DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2011.1.a01Published:
2011-01-01Issue:
Vol. 14 No. 1 (2011): January-JuneSection:
Scientific articleAlometría y crecimiento de seis especies arbóreas en un bosque de tierra firme en la amazonia colombiana
Allometry and growth of six tree species in a terra firme forest in colombian amazonia
Keywords:
climate change, wood specific gravity, functional groups, PNN Amacayacú (en).Keywords:
cambio climático, densidad de la madera, grupos funcionales, PNN Amacayacú (es).Downloads
References
Arias, G. J. C., A. J. Duque & L. D. Cárdenas. 2009. Crecimiento diamétrico de un bosque del Noroccidente amazónico. Revista Colombia Amazónica 2: 57-64.
Aróstegui, A. 1982. Recopilación y análisis de estudios tecnológicos de maderas peruanas. Documento de trabajo No. 2. Proyecto PNUD/ FAO/PER/81/2002 Fortalecimiento de los programas de desarrollo forestal en selva central. Lima. 57 p.
Baker, T.R., O.L. Phillips, S. Almeida, L. Arroyo, A. Difiore, T. J. Killeen, S. G. Laurance, W. F. Laurance, S. L. Lewis, J. Lloyd, A. Monteagudo, D. A. Neill, S. Patiño, N. C. A. Pitman, J. N. M. Silva & R. V. Martínez. 2004. Variation in Wood density determines spatial patterns in Amazonian forest biomass. Global Change Biology 10: 545-562.
Barreto, J. S., A. Duque, D. Cárdenas & F. Moreno.2010. Variación florística a escala local en un bosque de tierra firme en la Amazonia Colombiana. Acta Amazónica 40:179-188
Brown, S., A. Gillespie & A. E. Lugo. 1989. Biomass estimation methods for tropical forests with applications to forest inventory data. Forest Science 35: 881-902.
Condit, R., S. P. Hubbell & R. B. Foster. 1996. Assessing the response of plant functional types to climate change in tropical forest. Journal of Vegetation Science 7: 405-416.
Condit, R. 1998. Tropical Forest census plots. Springer-Verlag and R.G. Landes Company. Berlin, Heidelberg, New York. 211 p.
Condit, R., S. Aguilar, A. Hernandez, R. Perez, S. Lao, G. Angehr, S. P. Hubbell & R. B. Foster, 2004. Tropical Forest dynamics across a rainfall gradient and the impact of the Niño dry season. Journal of tropical Ecology 20: 51-72.
Chamorro, C. 1989. Biología de los suelos del Parque Nacional Natural Amacayacu y zonas adyacentes. Amazonas, Colombia. Revista Colombia Geográfica 15: 45-63.
Chave, J. 1999. Study of structural, successional and spatial patterns in tropical rain forests using TROLL, a spatially explicit forest model. Ecological Modelling 124: 233-254.
Chave, J., C. Andalo, S. Brown, M. A Cairns, J. J. Chambers, D. Eamus, H. Folster, D. F. Fromar, N. Higuchi, T. Kira, J. J. Lescure, B. W. Nelson, H. Ogawa, H. Puig, B. Riera & T. Yamacura. 2005. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Ecosystem Ecology 145: 87-99.
Delgado, L. A., M. F. Acevedo, H. Castellanos, H. Ramirez & J. Errano. 2005. Relaciones alométricas y patrones de crecimiento para especies de árboles de la reserva forestal Imataca, Venezuela. Interciencia 30: 275-283.
Detienne, P. & P. Jacquet. 1983. Atlas d'Identification des Bois de l'Amazonie et des Regions Voisines. Centre Technique Forestier Tropical, Nogent-sur-Marne. France. 640 p.
Duivenvoorden, J. F. & J. M. Lips. 1995. A land-ecological study of soil, vegetation, and plant diversity in Colombian Amazonia. Tropenbos Series 12. The Tropenbos Foundation, Wageningen. 438 p.
Enquist, B. J., J. H. Brown & G.B. West. 1998. Allometric scaling of plant energetics and population density. Nature 395: 163-165.
Fearnside, P. M. 1997. Wood density for estimating forest biomass in Brazilian Amazonia. Forest Ecology and Management 90: 59-87.
Hoorn, C. 1994. An environmental reconstruction of the palaeo-Amazon river system (Middle to late Miocene, NW Amazonia). Palaeogeography, Palaeoclimatology, Palaeoecology 112: 187-238.
Huxley, J. S. 1932. Problems of Relative Growth. MethuenLondres. 276 p.
Instituto Brasileiro de Desenvolvimento Florestal -IBDF-. 1981. Madeiras de Amazônia, caracteristicas e utilização, Vol. 1. Floresta Nacional do Tapajós. Brasilia. 113 p.
Kraft, N. J. B., R. Valencia & D. D. Ackerly. 2008. Functional traits and niche-based tree community assembly in an Amazonian forest. Science 322: 580-582.
Keeling, C. H., R. T. Baker, V. R. Martinez, A. Monteagudo & L. O. Phillips. 2008. Contrasting patterns of diameter and biomass increment across tree functional groups in Amazonian forests. Oecología 158: 521-534.
Ketterings, Q. M., R. Coe & M. Van Noordwijk. 2001. Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests. Forest Ecology and Management 146: 199-209.
King, D. A. 1996. Allometry and life history of tropical trees. Journal Tropical Ecology 12: 25-44.
King, D. A., S. J. Davies, M. N. Supardi & S. Tan. 2005. Tree growth is related to light interception and Wood density in to mixed dipterocarp forests of Malaysia. Functional Ecology 19: 445-453.
Laurance, W.F., A. Alexandre, S. Oliveira, G.W. Laurance, R. Condit, S. Henrique, E. M. Nascimento, A. C. Sanchez-Thorin, T E. Lovejoy, S. Ana C. Andrade, S. Agra, J. E. Ribeiro, L.S. Christopher & W. Dick. 2004. Pervasive alteration of tree communities in undisturbed Amazonian forests. Nature 428: 171-175.
Lorenzi, H. 1992. Árboles brasileiras: Manual de identificação e cultivo de plantas arbóreas nativas do Brasil. Instituto Plantarum de Estudos da Flora Ltda-Nova Odessa. São Pablo.
Muller-Landau, H. C. 2004. Interspecific and Inter-site Variation in Wood Gravity of Tropical Trees. Biotropica 36: 20-32.
Nebel, G., J. Dragsted & A. S. Vegas. 2001. Litter fall, biomass and net primary production in flood plain forest in the Peruvian Amazon. Forest Ecology and Management 150: 93-102.
Niklas, K. J. 1994. Plant allometry: The scaling of form and process. University of Chicago Press. Chicago. 395 p.
