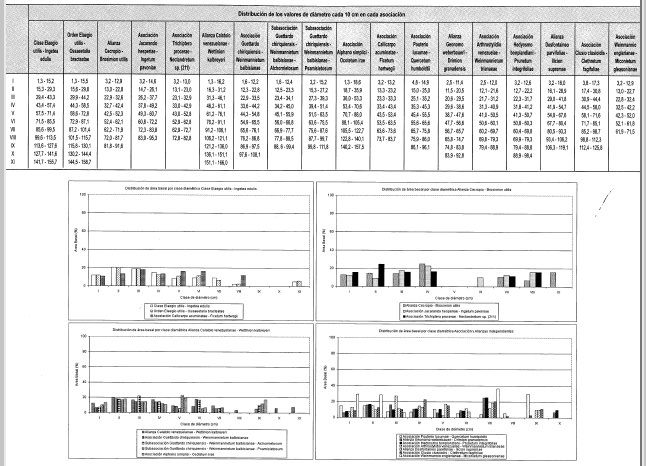
Distribución del área basal por clase diamétrica (Figura 8)
La distribución de 7,3 m2 de área basal se da en nueve clases con valores entre 3,2 y 91,6 cm. La clase IV (32,7-42,4 cm) es la de mayor frecuencia, agrupando al 25% de esta, seguida de la clase IX (81,8-91,6 cm) con el 16%. Las dos reúnen el 41% del área basal total.
Índices de valor de importancia IVI (Tabla 3) y de predominio fisionómico IPF (Tabla 4)
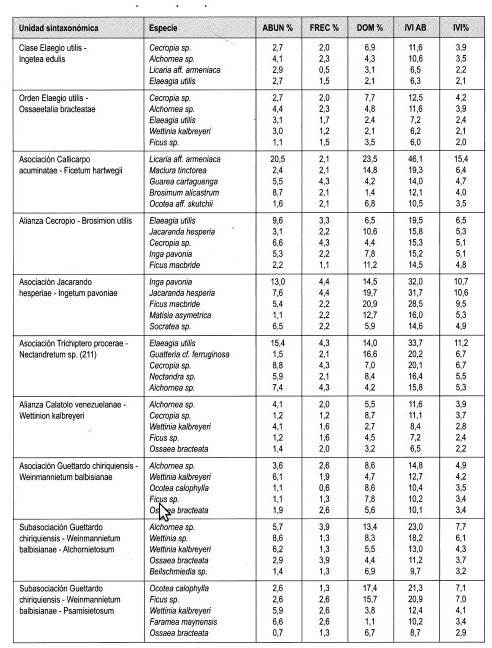
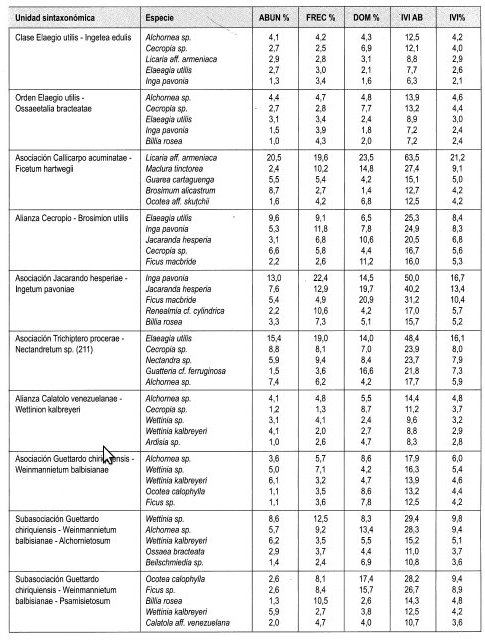
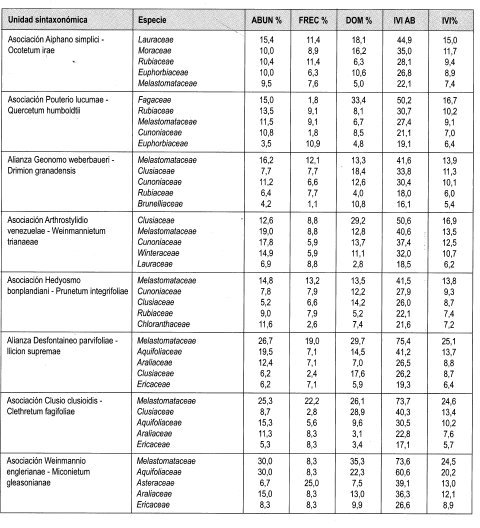
Elaeagia utilis con 6.5% de IVI es la especie con mejor expresión y es además la más abundante con 9.6%; le sigue en importancia Jacaranda hesperia (5.3%) y Cecropia sp. (5.1%). El mayor IPF lo obtuvo Elaeagia utilis con 8.4%. Otras especies importantes fueron Inga pavonia (8.3%) y Jacaranda hesperia (6.8%).
Riqueza
Número de especies
El número total de especies en todos los estratos fue 212. El número de especies por levantamiento varía entre 45 (TAT 201) a 550 m y 91 (TAT 211) a 920 m. Según estratos, se presenta la siguiente variación:
Arbóreo superior
Se encontraron ocho especies, variando entre una (TAT 203) a 1090 m y tres (TAT 211) a 920 m.
Arbóreo inferior
Se registraron 27 especies. Fluctúa entre cuatro (TAT 201) a 550 m y 11 especies (TAT 202, 211) a 730 y 920.
Subarbóreo o arbolitos
Se identificaron 59 especies en este estrato. Varía entre 21 (TAT 211) a 920 y 16 (TAT 203) a 1.090m.
Arbustivo
Se encontraron 108 especies. Los valores extremos se presentaron en el TAT 201 a 550 m con 19 y el TAT 211 a 920 m con 43 especies.
Herbáceo
El total fue 101, con fluctuación entre 19 (TAT 201) a 550 m y 39 (TAT 203) a 1.090 m.
Riqueza absoluta (Tabla 5)
En todos los estratos arbóreos varía entre 0,9 (TAT 201) a 550 m y 1,88 (TAT 202) a 73Wm., En elementos con DAP > 10 cm, la variación es de 0,26 (TAT 201, 203) a 550 m y 1.090 m respectivamente, y 0,39 (TAT 211) a 920 m.
Indice de valor de importancia para familias (Tabla 6)
El 11,1% de IVIF lo obtuvo Rubiaceae al ser más abundante; le siguen en importancia Moraceae y Leguminosae con 10,8% cada una.