Niklas, K. J. 1995. Size-dependent allometry of tree height, diameter and trunk- taper. Annals of Botany 75: 217-227.
Nogueira, E. M., B. W. Nelson & P. M. Fearnside. 2005. Wood density in dense forest in central Amazonia. Brazil. Forest Ecology and Management 208: 261-286.
Ogawa, H., K. Yoda, K. Ogino & T. Kira. 1965. Comparative ecological studies on three main types of forest vegetation in Thailand II Plant biomass. Nature an Life Southeast Asia 4: 49-80.
Patiño, S., J. Lloyd, R. Paiva, C. A. Quesada, L. M. Mercado, T. R. Baker, C. I. Czimczik, M. Schwarz, J. Schmerler, A. Sota, A. Santos, V. Horna, J. Peacock, M. Wagner, L. Arroyo, S. Almeida, E. Alvarez, A. Aguilar, D. Bonal, J. Gallo, R. Herrera, N. Higuchi, E. J. Hoyos, E. M. Jimenez, T. Killeen, E. Leal, F. Luizaño, Y. Malhi, P. Meir, A. Monteagudo, D. Neill, V. P. Nuñez, W. Palomino, A. Peña-Cruz, M.C. Peñuela, O.L. Phillips, N. L. Pitman, F. N. Priante, A. Prieto, S. N. Panfil, A. Rudas, R. Salomao, N. Silva, M. Silveira, A. Lezama, J. D. Turriago, R. Vasquez-Martinez, I. Vieira, B. Villanueva & P. Vitzthum. 2008. Branch xylem density variation across the Amazon Basin. Biogeosciences Discuss 5: 2003-2047.
Phillips, O. L., L. Aragão & S. Lewis. 2009. Drought Sensitivity of the Amazon Rainforest. Science 323: 1344-1347.
Prieto, A. 1994. Análisis estructural florístico y de la vegetación de la isla Mocagua, río Amazonas. Amazonas, Colombia. Tesis. Universidad Nacional de Colombia. Bogotá.
Rudas, L. A. 1996. Estudio florístico de la vegetación del Parque Nacional Natural Amacayacu. Amazonas, Colombia. Tesis de MSc. Universidad Nacional de Colombia. Bogotá.
Saldarriaga, J. G., D. C. West, M. L. Tharp & C. Uhl. 1988. Long-term chronosequence of forest succession in the Upper Rio Negro of Colombia and Venezuela. Journal Ecology 76: 938-958.
Suzuki, E. 1999. Diversity in specific gravity and water content of wood among Bornean tropical rainforest trees. Ecological Research 14: 211-224.
Sposito, T. C. & F. A. M. Santos, 2001. Sacling of stem and crown in eight Cecropias (Cecropiaceae) species of Brazil. American Journal of Botany 88: 939-949.
Swaine, M. D. & T. C. Whitmore. 1988. On the definition of ecological spices in tropical rain forests. Vegetation 75: 81-86.
Ter Steege, H., N. C. A. Pitman, O.L. Phillips, J. Chave, D. Sabatier, A. Duque, J. F. Molino, M. F. Prévost, R. Spichiger, H. Castellanos, P. Von Hildebrand & R. Vásquez. 2006. Continental-scale patterns of canopy tree composition and function across Amazonia. Nature 443: 444-447.
Thompson, D. W. 1917. On Growth and Form. Cambridge University Press. Cambridge.793 p.
Torres, A. J. 2006. Densidad de madera en cuatro tipos de bosque localizados sobre suelos contrastantes en la región de Leticia, Amazonia Colombiana. Departamento de ciencias biológicas, Facultad de Ciencias, Universidad de los Andes. Bogotá. 55 p.
Turner, I. M. 2001. The ecology of trees in the tropical rain forest. Cambridge University Press. Cambridge. 290 p.
West, G.B., J. H. Brown & B. J. Enquist. 1997. A general model for the origin of allometric scaling laws in biology. Science 276: 122-126.
Woodcock, D. W. 2000. Wood specific gravity of trees and forest types in the Southern Peruvian Amazon. Acta Amazónica 30: 589-599.
Wright, S. J. 2005. Tropical forest in a changing environment. Trends in Ecology Evolution 20: 553-560.
Zimmerman, J. K., E. M. Everham II, R. B. Waide, D. J. Lodge, C. M. Taylor & N. V. L. Brokaw, 1994. Responses of tree species to hurricane winds in subtropical wet forest in Puerto Rico: Implications for tropical tree life histories. Journal Ecology 82: 911-922.
How to Cite
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Download Citation
 |
Giraldo-Pamplona, W.A., Cárdenas, C., & Duque-Montoya, A.J. (2011). Alometría y crecimiento de seis especies arbóreas en un bosque de tierra firme en la amazonia colombiana. Colombia Forestal, 14(1), 9-21. |
ALOMETRÍA Y CRECIMIENTO DE SEIS ESPECIES ARBÓREAS EN UN BOSQUE DE TIERRA FIRME EN LA AMAZONIA COLOMBIANA
Allometry and growth of six tree species in a terra firme forest in colombian amazonia
Alometria e crescimento de seis espécies de árvores num bosque de terra firme na floresta amazônica colombiana
Wilson A. Giraldo Pamplona1, Dairon Cárdenas2 & Álvaro J. Duque Montoya1
1Departamento de Ciencias Forestales, Universidad Nacional de Colombia sede Medellín. wagiral0@unal.edu.co, ajduque@ unal.edu.co.
2Instituto Amazónico de Investigaciones Científicas SINCHI. dcardenas@sinchi.org.co
Recepción: Marzo 6 de 2010/Aprobación: Noviembre 30 de 2010
RESUMEN
En el presente estudio, llevado a cabo en el Parque Nacional Natural Amacayacu, Amazonia colombiana, se evaluó la variación alométrica entre variables dendrométricas y el crecimiento en dap y biomasa de seis especies clasificadas en tres grupos funcionales, de acuerdo con la densidad de la madera (ρ). Las especies seleccionadas fueron: Eschweilera rufolia, Eschweilera itayensis, Conceveiba guianensis, Otoba parvifolia, Pseudolmedia laevis y Apeiba aspera. El dap fue la variable estructural explicatoria más importante. Cuando se evaluó la relación entre altura total y dap, el parámetro b o coeficiente de escalamiento alométrico varió entre especies, lo que mostró una tendencia al aumento y, por tanto, una reducción en la conicidad, proporcional a la densidad de la madera. No se encontraron diferencias significativas en el incremento diamétrico ni entre especies (P=0.119, F=1.80), ni entre grupos funcionales (P=0.153, F=1.19). Asimismo, el incremento en biomasa entre especies o grupos funcionales tampoco mostró diferencias significativas (P = 0.0784, F = 2.05; P = 0.0711, F = 2.71, respectivamente). No obstante, se encontró una tendencia a un mayor incremento diamétrico en promedio de las especies clasificadas como pioneras tempranas y una clara tendencia al aumento en biomasa en las especies clasificadas como tolerantes, es decir, aquellas con mayor densidad de madera. En este estudio, a excepción de lo encontrado en Otoba parvifolia, el coeficiente de forma de fuste proveniente del modelo H-dap propone formas de fuste más cilíndricas en especies de más lento crecimiento. Se sugiere que el bosque se recupera en biomasa a tasas constantes independientemente de la edad del parche. Este resultado resalta la importancia que tienen las especies pioneras y la formación de claros en los bosques amazónicos de tierra firme sobre la dinámica del carbono y la coexistencia de especies.
Palabras clave: cambio climático, densidad de la madera, grupos funcionales, PNN Amacayacú.
ABSTRACT
In this study carried out in the Amacayacu National Park in the Colombian Amazonia, we assessed the alometric relationship among different tree structural variables and the growth in diameter and biomass of six species classified according to their wood specific gravity (ρ). The tree species chosen were Eschweilera rufolia, Eschweilera itayensis, Conceveiba guianensis, Otoba parvifolia, Pseudolmedia laevis, and Apeiba aspera. The dbh was the most important structural explanatory variable. Regarding the total height – dbh model, the alometric coefficient b changed between species showing a trend to increase, and thus a taper decrease, proportional to ρ. There were no significant differences in diameter growth between species (P=0.119, F=1.80) or functional groups (P=0.153, F= 1.19). Likewise, biomass growth did not show significant differences neither between species (P=0.0784, F=2.05) nor functional groups (P=0.0711, F=2.71). However, there was a positive trend between ρ and diameter growth and a negative one between ρ and biomass growth. The results of this study suggest that this forest is recovering in biomass at a constant rate independent of the patch age, which emphasizes on the importance of pioneer species and gap formation on the carbon dynamics and the species coexistence in Amazonian tierra fi rme forests.
Key words: climate change, wood specific gravity, functional groups, PNN Amacayacú.
RESUMO
O presente estudo foi levado a cabo no Parque Nacional Natural Amacayacu, Amazonas Colombiano, se avaliou a variação alométrica entre as variáveis dendrométricas e o crescimento em dap e biomassa de seis espécies classificadas em três grupos funcionais de acordo com a densidade da madeira. As espécies selecionadas foram: Eschweilera rufifolia, Eschweilera itayensis, Conceveiba guianensis, Otoba parvifolia, Pseudolmedia laevis e Apeiba aspera. O dap foi a variável estrutural explicatória mais importante. Quando se avaliou a relação entre altura total e dap, o parámetro do b o coeficiente de escalamento alométrico variou entre espécies, o que mostrou uma tendencia ao aumento e, portanto, uma redução na conicidade, proporcional a densidade da madeira.Não se encontraram diferenças significativas no incremento diamétrico nem entre espécies (P=0.119, F=1.80), nem entre grupos funcionais (P=0.153, F=1.19). Assim mesmo, o incremento na biomasa entre espécies ou grupos funcionais tampouco mostrou diferenças significativas (P = 0.0784, F = 2.05; P = 0.0711, F = 2.71, respectivamente). Não obstante, se encontrou una tendência a um maior incremento diamétrico em média das espécies classificadas como pioneiras precoces e uma clara tendência ao aumento da biomasa nas espécies classificadas como tolerantes, ou seja, aquelas com maior densidade de madeira. Neste estudo, à exceção do encontrado em Otoba parvifolia, o coeficiente de forma de fuste proveniente do modelo H-dap propõe formas de fuste mais cilíndricas nas espécies de crescimento mais lento. Se sugere que o bosque se recupera em biomassa com taxas constantes independentemente da idade do parche. Este resultado resalta a importância que tem as espécies pioneiras e a formação de claros nos bosques amazônicos de terra firme sobre a dinâmica do carbono e a coexistência de espécies.
Palavras chave: Mudança climática, densidade da Madeira, grupos funcionais, PNN Amacayacú.
INTRODUCCIÓN
Las estrategias de historia de vida de los árboles se definen como el reflejo de las respuestas fisiológicas, demográficas y filogenéticas, a las limitaciones impuestas por factores bióticos y abióticos (Condit et al 1996, Chave 1999). Por tanto, conocer las relaciones entre diferentes componentes estructurales de los árboles es importante para poder entender diferencias adaptativas entre especies (King 1996). Este tipo de relaciones y cambios en las dimensiones entre componentes de un organismo a medida que crece se han estudiado mediante el escalamiento alométrico, que se resume por medio de la función de potencia Y = aXb (Thompson 1917; Huxley 1932). En este modelo, la variable biológica “Y”, depende de la masa del cuerpo “X”, mediante el exponente alométrico “b”; la constante “a” puede variar con la especie y el tipo de organismo (Delgado et al. 2005).
Se ha propuesto que los cambios en tamaño y forma reflejan adaptaciones de las especies a las condiciones ambientales, como respuesta a la presencia de factores de estrés (Niklas 1995; Sposito & Santos 2001). De acuerdo con el estado de desarrollo, las características funcionales y las condiciones ecológicas en las que se encuentran las especies, la constante alométrica puede variar. Por ejemplo, se ha reportado una relación directamente proporcional entre el incremento en la relación altura-diámetro, a medida que aumenta la competencia entre individuos por luz (Niklas 1995).
La densidad de la madera, entendida como su peso en gramos por unidad de volumen, ha mostrado estar fuertemente relacionada con la historia de vida de las especies (King et al 2005) y, por tanto, se considera un buen indicador de las características funcionales de estas (Muller-Landau 2004). Estudios recientes han reportado que la baja densidad de la madera tiene un bajo costo energético, por tanto, se ha encontrado inversamente correlacionada con las tasas de crecimiento de árboles tropicales (Suzuki 1999, Keeling et al. 2008). En contraste, la alta densidad de la madera se piensa que concede ventajas de supervivencia, ya que puede reducir la probabilidad de daños físicos (Zimmerman, et ál., 1994) y aumentar la resistencia a plagas y de enfermedades (Turner 2001). En síntesis, se puede concluir que la densidad de la madera está positivamente asociada con la posición sucesional (Saldarriaga et al. 1988; Swaine & Whitmore 1988).