En la vegetación de la alianza Cecropio-Brosimion utilis, se presentan algunas particularidades en las asociaciones que la conforman, así:
Asociación Jacarando hesperiae-Ingetum pavoniae
Se distribuye entre 550 m y 730 m, en la franja media de la región de vida Tropical. Registra cobertura muy dominante en el estrato Arbóreo inferior (75%) y presenta la mayor altura promedio del dosel (27 m). Los mayores valores de IVI e IPF lo obtienen Inga pavonia, Jacaranda hesperia y Ficus macbride. Registró el promedio mayor de área basal por 0,1 ha en la región de vida (3,9 m2).
Asociación Trichiptero procerae-Nectandretum (211)
Se distribuye entre 920 y 1.090 m, en la franja alta de la región de vida Tropical. El mayor valor de cobertura se registró en el estrato Arbustivo (69%), con alta dominancia (53%) en el Arbóreo inferior. La altura promedio del dosel es 20 m. Los mayores índices IVI e IPF lo obtienen Elaeagia utilis, Guatteria cf. ferruginosa, Cecropia sp. y Nectandra sp. Contiene mayor número de individuos con DAP > 10 cm en 0,1 ha (77) y mayor número de especies (140) géneros (108) y familias (51).
REGIÓN DE VIDA SUB ANDINA
La vegetación de la alianza Calatolo venezuelanae-Wettinion kalbreyeri; con área de distribución entre 1.350 m y 2.300 m, representa esta región de vida. Sus características principales son:
Arbóreo inferior
Varía entre 27% (TAT 207) a 1.900 m y 93% (TAT 204) a 1.350 m. Presenta un promedio de 52%.
Arbolitos o subarbóreo
Fluctuó entre 53% (TAT 106, 107) a 1.540 m y 1900 m, y 19% (TAT 208) a 2.140 m. El promedio de cobertura fue 38%.
Arbustivo
Registra valores extremos entre 24% (TAT 201) a 550 m y 61% (TAT 211) a 920 m, con promedio para el estrato de 47%.
Herbáceo
Varía entre 19% (TAT 212) a 1830 m y 65% (TAT 208) a 2140 m. El promedio fue 39%.
Al observar el diagrama estructural (Figura 2), se encuentra que el estrato Arbóreo inferior es el más dominante, mientras el Arbóreo superior sigue siendo el más bajo con 23%.
Distribución de la cobertura (m2) (Figura 3)
Los valores de cobertura se distribuyen en 11 clases, cuyos límites oscilan entre 1,0 y 109,1 m2. La clase I (1,0-10,7 m2) contiene al 78% de los individuos y la clase II (10,8-20,5 m 2) el 14%.
Altura del dosel (Tabla 1, Figura 4)
La altura promedio del dosel es 22 m, con variación entre 12 m (TAT 207) a 1.900 m y 32 m (TAT 212) a 1.830 m.
Distribución de las alturas (tallas) (Figura 5)
Se distribuyen en 11 clases con valores entre 6,0 y 35,4 m. La clase I (6,0-8,8 m) agrupa al 48% de los individuos y la clase II (8,9-11.8 m) el 16%; juntas reúnen el 64% de los individuos totales. Al agrupar clases se diferencian cuatro estratos: el suprimido, con menos de 9 m, contiene el 48% de los individuos; el dominado, entre 9 y 18 m, que retiene el 34%; el codominante, entre 18 y 21 m, con el 8%; y el dominante, entre 21 y 35 m, con el 10%.
Número de individuos en estrato arbóreo
Se registraron 583 individuos en un área de 4.630 m2 , de los cuales 355 tienen DAP > 10 cm.
Arbóreo superior
Se encuentra un promedio de cuatro individuos por levantamiento, con el mayor valor en el levantamiento (TAT 212) a 1.830 m, con 13 individuos.
Arbóreo inferior Varía entre seis individuos (TAT 207,234) a 1.900 m y 2300 m respectivamente, y 29 (TAT 204) a 1.350 m. El promedio es de 16 individuos por levantamiento.
Arbolitos o subarbóreo
El número de individuos fluctúa entre 26 (TAT 208)-a- 2.140 m y 82 (TM 206) a 1.540 m, con un promedio de 47 individuos por lévantamiento.
Densidad absoluta individuos con DAP 210 cm (Tabla 5)
La variación es 0,61 (TAT 207, 210) a 1.900 m y 1.950 m respectivamente, y 1,0 (TAT 209) a 2.140 m.
Distribución del DAP (Figura 6)
El DAP se distribuye en 11 clases con valores entre 1,3 y 166,0 cm. La clase I (1,3-16,2 cm) agrupa el 68% de los individuos y la clase II (16,3-31,2 cm) el 21%; juntas reúnen el 89% de los individuos totales.
Distribución del DAP cada 10 cm en los estratos arbóreos (DAP 2 10 cm), (Figura 7)
El DAP se distribuye en 11 clases con valores entre 1,3 y 166,0 cm. La clase I (1.0-10 cm) agrupa el 45% de los individuos y la clase II (11-20 cm) el 34%; juntas reúnen el 79% de los individuos totales.
Área basal (Tabla 2)
Contiene un total de 24,3 m2 de área basal total, que varía entre 1,8 m2 (TAT 204) a 1.350 y 4,7 m2 (TAT 212) a 1.830. Se estima un promedio de 5,4 m2 por 0,1 ha.
Distribución del área basal por clase diamétrica (Figura 8)
El área basal total fue 24.3 m 2 distribuida en 11 clases con valores entre 1,3 y 166,0 cm. Las clases II (16,3-31,2 cm) con 19% y seguida de la clase IV (46,2-61,1 cm) con 17%, juntas reúnen 33% del área basal.
Índices de valor de importancia IVI
(Tabla 3) y de predominio fisionómico IPF (Tabla 4)
Una especie indeterminada de Akhornea con 3,9% de IVI es la especie con mejor expresión; siguen en importancia Cecropia sp. con 3,7% y Wettinia kalbreyeri con 2,8%. El mayor IPF lo obtuvo Akhornea sp. con 4,8%, por su alta dominancia en cobertura, 4,8%. Cecropia sp. con 3,7% y Wettinia sp. con 3,2%, le siguieron en importancia.
Riqueza
Número de especies
El número total de especies en el grupo fue 418. El número de especies por levantamiento varía entre 45 (TAT 201) a 550 m y 99 (TAT 207) a 1.900 m.
Entre estratos se presentó la siguiente variación:
Arbóreo superior
Se encontraron 18 especies, con máximo de nueve (TAT 212) a 1.830 m.
Arbóreo inferior
Registra 66 especies con máximo de doce (TAT 204, 205, 209) a 1.350 m, 1.650 m y 2.140 m respectivamente.