Entender estas diferencias proporciona herramientas de manejo y conservación de los ecosistemas forestales, ya que nos permite entender mejor la dinámica del crecimiento y la forma como la composición de especies puede influenciar el funcionamiento global de los ecosistemas naturales (Keeling et al. 2008). Por esta razón, en el presente estudio de tipo exploratorio llevado a cabo en el Parque Nacional Natural Amacayacu, Amazonia Colombiana, se estudió el crecimiento y la variación alométrica entre variables estructurales como diámetro a la altura del pecho (dap), altura total (H) y diámetro de copa (dc), de seis especies clasificadas en tres grupos funcionales, de acuerdo con la densidad de la madera (ρ). Las preguntas de investigación formuladas fueron: (1) ¿cambian las relaciones alométricas entre componentes estructurales en especies de diferente grupo funcional, definidos de acuerdo con la densidad de la madera?; (2) ¿existen diferencias en las tasas de crecimiento promedio anual en dap entre especies y grupos funcionales?; (3) ¿cómo son los patrones de crecimiento de la comunidad total en relación con el tamaño de los individuos?; (4) ¿cómo es el incremento anual de la biomasa aérea entre las especies de diferente grupo funcional defi nidos, de acuerdo con la densidad de la madera?
De esta forma, haciendo uso del modelamiento alométrico y la densidad de la madera en la clasificación funcional de especies arbóreas, esperamos poder avanzar en el conocimiento de la estructura y funcionamiento de los bosques amazónicos colombianos. Es parte de nuestro propósito, además, contribuir al conocimiento del efecto del cambio global sobre estos ecosistemas, y más en particular, sobre la dinámica del carbono (Baker et al. 2004, Wright 2005).
MATERIALES Y MÉTODOS
ÁREA DE ESTUDIO
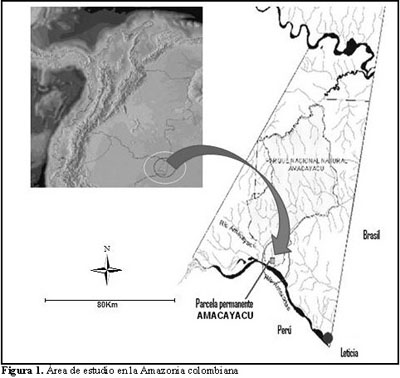
El presente estudio se llevó a cabo dentro de la parcela permanente ubicada en el Parque Nacional Natural (PNN) Amacayacu, localizado en el departamento del Amazonas, extremo sur de la Amazonia colombiana (Figura 1). El PNN Amacayacu hace parte del trapecio amazónico y se encuentra localizado entre 3º 02’ y 3º 47’ de latitud sur y 69º 54’ y los 70º 25’ longitud Oeste, con una extensión de 2930 km2 y elevaciones entre 80 y 100 m (Rudas 1996).
El área del parque reúne sistemas selváticos en ambientes boscosos de planicie aluvial inundable, bosque de tierra firme y algunas zonas pantanosas. En general, en los suelos del área predominan las estructuras que oscilan desde arcillosas, franco-arcillosas hasta franco-arcilloso-limoso y en ocasiones francas, principalmente, en las superficies de denudación de origen sedimentario y en los planos aluviales del río Amazonas (Chamorro 1989). La parcela permanente está ubicada en el plano sedimentario terciario, sobre suelos derivados de la formación Pebas (Hoorn 1994), los cuales se caracterizan por poseer una marcada pobreza en bases intercambiables como resultado del proceso de lavado al que han sido constantemente sometidos. No obstante, estos suelos de origen andino son, generalmente, más fértiles que los encontrados en superfi cies de denudación terciaria, originados a partir del escudo Guyanés (Duivenvoorden & Lips 1995).
De acuerdo con la información disponible en el periodo de 1968-1990, en los registros de la estación climática del aeropuerto Vásquez Cobo de Leticia, la temperatura media anual es de 25,8 ºC, la humedad relativa promedio anual de 86% y la precipitación promedio anual de 3215,9 mm (Prieto, 1994). La precipitación es del tipo unimodal-biestacional, con un periodo de concentración de lluvias entre octubre y mayo; el mayor valor de precipitación se registra en enero, con un promedio de 352,8 mm y el menor está entre junio y septiembre, con un valor de 158,3 mm.
TOMA DE DATOS
Dentro de las primeras 5 ha de la parcela permanente en el PNN Amacayacu, las cuales fueron establecidas en julio del año 2005 y recensadas en mayo del 2007, se mapeó, marcó y midió el diámetro a 1,3 m de altura (DAP) a todos los árboles con 10 cm o más de este. El punto de medición del dap fue marcado con pintura permanente. En estas primeras 5 ha, que poseían identificación botánica de todos los individuos, se seleccionaron seis especies que fueran lo suficientemente abundantes y que representaran los grupos funcionales esperados dentro de toda la comunidad. Las especies seleccionadas fueron: Eschweilera rufolia, Eschweilera itayensis, Conceveiba guianensis, Otoba parvifolia, Pseudolmedia laevis y Apeiba aspera. A cada árbol se le tomó información a las siguientes variables dendrométricas: altura total (H), diámetro a la altura pecho (DAP), diámetro de copa (DC) y la densidad (ρ) de la madera, la cual se obtuvo mediante revisión bibliográfica de estudios realizados en la cuenca Amazónica, dándole prelación a los sitios circundantes al área de estudio.
En árboles con alturas mayores de 15 m, la altura total se midió con un Vertex Md05; en árboles con alturas por debajo de los 15 m, se empleó una Vara Extensible de igual longitud. En cuanto al dap, inicialmente, se realizó la medición de la circunferencia a la altura del pecho (CAP) a una altura de 1.30 m sobre el nivel del suelo (Condit, 1998) con una cinta métrica, para el posterior cálculo del dap. El diámetro de copa se calculó como el promedio de la proyección vertical sobre el suelo del perímetro de la copa en direcciones perpendiculares. Aquellos individuos que se remidieron exactamente en el mismo punto a la altura del DAP se les estimó el crecimiento diamétrico.
Dentro del proceso de estimación de las tasas de crecimiento en dap, se consideraron tres posibles fuentes importantes de error: 1) discrepancias en la remedición de algunos tallos por su complejidad debido a bifurcaciones, raíces fúlcreas o bambas; 2) errores de campo en la lectura o posición de la cinta al momento de hacer la medición del dap; 3) errores de digitación de los datos. Debido a esto, se adoptaron los criterios propuestos por Condit et al. (2004) para la exclusión de datos que pudieran estar asociados con las fuentes de error mencionadas. Estos son: valores mayores a 5 mm*año-1 de crecimiento en dap y valores menores a 2 mm*año-1 en crecimiento negativo. Si se consideran las tres posibles fuentes de error, de los 150 individuos tomados, solo 105 de ellos fueron incluidos en el proceso de análisis del crecimiento diamétrico y crecimiento en biomasa.
ANÁLISIS DE DATOS
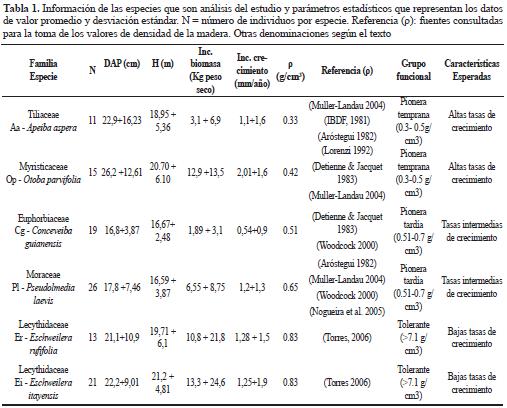
Inicialmente, se hizo una diferenciación de grupos funcionales, de acuerdo con la densidad de la madera (ρ), así: especies pioneras tempranas aquellas con densidades entre 0.3 - 0.5 gr/cm3, pioneras tardías entre 0.51- 0.7 g/cm3 y tolerantes, mayores de 0.71 g/cm3. De acuerdo con la clasificación funcional se plantearon algunos supuestos esperados para cada especie, principalmente en cuanto a las tasas de crecimiento en diámetro (Tabla 1).
Luego, se corrieron modelos alométricos Y = aXb para establecer las relaciones entre las distintas variables estructurales consideradas para cada individuo y especies. Para la obtención de los parámetros, se ajustó la forma linealizada de la ecuación alométrica. Se evaluaron modelos alométricos para la H y dc en función de dap con base en los datos brutos, asumiendo una significancia del 95% (α = 0.05), a fin de comparar y buscar las mejores relaciones entre los diferentes componentes estructurales de los árboles en las diferentes especies. Las diferencias de los modelos alométricos entre los componentes estructurales de las especies con diferente grupo funcional fueron abordadas mediante un análisis descriptivo, con el objetivo de evaluar, de acuerdo con el coeficiente alométrico, la posible variación en la forma de crecimiento de cada individuo.
El cálculo del crecimiento fue definido como el cambio en el dap, dividido por el tiempo en años (Condit et al 2004), expresado por la siguiente ecuación:
Donde A�os = # d�as / 365
Se llevó a cabo un análisis de varianza de una vía (ANAVA), a fin de evaluar si existían diferencias significativas en las tasas de crecimiento promedio anual en dap entre especies y grupos funcionales. De igual manera, en la comunidad (las seis especies juntas), se evaluó mediante un ANAVA la existencia de diferencias estadísticamente significativas entre clases de diámetro. Las clases diamétricas definidas fueron: 10-15, 15,1-20, 20,1-25, 25,1-30, 30,1-35, 35,1- 40 y ≥ 40,1. Dado que se aplicó una transformación logarítmica sobre los datos brutos, para graficar los resultados se le sumó una constante (+ 2) con el objetivo de evitar los valores negativos.
Para el cálculo de la ganancia en biomasa, se estimaron las alturas en t0, con los modelos definidos para cada una de las especies (H vs dap), de manera que se tuviera disponible un intervalo de tiempo de 2 años. El incremento en biomasa (kg en peso seco) de los árboles de las diferentes especies, de acuerdo con el grupo funcional, fue calculado usando una ecuación alométrica que relaciona el dap (cm), ρ (g*cm-3) y H (m), desarrollada por Chave et ál. (2005) para bosques húmedos, como sigue:
Del mismo modo que en el incremento diamétrico, para graficarlo, se le sumó una constante (+400) a cada valor de incremento en biomasa con el objeto de evitar los valores negativos provenientes de la transformación logarítmica. Mediante un análisis de varianza de una vía (ANAVA) se evaluaron las diferencias significativas existentes entre el incremento en biomasa de los diferentes grupos funcionales, definido de acuerdo con la densidad de la madera. Todo el análisis estadístico fue desarrollado en el paquete STATGRAPHICS Centurion XVII.
RESULTADOS
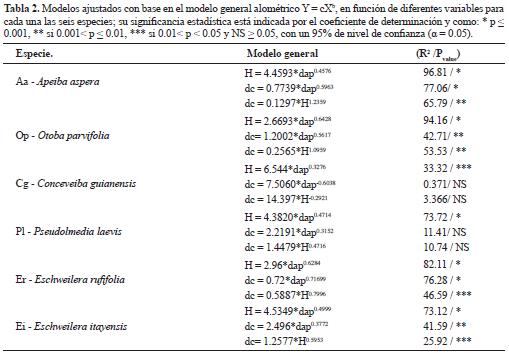
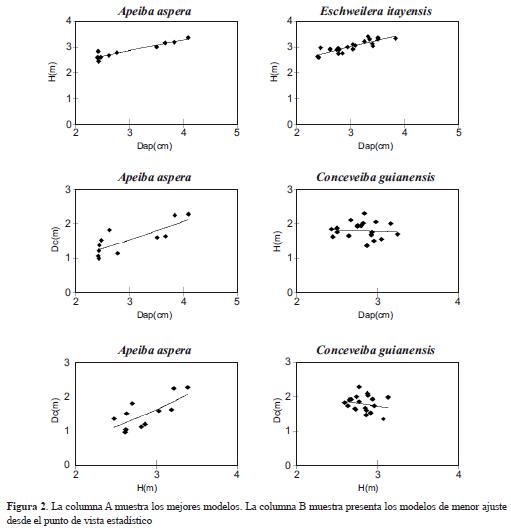
En general, los patrones alométricos entre componentes estructurales de las especies presentaron modelos con buenos ajustes, siendo el dap la variable predictora más importante (Tabla 2). Los modelos ajustados para H en función del dap fueron los más significativos estadísticamente. En las especie Conceveiba guianensis y Pseudolmedia laevis, categorizadas como especies pionera tardías, los modelos alométricos ajustados únicamente fueron significativos, cuando la H estuvo en función del dap; los modelos en función de dc fueron marginalmente o no significativos (Tabla 2, Figura 2). El parámetro b o coeficiente de escalamiento alométrico varió entre especies, mostrando una tendencia al aumento, y por tanto una reducción en la conicidad, proporcional a la densidad de la madera. La especie Otoba parvifolia (b = 0.6428), no obstante, presentó un patrón contrastante (Tabla 2, Figura 2).