Subarbóreo o arbolitos
Alcanzó un total de 129 especies, variando entre 13 (TAT 208) a 2.140 y 27 (TAT 207, 209) a 1.900 m y 2140 m.
Arbustivo
183 especies fueron encontradas en este estrato. Los valores varían entre 19 (TAT 201) a 550 m y 44 (TAT 207) a 1.900 m.
Herbáceo El total fue 220 especies, con fluctuación entre 19 (TAT 201) a 550 m y 46 (TAT 206,205) a 1.540 m y 1650m.
Riqueza absoluta (Tabla 5)
En todos los estratos varía entre 1,16 (TAT 212) a 1.830 m y 1,96 (TAT 209) a 2.140 m. En elementos con DAP > 10 cm, la variación es de 0,3 (TAT 204, 206) a 1.350 m y 1.540 m respectivamente, y 0,52 (TAT 209) a 2.140 m.
Índice de valor de importancia para familias (Tabla 6)
Lauraceae (10,5%), Rubiaceae (10,2%) y Moraceae (10%) fueron las familias con mayor IVIF.
Cuatro grupos en asociación se distribuyen en esta región de vida; sus características más sobresalientes, diferenciando entre vertientes, se resumen a continuación.
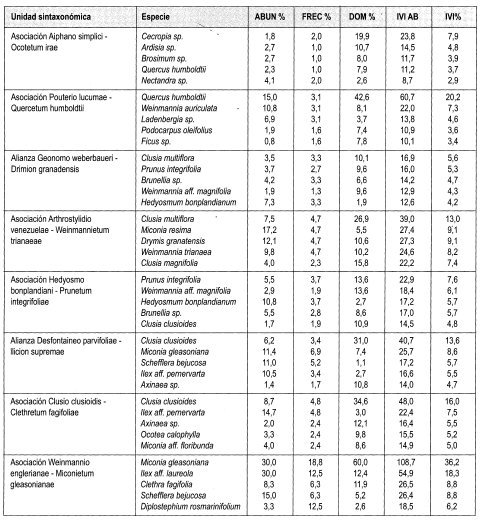
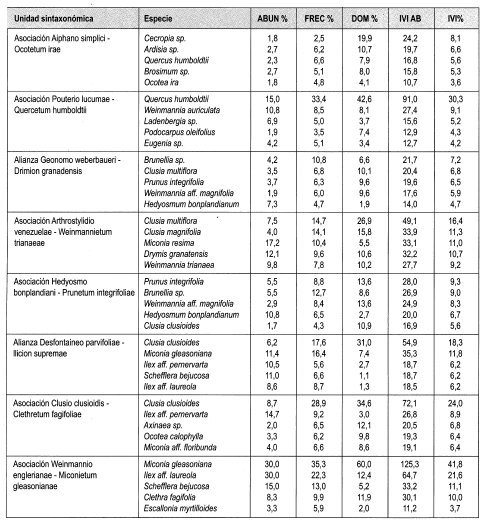
En la vertiente occidental se diferencian tres asociaciones: en la asociación Aiphano simplici-Ocotetum irae, se presenta mayor cobertura en el estrato Arbóreo superior (40%) y mayor altura promedio del dosel (27 m). Las especies más importantes según IVI e IPF son Cecropia sp., Ardisia sp, Brosimum sp. y Quercus humboldtii. Contiene uno de los valores promedio mayores en número de individuos con DAP > 10 cm en 0,1 ha (86) y el mayor promedio de área basal por 0,1 ha (6,6 m 2). La asociación Callicarpo acuminatae-Ficetum hartwegii, presenta mayor dominancia en los estratos Arbóreo inferior (63%) y Subarbóreo (58%). Las especies más representativas según IVI e IPF fueron Licania aff. armeniaca, Madura tinctorea y Guarea cartaguena. En la asociación Pouterio lucumae-Quercetum humboldtii, los mayores valores de IVI e IPF son para Quercus humboldtii, Weinmannia Auriculata y Ladembergia sp. Comparte el mayor número de individuos con DAP > 10 cm en 0,1 ha (86) con Aiphano simplici-Ocotetum irae.
La asociación Guettardo chiriquiensis-Weinmannietum balbisianae, con área de distribución en la vertiente oriental, registra los mayores valores de cobertura en los estratos Arbustivo (47%) y Herbáceo, (50%). El mayor IVI e IPF lo obtienen Akhornea sp., Wettinia kalbreyeri y Ocotea calophylla. Registra el mayor número de especies (203), géneros (136) y familias (56) en todos los estratos.
REGIÓN DE VIDA ANDINA
La vegetación de la alianza Geonomo weberbaueri-Drimion granadensis, representa esta región de vida entre 2.700 m y 3.130 m, en las dos vertientes de la cordillera. Sus características principales son:
Cobertura (Figura 2)
Arbóreo superior
Se obtuvo el valor máximo de 41% (TAT 227) a 3.130 m. El valor promedio fue 14%.
Arbóreo inferior
Varío entre 39% (TAT 230, 218) a 2.700 m y 3.100 respectivamente, y 67% (TAT 216) a 2.740 m. El valor promedio fue 46%.
Arbolitos o subarbóreo
Fluctuó entre 18% (TAT 228) a 2.920 m y 72% (TAT 218) a 3.100 m. El promedio de cobertura fue 52%.
Arbustivo
Los valores extremos fueron de 25% (TAT 230) a 2.700 m y 74% (TAT 227) a 3.130 m, con promedio para el estrato de 38%.
Herbáceo
Varia entre 27% (TAT 230) a 2.700 m y 51% (TAT 228) a 2.920 m. El promedio fue 35%.
El diagrama estructural (Figura 2) muestra que los estratos Arbustivo y Arbóreo inferior son los más representativos con 52% y 48%, respectivamente.
Distribución de la cobertura (m2) (Figura 3)
Los valores de cobertura se distribuyen en diez clases, cuyos límites oscilan entre 1,0 y 70,8 m 2. La clase I (1,0-7,9 m 2) contiene al 74% de los individuos y la clase II (8.0-14,9 m 2) el 14%.
Altura del dosel (Tabla 1, Figura 4)
La altura promedio del dosel es 17 m, con variación entre hm (TAT 228) a 2.920 m y 26 m (TAT 227) a 3.130 m.