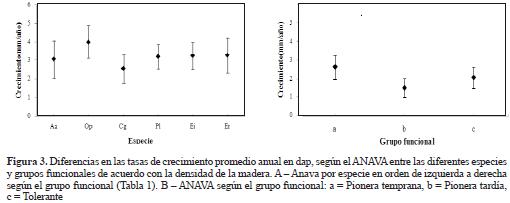
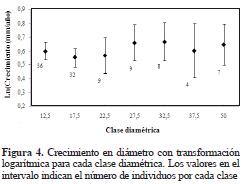
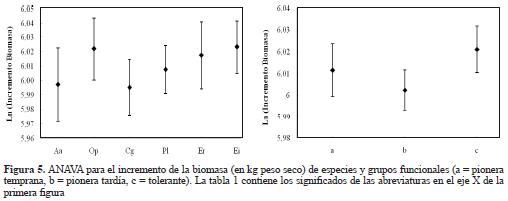
No se encontraron diferencias significativas en el incremento diamétrico ni entre especies (P = 0.119, F = 1.80), ni entre grupos funcionales (P = 0.153, F = 1.19). No obstante, es clara la tendencia a un mayor incremento diamétrico en promedio de las especies clasificadas como pioneras tempranas (Figura 3). En la comunidad, tampoco se detectaron diferencias en crecimiento diamétrico con respecto al tamaño (P = 0.343, F = 1.14); no obstante, y similar al anterior, se evidenció una clara tendencia a un aumento en el crecimiento en diámetro directamente proporcional al tamaño de los individuos (Figura 4). El incremento en biomasa entre especies o grupos funcionales tampoco mostró diferencias significativas (P = 0.0784, F = 2.05; P = 0.0711, F = 2.71, respectivamente). Sin embargo, se evidenció de nuevo una tendencia al aumento en biomasa en las especies clasificadas como tolerantes, es decir, aquellas con mayor densidad de madera (Figura 5).
DISCUSIÓN
COEFICIENTES ALOMÉTRICOS COMO INDICADORES DE GRUPOS FUNCIONALES
El uso de ecuaciones alométricas en los bosques tropicales enfrenta algunas complicaciones, en gran parte, debido a la alta heterogeneidad arquitectural de las especies (Niklas 1994, West et al. 1997). En general, patrones de bifurcación, ramaje y formas de copas irregulares y relativamente complejas dificultan el ajuste de modelos correlativos entre componentes, incluso, en una misma especie. Ese fue el caso particular observado con la especie Conceveiba guianensis, la cual presentó bajos o no significativos ajustes en los modelos alométricos, especialmente, cuando se consideró la copa. Sin embargo, y tal como se corrobora en otros estudios realizados en un amplio rango de bosques tropicales (Ogawa et al. 1965, Brown et al. 1989, Chave et al. 2005), el DAP fue la variable predictora que presentó mejor ajuste en todos los modelos. Esta condición justifica en buena medida el uso generalizado de la relación alométrica H-dap como herramienta en el estudio de la dinámica del crecimiento de los bosques (Saldarriaga et al. 1998), de forma similar a como se hizo en este estudio.
Los resultados referentes a los exponentes alométricos para cada especie muestran una leve tendencia al cambio de la constante alométrica según las características funcionales (Enquist et al. 1998; West et al. 1997). En este estudio, a excepción de lo encontrado en Otoba parvifolia, el coeficiente de forma de fuste proveniente del modelo H-dap para todas las especies mostró una tendencia al incremento de pioneras a tolerantes, lo que propone formas de fuste más cilíndricas en especies de más lento crecimiento. En bosques tropicales de Venezuela, fueron reportadas tendencias más claras, en cuanto a la caracterización alométrica por el grupo funcional (Delgado et al. 2005); no obstante, este último estudio empleó un modelo diferente al alométrico, lo cual difi culta las comparaciones. Adicionalmente, las diferencias en las características ambientales y la escala espacial podrían promover diferencias en la respuesta local de las especies (Patiño et al. 2008).
GRUPOS FUNCIONALES EN RELACIÓN CON EL CRECIMIENTO DIAMÉTRICO Y EL CRECIMIENTO EN BIOMASA
A pesar de que la densidad de la madera ha sido reconocida como determinante de las diferencias en biomasa aérea a lo largo de gradientes sucesionales (Ketterings et al.2001, Nebel et al.2001), los resultados de este estudio no reportan diferencias en los patrones de crecimeinto en dap o biomasa entre grupos funcionales en este bosque amazónico de tierra firme. Un patrón de este tipo sugiere que el bosque se recupera en biomasa a tasas constantes independientemente de la edad del parche. Dado que especies de lento crecimiento como Lecythidaceaes han sido reportadas como las más abundantes dentro de la parcela (Barreto et al. en prensa), este resultado resalta la importancia que tienen las especies pioneras y la formación de claros en los bosques amazónicos de tierra firme sobre la dinámica del carbono y la coexistencia de especies (Laurance et al. 2004). De esta manera, y teniendo en cuenta que la parcela fue establecida inmediatamente después de la sequía del 2005, el hecho de que las especies tolerantes no estén acumulando materia viva a tasas mayores que las pioneras tempranas permite concluir que este bosque podría estar mostrando un leve incremento en biomasa como producto de la recuperación de factores de disturbio. Por lo anterior, es posible pensar que la parcela de Amacayacu no parece haber sido severamente afectada por esta sequía; su clima no estacional y las características de los suelos relativamente fértiles podrían haber favorecido un menor impacto con respecto a la mortalidad de individuos y la liberación de carbono que el reportado en otras regiones amazónicas (Phillips et al. 2009).
Una posible explicación a la falta de significancia en las tendencias observadas está sustentada en el tamaño de la muestra aún bastante pequeño, que promueve el incremento de las varianzas. Esta condición se confirmó, por ejemplo, en el caso del análisis del crecimiento diamétrico que no mostró diferencias significativas con nuestros datos (105 árboles), pero sí cuando se incluyeron todos los individuos de todas las especies colectadas dentro de las primeras 5-ha (1291 individuos), incluyendo las seis especies seleccionadas para el presente estudio; con base en todos los individuos se constató un mayor crecimiento, especialmente, de los individuos con diámetros entre 30 y 40 cm (Arias et al. 2009). De igual forma, aunque las diferencias en crecimiento diamétrico y biomasa no fueron significativas, los resultados de este estudio mostraron tendencias opuestas entre el crecimiento diamétrico y el incremento en biomasa, de acuerdo con la densidad de la madera de cada una de las especies. Resultados similares fueron reportados en un estudio a gran escala en el que se encontró una débil relación entre estas tres variables, pero positivamente influenciada por la fertilidad de los suelos (Keeling et al. 2008). En este sentido, y contrario a lo reportado en este estudio, se esperaría que las especies pioneras tempranas o que demandan mucha luz asignen una gran proporción del carbón fijado a los tejidos en el incremento diamétrico, en comparación con las pioneras tardías y tolerantes que deberían invertir más en la masa foliar (Keeling et al. 2008), aumentando su tasa fotosintética y producción en biomasa.