Distribución de las alturas (tallas) en estratos arbóreos (Figura 5)
Se distribuyen en 10 clases con valores entre 6,0 y 30,9 m. La clase I (6,0-8,4 m) agrupa al 50% de los individuos y la clase II (8,5-10,9 m) el 18%; juntas reúnen el 68% de los individuos totales. Al agrupar clases se diferencian cuatro estratos: el suprimido, menor de 8 m, contiene el 50% de los individuos; el dominado, entre 8 y 13 m, retiene el 27%; el codominante, entre 13 y 18 m, con el 13%; y el dominante, entre 18 y 31 m, con el 10%.
Número de individuos estrato arbóreo
Se registraron en este grupo un total de 518 individuos mayores de 6 m en un área de 3.558 m2, de los cuales 340 tienen DAP > 10 cm.
Arbóreo superior
Se presenta un promedio de tres individuos por levantamiento, con el mayor valor en el levantamiento 227 a 3.130 m, con 7 individuos.
Arbóreo inferior
Varía entre nueve individuos (TAT 228) a 2.920 m, y 25 (TAT 216) a 2.740 m. El promedio es de 15 individuos por levantamiento.
Arbolitos o subarbóreo El número de individuos fluctúa entre 22 (TAT 228) a 2.920 m y 85 (TAT 216) a 2.740 m,con un promedio de 62 individuos por levantamiento.
Densidad absoluta individuos con DAP 210 cm (Tabla 5)
La variación es de 0,49 (TAT 228) a 2.920 m y 1,43 (TAT 216) a 2.740 m.
Distribución del DAP (Figura 6)
El DAP se distribuye en diez clases con valores entre 2,5 y 92,8 cm. La clase I (2,5-11,4 cm) agrupa el 42% de los individuos y la clase II (11,5-20,5 cm) el 34%; juntas reúnen el 76% de los individuos totales.
Distribución del DAP cada 10 cm en los estratos arbóreos (DAP > 10 cm) (Figura 7)
Los individuos se distribuyen en 10 clases con valores entre 1 y 100 cm. La clase I (1-10 cm) agrupa al 36% de los individuos y la clase II (11-20 cm) el 40%; juntas reúnen el 76% de los individuos totales.
Área basal (Tabla 2)
Contiene un total de 19,3 m 2 de área basal total, que varía entre 1,4 m 2 (TAT 230) a 2.700 m y 4,9 m2 (TAT 227) a 3.130 m. Se estima un promedio de 5,5 m2 por 0,1 ha.
Distribución del área basal por clase diamétrica (Figura 8)
La distribución de 19,3 m2 de área basal se da en 10 clases con valores entre 2,5 y 92,8 cm. La clase VII (56,7-65,7 cm) es la de mayor frecuencia, agrupando al 25% de esta, seguida de la clase II (11,5-20,5 cm) con el 16%; Las dos reúnen el 41% del área basal total.
Índices de valor de importancia IVI (Tabla 3) y de predominio fisionómico IPF (Tabla 4)
Clusia multiflora con 5,6% de IVI es la espe cie con mejor expresión y es además la más dominante con 10,1%; le sigue en importan cia Prunus integrifolia (5,3%) y Brunellia sp. (4,7%). El mayor IPF lo obtuvo Brunellia sp. con 7,2%. Otras especies importantes fueron Clusia multiflora con (6,8%) y Prunus integrifolia (6,5%).
Riqueza
Número de especies
El número total de especies en todos los estratos fue 269. El número de especies por levantamiento varía entre 51 (TAT 227) a 3.130 m y 66 (TAT 228, 217) a 2.920 m y 2.900 m. Según estratos, se presenta la siguiente variación:
Arbóreo superior
Se encontraron 6 especies, con máximo de cuatro (TAT 227) a 3.130 m.
Arbóreo inferior Se registraron 36 especies. Fluctúa entre seis (TAT 228, 217, 218) a 2.920 m, 2900 m y 3100 m respectivamente, y 10 especies (TAT 229) a 2.800 m.
Subarbóreo o arbolitos Se registraron 84 especies. Varía entre 12 (TAT 227) a 3.130 y 24 (TAT 217) a 2.900 m.
Arbustivo
Se encontraron 107 especies, con valores extremos de 18 (TAT 230, 227) a 2.700 y 3130 m respectivamente, y 30 (TAT 217) a 2.900 m.
Herbáceo
El total fue 140, con fluctuación entre 26 (TAT 217) a 2.900 m y 36 (TAT 229) a 2.900 m.
Riqueza absoluta (Tabla 5)
En todos los estratos varía entre 1,02 (TAT 227) a 3.130 m y 1,51 (TAT 216) a 2.740 m. En elementos con DAP > 10 cm, la variación es de 0,28 (TAT 227) a 3.130 m y 0,42 (TAT 230) a 2.700 m.
Índice de valor de importancia para familias (Tabla 6)
El 13,9% del IVIF lo obtuvo Melastomataceae al ser más abundante (16,2%), le sigue en importancia Clusiaceae con 11,3% y Cunoniaceae con 10,1%.
La vegetación de la alianza Geonomo weberbaueri-Drimion granadensis presenta algunas particularidades en las asociaciones que la conforman así:
La asociación Arthrostylidio venezuelae Weinmannietum trianaeae, distribuida en la vertiente oriental, presenta mayor cobertura en el estrato subarbóreo (59%) Las especies más importantes según IVI e IPF son Clusia multiflora, Miconia resima, Drymis granatensis y Clusia magnifolia. Contiene el mayor promedio de número de individuos con DAP > 10 cm en 0,1 ha (100). La asociación Hedyosmo bonplandiani-Prunetum integrifoliae, presenta mayor dominancia en los estratos Arbóreo superior (23%), Inferior (50%), Arbustivo (43%) y Herbáceo (36%) y mayor altura promedio del dosel (19 m). Las especies más representativas según IVI e IPF fueron Prunus integrifolia, Weinmannia aff. magnifolia, Hedyosmum bomplandianum y Brunellia sp. Contiene el mayor promedio de área basal por 0,1 ha (9,6 m 2). Registra el mayor número de especies (217), géneros (133) y familias (69) en todos los estratos.
FRANJA ALTOANDINA
La vegetación de la alianza Desfontaineo parvifoliae-Ilicion supremae representa esta franja entre 3.300 m y 3.700 m. Sus características principales son:
Cobertura (Figura 2)
Arbóreo superior
Se obtuvo el valor máximo de 7% (TAT 219) a 3.300 m. El valor promedio fue 7%.
Arbóreo inferior Varío entre 2% (TAT 220) a 3.500 m y 67% (TAT 226) a 3.320 m. El valor promedio fue 35%.
Arbolitos o subarbóreo Fluctuó entre 17% (TAT 221) a 3.700 m y 84% (TAT 222) a 3.520 m. El promedio de cobertura fue 36%.
Arbustivo Los valores extremos fueron de 32% (TAT 226) a 3.320 m y 137% (TAT 221) a 3.700 m, con promedio para el estrato de 77%.
Herbáceo Varia entre 20% (TAT 221) a 3.700 m y 47% (TAT 226) a 3.320 m. El promedio fue 32%. El diagrama estructural (Figura 2), muestra el estrato Arbustivo más dominante con 67% de cobertura.
Distribución de la cobertura (m2)(Figura 3)
Los valores de cobertura se distribuyen en nueve clases, cuyos límites oscilan entre 1,0 y 52,4 m2. La clase I (1,0-6,6 m2) contiene al 80% de los individuos y la clase II (6,7-12,4 m2) el 12%.
Altura del dosel (Tabla 1, Figura 4)
La altura promedio del dosel es 10 m, con variación entre 6 m (TAT 223, 221, 220) a 3.700 y 3.500 m, y 19 m (TAT 226) a 3.320 m.
Distribución de las alturas (tallas)(Figura 5)
Se distribuyen en nueve clases con valores entre 6,0 y 29,5 m. La clase I (6,0-8,5 m) agrupa el 64% de los individuos y la clase II (8,6-11,2 m) el 14%; juntas reúnen el 78% de los individuos totales. Al agrupar clases se diferencian cuatro estratos: el suprimido, menor a 9 m, contiene el 64% de los individuos; el dominado, entre 9 y 14 m, retiene el 20%; el codominante, entre 14 y 19 m, con el 10%; y el dominante, entre 19 y 30 m, el restante 6%.
Número de individuos en estrato arbóreo
Se registraron en este grupo un total de 210. individuos mayores de 5 m en un área de 1.040 m2, de los cuales 130 tienen DAP > 10 cm.
Arbóreo superior
Se presenta un promedio de dos individuos por levantamiento, con el mayor valor en el levantamiento 219 a 3.300 m, con dos individuos.
Arbóreo inferior
El máximo valor encontrado fue 16 (TAT 219) a 3.300 m. El promedio es de 11 individuos por levantamiento.
Arbolitos o subarbóreo
El número de individuos fluctúa entre seis (TAT 221) a 3.700 m y 44 (TAT 220) a 3.500 m, con un promedio de 29 individuos por levantamiento.
Densidad absoluta individuos con DAP 210 cm (Tabla 5)
La variación es de 0,42 (TAT 221) a 3.700 m y 2,13 (TAT 226) a 3.320 m.
Distribución del DAP (Figura 6)
El DAP se distribuye en nueve clases con valores entre 3,2 y 119,1m. La clase I (3,2-16,0 cm) agrupa al 72% de los individuos y la clase II (16,1-28,9 cm) el 12%; juntas reúnen el 84% de los individuos totales.
Distribución del DAP cada 10 cm en los estratos arbóreos (DAP 2 10 cm) (Figura 7)
Los individuos se distribuyen en 12 clases con -valores entre 1 y 120 cm. La clase I (1-10 cm) agrupa al 40% de los individuos y la clase II (11-20 cm) el 38%; juntas reúnen el 78% de los individuos totales.
Área basal (Tabla 2)
Contiene un total de 10,4 m 2 de área basal total, que varía entre 0,1 m 2 (TAT 221) a 3.700 m y 5,4 m2 (TAT 226) a 3.320 m. Se estima un promedio de 5,5 m2 por 0,1 ha.
Distribución del área basal por clase diamétrica (Figura 8)
La distribución de 10,4 m2 de área basal se da en 9 clases con valores entre 3,2 y 119,1 cm. La clase VII (80,5-93,3 cm) es la de mayor frecuencia, agrupando al 18% de esta, seguida de la clase VI (67,7-80,4 cm) con el 7%; Las dos reúnen el 25% del área basal total.
Índices de valor de importancia ¡VI (Tabla 3) y de predominio fisionómico IPF (Tabla 4)
Clusia clusioides con 13,6% de ¡VI es la es pecie con mejor expresión; además es la más dominante (31.0%); le sigue en importancia Miconia gleasoniana (8,6%) y Scheffiera bejucosa (5,7%). El mayor IPF lo obtuvo Clusia clusioides (18,3%). Otras especies importantes fueron Miconia gleasoniana (11.8%) y Ilex aff pernervarta (6.2%).
Riqueza
Número de especies
El número total de especies en todos los estratos fue 172. El número de especies por levantamiento varía entre 37 (TAT 22772) a 3.520 m y 69 (TAT 226) a 3.320 m. Según estratos, se presenta la siguiente variación:
Arbóreo superior
Se encontraron dos especies, con máximo de dos (TAT 219) a 3.300 m.
Arbóreo inferior
Se registraron 15 especies. Fluctúa entre dos (TAT 220) a 3.500 m, y ocho especies (TAT 229) a 2.800 m.
Subarbóreo o arbolitos
Hay 84 especies en este estrato. Varía entre 12 (TAT 227) a 3.130 y 24 (TAT 219) a 3.300 m.
Arbustivo
Se encontraron 41 especies. Los valores extremos fueron de cuatro en el TAT (221) a 3.700 my 15 en el TAT 226 a 3.320 m.
Herbáceo
El total fue 102, con fluctuación entre 24 (TAT 223) a 3.700 m y 40 (TAT 226) a 3.320 m.
Riqueza absoluta (Tabla 5)
En todos los estratos varía entre 1,83 (TAT 219) a 3.300 m y 4,6 (TAT 226) a 3.320 m. En elementos con DAP > 10 cm, la variación es de 0,25 (TAT 222, 223) a 3.520 m y 3.700 m respectivamente, y 0,87 (TAT 226) a 3.320 m.
Índice de valor de importancia para familias (Tabla 6)
El 25% del IVIF lo obtuvo Melastomataceae al ser más abundante (26,7%); le sigue en importancia Aquifoliaceae con 13,7% y Araliaceae con 8.8%.
Cuatro grupos en asociación se distribuyen en las dos alianzas; sus características más sobresalientes se resumen a continuación.
En las asociaciones que se agrupan en la alianza Desfontaineo parvifoliae-Ilicion supremae, se presentan algunas particularidades.