CONCLUSIONES
Entender el crecimiento de los árboles, en relación con las características funcionales de las especies, es crucial para comprender mejor el impacto de las oscilaciones climáticas sobre la dinámica del carbono en los bosques tropicales (Condit et al. 2004, Phillips et al. 2009), o el posible efecto de eventos de disturbio pasados (Wright, 2005). Los resultados aquí reportados sugieren que la clasificación de pionera tardía fue inoperante y que, simplemente, se comporta de manera similar a las especies tolerantes, cuando se usa la densidad de la madera como variable surrogada de la condición funcional. Posiblemente, mediante el uso de otras características, como por ejemplo el tamaño de los frutos y diásporas, se logre una mejor clasificación de las especies, de acuerdo con sus respuestas fisológicas (Ter Steege et al. 2006, Kraft et al. 2008). Aunque los resultados de este estudio aún se deben considerar prematuros y sobre los cuales se espera depurar con base en los censos y recensos futuros de la parcela de 25-ha (1 cm dap) en todo su conjunto, estos nos permiten resaltar la importancia que tienen los estudios de monitoreo a largo plazo y los tamaños de muestra grandes para poder encontrar mecanismos de adaptación y manejo de los bosques tropicales al cambio global.
AGRADECIMIENTOS
Los autores del presente estudio extendemos nuestro especial agradecimiento a la Dirección de Parques Nacionales Naturales de Colombia y muy en especial a los miembros del PNN Amacayacu. Deseamos agradecer muy en particular a los dos directores de turno de este parque, Jaime Alberto Celis y Alexander Alfonso Segura. A todos los colaboradores de campo y, en general, a toda la comunidad de Palmeras. A un revisor anónimo quien con sus comentarios contribuyó a mejorar el trabajo. El presente estudio fue posible gracias a la financiación recibida de Colciencias (CT 1118- 333-18676) y el Center for Tropical Forest Science (CTFS; subcontrato # 08-440-0000137735).
REFERENCIAS BIBLIOGRÁFICAS
Arias, G. J. C., A. J. Duque & L. D. Cárdenas. 2009. Crecimiento diamétrico de un bosque del Noroccidente amazónico. Revista Colombia Amazónica 2: 57-64.
Aróstegui, A. 1982. Recopilación y análisis de estudios tecnológicos de maderas peruanas. Documento de trabajo No. 2. Proyecto PNUD/ FAO/PER/81/2002 Fortalecimiento de los programas de desarrollo forestal en selva central. Lima. 57 p.
Baker, T.R., O.L. Phillips, S. Almeida, L. Arroyo, A. Difiore, T. J. Killeen, S. G. Laurance, W. F. Laurance, S. L. Lewis, J. Lloyd, A. Monteagudo, D. A. Neill, S. Patiño, N. C. A. Pitman, J. N. M. Silva & R. V. Martínez. 2004. Variation in Wood density determines spatial patterns in Amazonian forest biomass. Global Change Biology 10: 545-562.
Barreto, J. S., A. Duque, D. Cárdenas & F. Moreno.2010. Variación florística a escala local en un bosque de tierra firme en la Amazonia Colombiana. Acta Amazónica 40:179-188
Brown, S., A. Gillespie & A. E. Lugo. 1989. Biomass estimation methods for tropical forests with applications to forest inventory data. Forest Science 35: 881-902.
Condit, R., S. P. Hubbell & R. B. Foster. 1996. Assessing the response of plant functional types to climate change in tropical forest. Journal of Vegetation Science 7: 405-416.
Condit, R. 1998. Tropical Forest census plots. Springer-Verlag and R.G. Landes Company. Berlin, Heidelberg, New York. 211 p.
Condit, R., S. Aguilar, A. Hernandez, R. Perez, S. Lao, G. Angehr, S. P. Hubbell & R. B. Foster, 2004. Tropical Forest dynamics across a rainfall gradient and the impact of the Niño dry season. Journal of tropical Ecology 20: 51-72.
Chamorro, C. 1989. Biología de los suelos del Parque Nacional Natural Amacayacu y zonas adyacentes. Amazonas, Colombia. Revista Colombia Geográfica 15: 45-63.
Chave, J. 1999. Study of structural, successional and spatial patterns in tropical rain forests using TROLL, a spatially explicit forest model. Ecological Modelling 124: 233-254.
Chave, J., C. Andalo, S. Brown, M. A Cairns, J. J. Chambers, D. Eamus, H. Folster, D. F. Fromar, N. Higuchi, T. Kira, J. J. Lescure, B. W. Nelson, H. Ogawa, H. Puig, B. Riera & T. Yamacura. 2005. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Ecosystem Ecology 145: 87-99.
Delgado, L. A., M. F. Acevedo, H. Castellanos, H. Ramirez & J. Errano. 2005. Relaciones alométricas y patrones de crecimiento para especies de árboles de la reserva forestal Imataca, Venezuela. Interciencia 30: 275-283.
Detienne, P. & P. Jacquet. 1983. Atlas d'Identification des Bois de l'Amazonie et des Regions Voisines. Centre Technique Forestier Tropical, Nogent-sur-Marne. France. 640 p.
Duivenvoorden, J. F. & J. M. Lips. 1995. A land-ecological study of soil, vegetation, and plant diversity in Colombian Amazonia. Tropenbos Series 12. The Tropenbos Foundation, Wageningen. 438 p.
Enquist, B. J., J. H. Brown & G.B. West. 1998. Allometric scaling of plant energetics and population density. Nature 395: 163-165.
Fearnside, P. M. 1997. Wood density for estimating forest biomass in Brazilian Amazonia. Forest Ecology and Management 90: 59-87.
Hoorn, C. 1994. An environmental reconstruction of the palaeo-Amazon river system (Middle to late Miocene, NW Amazonia). Palaeogeography, Palaeoclimatology, Palaeoecology 112: 187-238.
Huxley, J. S. 1932. Problems of Relative Growth. MethuenLondres. 276 p.
Instituto Brasileiro de Desenvolvimento Florestal -IBDF-. 1981. Madeiras de Amazônia, caracteristicas e utilização, Vol. 1. Floresta Nacional do Tapajós. Brasilia. 113 p.
Kraft, N. J. B., R. Valencia & D. D. Ackerly. 2008. Functional traits and niche-based tree community assembly in an Amazonian forest. Science 322: 580-582.