En la asociación Clusio clusioidis-Clethretum fagifoliae son más representativos los estratos Arbóreo inferior (35%) y Herbáceo (36%). Registra mayor altura promedio del dosel del3 m en la alianza. Los mayores valores de IVI e IPF son para Clusia clusioides, Ilex aif pernervata y Axinaea sp. Contiene el mayor promedio de número de individuos con DAP > 10 cm en 0,1 ha (143) y el mayor promedio de área basal por 0,1 ha (35,3 m2). Registra el mayor número de especies (130), géneros (87) y familias (47) en todos los estratos. La asociación Weinmannio englerianae-Miconietum gleasonianae registra el mayor promedio de cobertura en los estratos Subarbóreo (43%) y Arbustivo (87%). El mayor IVI e IPF lo obtienen Miconia gleasoniana, Ilex aff. laureola, Clethra fagifolia y Scheffiera bejucosa.
DISCUSIÓN Y CONCLUSIONES
VARIACIONES EN ESTRUCTURA,RIQUEZA Y DIVERSIDAD VEGETAL EN EL TRANSECTO DE TATAMÁ
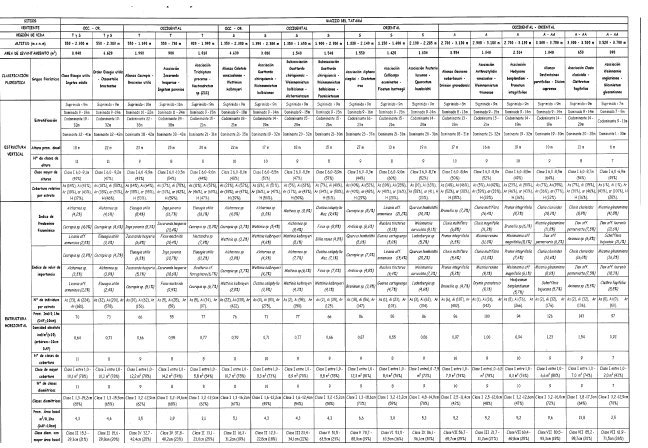
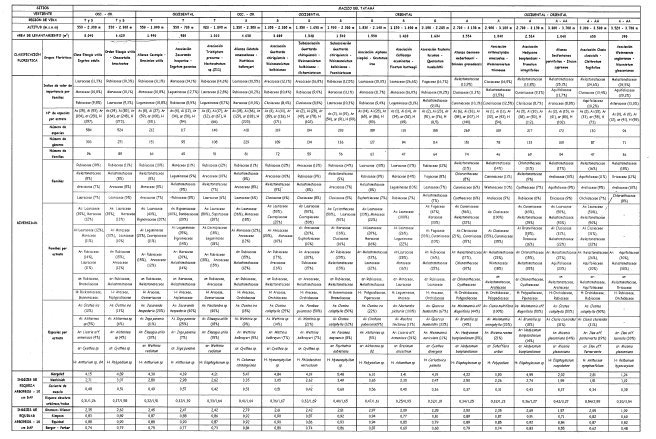
Las características climáticas y edáficas regionales permiten detectar variaciones en la estructura de la vegetación por efecto de la disimetría (exposición) de las dos vertientes y del gradiente altitudinal (regiones de vida).
Vertiente occidental
En la región de vida tropical, la vegetación está definida por el estrato Arbóreo superior con individuos emergentes mayores de 30 m y 14% de cobertura; por el Arbóreo inferior, el dominante, con 64% de cobertura; y por una altura promedio del dosel de 23 m. Las especies más importantes fueron Elaeagia utilis, Jacaranda hesperia, Inga pavonia y Cecropia sp. Las familias con mayor IVIF fueron Rubiaceae, Moraceae y Leguminosae. En 0,1 ha contiene un promedio de 66 individuos y 3,5 m 2 de área basal en elementos con DAP > 10 cm.
En la región de vida subandina se presenta una cobertura escasa en el estrato Arbóreo superior (6%) y máximo valor en el Arbóreo inferior (51%); la altura promedio del dosel es 15 m. Las especies más importantes son Alchornea sp., Wettinia kalbreyeri, Ocotea calophylla y Wettinia sp., mientras que las familias con mayor IVIF fueron Arecaceae, Rubiaceae y Clusiaceae. Contiene un promedio de 71 individuos con DAP > 10 cm en 0,1 ha, que registran un área basal de 4,3 m 2.
En la región de vida andina se registra mayor valor de cobertura en los estratos Arbóreo inferior (50%) y Subarbóreo (46%), con el valor mayor en la vertiente para el Arbóreo superior (23%). La altura promedio del dosel es 19 m. Los mayores valores de importancia lo obtienen Prunus integrifolia, Weinmannia aff. magnifolia, Hedyosmum bomplandianum y Brunellia sp., en las familias, Melastomataceae, Cunoniaceae y Clusiaceae. Contiene un promedio de 94 individuos con DAP > 10 cm en 0,1 ha con un área basal de 5,2 m2.
En la franja altoandina, los estratos Arbóreo superior e Inferior están ausentes y se registra el máximo valor de cobertura en el estrato Arbustivo (87%). La altura promedio del dosel es 6 m. Las especies más importantes fueron Miconia gleasoniana, Ilex aff. laureola, Clethra fagifolia y Schefflera bejucosa, mientras que en familias el mayor IVIF fue para Melastomataceae, Aquifoliaceae y Asteraceae. Un promedio de 97 individuos con DAP > 10 cm en 0,1 ha, contienen un área basal de 2,5 m2.
Vertiente oriental
En la región de vida subandina, la vegetación en su parte baja presenta el estrato subarbóreo más representativo con 40% de cobertura en promedio, mientras que el Arbóreo superior es poco dominante (10%). La altura promedio del dosel es 13 m. Los mayores valores de IVI e IPF lo obtienen Licania (J. armeniaca, Madura tinctorea y Guarea cartaguena, con Lauraceae, Moraceae y Leguminosae como familias importantes. Contiene un promedio de 55 individuos con DAP > 10 cm en 0,1 ha, con área basal de 3,0 m2; en su parte media, presenta altos valores de cobertura en los estratos arbóreos, con 40% en el Superior y 52% en el Inferior; el estrato Herbáceo es el de menor valor con 29%; la altura promedio del dosel es 27 m, el valor más representativo para el transecto del Tatamá. Las especies más importantes fueron Cecropia sp., Ardisia sp., Brosimum sp. y Quercus humboldtii, mientras que Lauraceae, Moraceae y Rubiaceae tuvieron mayor IVIF. Contiene un promedio de 86 individuos con DAP > 10 cm en 0,1 ha y un área basal de 6,6 m2; en su parte alta, presenta el estrato subarbóreo más representativo con 58% de cobertura en promedio, mientras que el Arbóreo superior es poco dominante (10%). La altura promedio del dosel es 19 m. Quercus humboldtii, Weinmannia Auriculata y Ladembergia sp. son las especies de mayor valor en estructura; Fagaceae, Rubiaceae y Melastomataceae son las familias importantes. Contiene un promedio de 86 individuos con DAP > 10 cm en 0,1 ha y un área basal de 5,3 m2.
En la región de vida andina se registra una cobertura escasa en el estrato Arbóreo superior (5%), con el máximo valor en el Subarbóreo (59%). La altura promedio del dosel es 16 m. Los índices más altos lo obtienen Clusia multiflora, Miconia resima, Drymis granatensis y Clusia magnifolia; las familias importantes son Clusiaceae, Melastomataceae y Cunoniaceae. Contiene un promedio de 100 individuos con DAP > 10 cm en 0,1 ha y un área basal de 5,2m2.
En la franja altoandina se registra el máximo valor de cobertura en el estrato Arbustivo (46%) y el más bajo en el Arbóreo superior (7%). La altura promedio del dosel es 13 m. Son importantes Clusia clusioides, Ilex aif pernervata y Axinaea sp.; en las familias Melastomataceae, Aquifoliaceae y Asteraceae. Contiene un promedio de 143 individuos con DAP > 10 cm en 0,1 ha y un área basal de 13,8 m 2, el valor más alto en el transecto.
Al comparar las dos vertientes, se encuentran algunas afinidades y diferencias estructurales en los grupos florísticos que las integran.
En las dos vertientes, el número promedio de individuos con DAP > 10 cm en 0,1 ha aumenta a medida que aumenta la altitud y es ligeramente mayor en la vertiente oriental. La distribución en clases de frecuencia de la altura, cobertura y diámetro a la altura del pecho (DAP) muestra que la mayoría de los individuos se agrupan en la primera clase: entre el 72 y 81% de los individuos tienen cobertura relativa entre 1 y 12 m 2; entre el 44 y 68% de los individuos arbóreos tienen alturas entre 6 y 12,1m; entre el 47 y 73% de los individuos arbóreos tienen DAP entre 1,3 y 18,6 cm. Los valores de área basal para individuos con DAP > 10 cm en 0,1 ha, aumentan en el gradiente altitudinal hacia la región de vida Andina, y es más marcada esta variación en la vertiente occidental. La riqueza absoluta para individuos con DAP > 10 cm fue mayor en la región de vida subandina (0,46 y 0,35). En la región de vida andina las familias más importantes en IVIF fueron Melastomataceae, Cunoniaceae y Clusiaceae, mientras que en la franja altoandina lo fueron Melastomataceae, Aquifoliaceae y Asteraceae.
En la vertiente occidental se presentan los mayores valores promedio de cobertura en los estratos arbóreos, a excepción de la franja Altoandina, donde no se registran individuos de los estratos Arbóreo superior e inferior, condición que determina en esta franja, la presencia de un estrato Arbustivo muy dominante y un valor bajo en el área basal para individuos con DAP > 10 cm. Las familias más importantes en la región de vida tropical fueron Rubiaceae, Moraceae y Leguminosae y en la subandina Arecaceae, Rubiaceae y Clusiaceae,
En la vertiente oriental, la región de vida subandina en su parte media (bosque de Lauraceas), registra elevada dominancia del estrato Arbóreo superior (40%), mientras que en la franja más alta (>2.100 m) (robledal) y hasta la región de vida andina, es muy representativo el estrato subarbóreo. La altura promedio del dosel en las regiones de vida subandina, Andina y la franja altoandina, registra mayores valores que en la vertiente occidental. Las familias más importantes en la región de vida subandina son Lauraceae, Moraceae, Leguminosae y Rubiaceae en las partes baja y media; y Fagaceae, Rubiaceae y Melastomataceae en la parte alta.
En síntesis, se evidencian cambios marcados al relacionar las vertientes en el transecto de Tatamá. El clima, específicamente la precipitación, y en menor medida el suelo y la topografía se consideran factores influyentes en la diferenciación estructural de la vegetación.
En la vertiente occidental, los mayores valores promedio de cobertura en los estratos arbóreos están estrechamente relacionados con los valores mayores de precipitación. No obstante, los valores bajos en la altura promedio del dosel y del área basal para individuos con DAP > 10 cm pueden estar relacionados con la presencia de suelos poco evolucionados (Inceptisoles) que, aunque presentan cenizas volcánicas, tienen baja saturación de bases (Typic Dystrandept) o tendencia a horizontes muy débiles (Entic Dystran'clept), sometidos al constante lavado (lixiviación) por acción de la elevada precipitación y la topografía inclinada.
En la vertiente oriental, aunque con precipitaciones más bajas que en la vertiente occidental, en la región de vida subandina los suelos poco evolucionados (Inceptisoles), de superficiales a muy superficiales (Lithic Humitropept), se desarrollan con alta presencia de materia orgánica, reflejando en los grupos de vegetación una mayor altura del dosel, mayores valores de área basa y mayor densidad de individuos arbóreos.
En la franja altoandina de las dos vertientes, con áreas de menor pendiente, con suelos poco evolucionados, con presencia de cenizas volcánicas, húmedos, con alto contenido de materia orgánica y con alto grado de descomposición, la vegetación muestra mayor dominancia del estrato arbustivo y menor altura del dosel con relación al transecto.
Los valores de área basal para individuos con DAP > 10 cm en 0,1 ha, aumentan en el gradiente -altitudinal hacia la región de vida andina.
La distribución en clases de frecuencia de la altura, cobertura y diámetro a la altura del pecho (DAP) muestra que la mayoría de los individuos se agrupan en la primera clase.
En las dos vertientes, la riqueza absoluta para individuos con DAP > 10 cm fue mayor en la región de vida subandina (0,46 y 0,35). En la Tabla 7 se presenta una síntesis de la variabilidad de las características de la estructura, la riqueza y la diversidad florística.

AGRADECIMIENTOS
La fase de campo se desarrolló en el marco del programa Ecoandes. A todos los colegas que participaron en las excursiones de campo, nuestro sincero agradecimiento.
REFERENCIAS BIBLIOGRÁFICAS
- Arellano, II. 2001. Estudio de la vegetación del sistema andino de la serranía del Perijá. Tesis de pregrado. Departamento de Biología. Universidad Nacional de Colombia. Bogotá. 160 p.
- Cantillo, E.E. 2001. Caracterización estructural, ordenación y dinámica de la vegetación en la zona de captación de aguas de la microcuenca El Tigre, municipio de Norcasia, Caldas. Tesis de Maestría. Universidad Nacional de Colombia. 178 p.