Keeling, C. H., R. T. Baker, V. R. Martinez, A. Monteagudo & L. O. Phillips. 2008. Contrasting patterns of diameter and biomass increment across tree functional groups in Amazonian forests. Oecología 158: 521-534.
Ketterings, Q. M., R. Coe & M. Van Noordwijk. 2001. Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests. Forest Ecology and Management 146: 199-209.
King, D. A. 1996. Allometry and life history of tropical trees. Journal Tropical Ecology 12: 25-44.
King, D. A., S. J. Davies, M. N. Supardi & S. Tan. 2005. Tree growth is related to light interception and Wood density in to mixed dipterocarp forests of Malaysia. Functional Ecology 19: 445-453.
Laurance, W.F., A. Alexandre, S. Oliveira, G.W. Laurance, R. Condit, S. Henrique, E. M. Nascimento, A. C. Sanchez-Thorin, T E. Lovejoy, S. Ana C. Andrade, S. Agra, J. E. Ribeiro, L.S. Christopher & W. Dick. 2004. Pervasive alteration of tree communities in undisturbed Amazonian forests. Nature 428: 171-175.
Lorenzi, H. 1992. Árboles brasileiras: Manual de identificação e cultivo de plantas arbóreas nativas do Brasil. Instituto Plantarum de Estudos da Flora Ltda-Nova Odessa. São Pablo.
Muller-Landau, H. C. 2004. Interspecific and Inter-site Variation in Wood Gravity of Tropical Trees. Biotropica 36: 20-32.
Nebel, G., J. Dragsted & A. S. Vegas. 2001. Litter fall, biomass and net primary production in flood plain forest in the Peruvian Amazon. Forest Ecology and Management 150: 93-102.
Niklas, K. J. 1994. Plant allometry: The scaling of form and process. University of Chicago Press. Chicago. 395 p.
Niklas, K. J. 1995. Size-dependent allometry of tree height, diameter and trunk- taper. Annals of Botany 75: 217-227.
Nogueira, E. M., B. W. Nelson & P. M. Fearnside. 2005. Wood density in dense forest in central Amazonia. Brazil. Forest Ecology and Management 208: 261-286.
Ogawa, H., K. Yoda, K. Ogino & T. Kira. 1965. Comparative ecological studies on three main types of forest vegetation in Thailand II Plant biomass. Nature an Life Southeast Asia 4: 49-80.
Patiño, S., J. Lloyd, R. Paiva, C. A. Quesada, L. M. Mercado, T. R. Baker, C. I. Czimczik, M. Schwarz, J. Schmerler, A. Sota, A. Santos, V. Horna, J. Peacock, M. Wagner, L. Arroyo, S. Almeida, E. Alvarez, A. Aguilar, D. Bonal, J. Gallo, R. Herrera, N. Higuchi, E. J. Hoyos, E. M. Jimenez, T. Killeen, E. Leal, F. Luizaño, Y. Malhi, P. Meir, A. Monteagudo, D. Neill, V. P. Nuñez, W. Palomino, A. Peña-Cruz, M.C. Peñuela, O.L. Phillips, N. L. Pitman, F. N. Priante, A. Prieto, S. N. Panfil, A. Rudas, R. Salomao, N. Silva, M. Silveira, A. Lezama, J. D. Turriago, R. Vasquez-Martinez, I. Vieira, B. Villanueva & P. Vitzthum. 2008. Branch xylem density variation across the Amazon Basin. Biogeosciences Discuss 5: 2003-2047.
Phillips, O. L., L. Aragão & S. Lewis. 2009. Drought Sensitivity of the Amazon Rainforest. Science 323: 1344-1347.
Prieto, A. 1994. Análisis estructural florístico y de la vegetación de la isla Mocagua, río Amazonas. Amazonas, Colombia. Tesis. Universidad Nacional de Colombia. Bogotá.
Rudas, L. A. 1996. Estudio florístico de la vegetación del Parque Nacional Natural Amacayacu. Amazonas, Colombia. Tesis de MSc. Universidad Nacional de Colombia. Bogotá.
Saldarriaga, J. G., D. C. West, M. L. Tharp & C. Uhl. 1988. Long-term chronosequence of forest succession in the Upper Rio Negro of Colombia and Venezuela. Journal Ecology 76: 938-958.
Suzuki, E. 1999. Diversity in specific gravity and water content of wood among Bornean tropical rainforest trees. Ecological Research 14: 211-224.
Sposito, T. C. & F. A. M. Santos, 2001. Sacling of stem and crown in eight Cecropias (Cecropiaceae) species of Brazil. American Journal of Botany 88: 939-949.
Swaine, M. D. & T. C. Whitmore. 1988. On the definition of ecological spices in tropical rain forests. Vegetation 75: 81-86.
Ter Steege, H., N. C. A. Pitman, O.L. Phillips, J. Chave, D. Sabatier, A. Duque, J. F. Molino, M. F. Prévost, R. Spichiger, H. Castellanos, P. Von Hildebrand & R. Vásquez. 2006. Continental-scale patterns of canopy tree composition and function across Amazonia. Nature 443: 444-447.
Thompson, D. W. 1917. On Growth and Form. Cambridge University Press. Cambridge.793 p.
Torres, A. J. 2006. Densidad de madera en cuatro tipos de bosque localizados sobre suelos contrastantes en la región de Leticia, Amazonia Colombiana. Departamento de ciencias biológicas, Facultad de Ciencias, Universidad de los Andes. Bogotá. 55 p.
Turner, I. M. 2001. The ecology of trees in the tropical rain forest. Cambridge University Press. Cambridge. 290 p.
West, G.B., J. H. Brown & B. J. Enquist. 1997. A general model for the origin of allometric scaling laws in biology. Science 276: 122-126.
Woodcock, D. W. 2000. Wood specific gravity of trees and forest types in the Southern Peruvian Amazon. Acta Amazónica 30: 589-599.
Wright, S. J. 2005. Tropical forest in a changing environment. Trends in Ecology Evolution 20: 553-560.
Zimmerman, J. K., E. M. Everham II, R. B. Waide, D. J. Lodge, C. M. Taylor & N. V. L. Brokaw, 1994. Responses of tree species to hurricane winds in subtropical wet forest in Puerto Rico: Implications for tropical tree life histories. Journal Ecology 82: 911-922.




License
Colombia Forestal retains the patrimonial rights (copyright) of the published works, and favors and allows the reuse of the same under the Creative Commons Attribution-ShareAlike 4.0 International license, so they can be copied, used, disseminated, transmitted and exhibited publicly, provided that:
You acknowledge the credits of the work in the manner specified by the author or licensor (but not in a way that suggests that you have their support or that they endorse your use of their work).