- Cantillo, E.E., K. Rodríguez y A. Avella. 2005. La vegetación leñosa de la Reserva Forestal Cárpatos, Guasca, Cundinamarca. Centro de Investigaciones y Desarrollo Científico. Universidad Distrital Francisco José de Caldas. Fondo de Publicaciones. 109 pp. Bogotá.
- Cantillo, E.E. y J.O. Rangel-Ch. (En imprenta). Aspectos de la estructura y del patrón de riqueza de la vegetación del transecto Sumapaz. En: T. van der Hammen & A. Cleef (eds). La Cordillera oriental (Colombia). Transecto Sumapaz. Estudios de Ecosistemás Tropandinos-Ecoandes.
- Cantillo, E.E., J.O. Rangel-Ch & II. Arellano. (En imprenta). Aspectos de la estructura y del patrón de riqueza de la vegetación de la Serranía del Perijá. Instituto de Ciencias Naturales. Universidad Nacional de Colombia. Bogotá.
- Duivenvoorden, J.F. & J.M. Lips. 1993. Ecología del paisaje del medio Caquetá. Estudios de la Amazonia Colombiana. Tropenbos. Colombia. Bogotá. 301 p.
- Florez, A. 1987. Geomorfología estructural y dinámica morfogenética del transecto Sumapaz (Cordillera oriental). Análisis Geográficos 12. Bogotá. 50 p.
- Flórez, A. 2005. Aspectos geomorfológicos del área del transecto Tatamá. En: T. vander Hammen, J.O. Rangel-Ch & A. Cleef(eds.). La cordillera occidental colombiana. Transecto Tatamá. Estudios de Ecosistemas Tropandinos-Ecoandes 6; 797-812. J. Cramer, Berlín-Stuttgart.
- Lincoln R.J., G.A. Boxshall & P.F. Clark. 1986. Diccionario de ecología, evolución y taxonomía. Fondo de Cultura Económica. México.
- Mori, 5. & B. Boom. 1987. The Lecythidaceae of the lowland neotropical forest: La Fumeé Montain, French Guiana. Mem. New York Bot, Garden 44: 9-29.
- Moreno, C.E. 2001. Métodos para medir la biodiversidad. Manuales y Tesis SEA. Vol 1. CYTED, ORCYT/UNESCO & SEA. Zaragoza. 84 p.
- Patiño, R.D. 2003. Estudio de la flora y la vegetación del bosque en la Estación de Primates Colosó, Sucre. Tesis de Maestría. Instituto de Ciencias Naturales. Universidad Nacional de Colombia. Bogotá. 114 pp.
- Rangel-Ch, J.O. y G. Lozano-C. 1986. Un perfil de vegetación entre La Plata (Huila) y el Volcán Puracé. Caldasia 14 (68-70): 533-547.
- Rangel-Ch., J.0., J. Aguirre-C & R. Sánchez-M. 1990. Aspectos estructurales, dinámicos y fisionómicos de la vegetación.En: J. Aguirre & J.O. Rangel-CH. (eds).Biota y ecosistemas de Gorgona. Fondo
- FEN Colombia. Bogotá. Pp. 127-151.
- Rangel-Ch, J.0.. 1991. Vegetación y ambiente en tres gradientes montañosos de Colombia. Tesis de Doctor, Universidad de Ámsterdam. Amsterdam. 349 pp.
- Rangel-Ch., J.O. y A. Garzón. 1994. Aspectos de la estructura, de la diversidad y de la dinámica de la vegetación del Parque Regional Natural Ucumari. En: J.O. Rangel-Ch. (ed.). Ucumarí: Un caso típico de la diversidad biótica andina: 59:84. Publicaciones de la CARDER. Pereira.
- Rangel-Ch, J.0., y A. Velásquez. 1997. Métodos de estudio de la vegetación. En J.O. Rangel-Ch, P. Lowy y M. Aguilar. Colombia Diversidad Biótica II. P: 59-87. Instituto de Ciencias Naturales. Universidad Nacional de Colombia. Bogotá.
- Rangel-Ch., J.0., A.M.Cleef & S. Salamanca. 2003. The ecuatorial interandean and subandean forest of the Parque Los Nevados Transect, Cordillera Central, Colombia. En: T. van der Hammen & A. Dos Santos (eds). La Cordillera Central Colombiana. Estudios de Ecosistemas Tropandinos 5:143-204. J. Cramer, Berlín-Stuttgart.
- Rangel-Ch, J.0.. 2005. Aspectos de la estructura y de la riqueza de especies en la vegetación de los transectos del Tatamá. En: T. Van der Hammen, J. O. Rangel-Ch & A. Cleef (eds). La cordillera occidental colombiana. Transecto Tatamá. Estudios de Ecosistemás Tropandinos-Ecoandes 6; 797-812. J. Cramer, Berlín-Stuttgart.
- Rangel-Ch, J.0., A. M. Cleef, S. Salamanca y C. Ariza. 2005a. La vegetación de los bosques y selvas del Tatámá. En: T. Vander Hammen, J.O. Rangel-Ch & A. Cleef (eds). La cordillera occidental Colombiana. Transecto Tatamá. Estudios de Ecosistemás Tropandinos-Ecoandes 6; 469-643. J. Cramer, Berlín-Stuttgart.
- Rangel-Ch, J.0., M. Aguilar-P. y A. Flórez. 2005b. Clima en el área del transecto Tatamá. En: T. Van der Hammen, J.O. Rangel & A. Cleef (eds). La cordillera occidental colombiana. Transecto Tatamá. Estudios de Ecosistemás Tropandinos-Ecoandes 6; 47-103. J. Cramer, Berlín-Stuttgart.
- Ruiz-B, E. 2005. Climosecuencia altitudinal de los suelos del transecto Tatamá. En: T. Van der Hammen, J.O. Rangel & A. Cleef(eds). La cordillera occidental colombiana. Transecto Tatamá. Estudios de Ecosistemás Tropandinos-Ecoandes 6; 155-223. J. Cramer, Berlín-Stuttgart.
- Sturges H.G. 1926. The Choice of class interval. Joumal of the American Statistical Association. 21:65-66
- UNESCO/PNUMA/FAO. 1980. Ecosistemas de los Bosques Tropicales. UNESCO/CIFCA. Madrid. Pp 126-162.
- Van der Hammen, T. 2005. Introduction: The study of the Tatamá transect. En: T. vander Hammen, J. O. Rangel-Ch & A. Cleef (eds). La cordillera occidental colombiana. Transecto Tatamá. Estudios de Ecosistemas Tropandinos-Ecoandes 6; 797-812. J. Cramer, Berlín-Stuttgart.
Creation date:




Licencia
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






