
DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2012.1.a04Publicado:
01-01-2012Número:
Vol. 15 Núm. 1 (2012): Enero-JunioSección:
Artículos de investigación científica y tecnológicaPATRONES DE ARQUITECTURA FOLIAR ASOCIADOS AL CRECIMIENTO FUNCIONAL DE CINCO ESPECIES LEÑOSAS NATIVAS DE LA CORDILLERA ORIENTAL UTILIZADAS EN RESTAURACIÓN ECOLÓGICA EN LA SABANA DE BOGOTÁ
Foliar architecture patterns of five native woody species utilized for ecological restoration in Bogotá
Palabras clave:
woody shrub, leaf area, forest biomass, global climate change, Andean plant ecophysiology, leaf diversity, canopy. (en).Palabras clave:
arbustos leñosos, área foliar, biomasa forestal, cambio climático global, ecofisiología de plantas andinas, diversidad foliar, dosel. (es).Descargas
Referencias
Ahuja, L.R., Reddy, V.R., Saseendran, S.A., & Quiang, Y. (2008). Response of crops to limited water: Understanding and modeling water stress effects on plant growth processes. Madison: American Society of Agronomy Inc. 436 p.
Begon, M., Townsend, C.R., & Harper, J.L. (2006). Ecology: From Individuals to Ecosystems (4th ed.). Melden-Oxford-Carlton: Blackwell Publishing Ltd. 738 p.
Blaser, J., Sarre, A., Poore, D., & Johnson, S. (2011). Status of Tropical Forest Management 2011 (Technical Series No. 38). Yokohama: International Tropical Timber Organization. 28 p.
Brantley, S.T., & Young, D.R. (2007). Leaf-area index and light attenuation in rapidly expanding shrub thickets. Ecology, 88(2), 524-530.
Castellanos, N.M. (2001). Programa piloto de adaptación de especies nativas para la restauración de ecosistemas de bosque andino de la cordillera oriental. Parque ecológico La Poma (Tesis inédita de pregrado). Ibagué: Universidad del Tolima. 115 p.
Cournac, L., Duboist, M., Chavet, J., & Riera, B. (2002). Fast determination of light availability and leaf area index in tropical forests. Journal of Tropical Ecology, 18, 295-302.
Fageria, N.K., Baligar, V.C., & Clarck, R.B. (eds.). (2006). Physiology of crop production. London: Routledge. 345 p.
Farmer, R.E. (2006). Comparative analysis of first year growth in six deciduous trees. Canadian Journal of Forestry Restoration, 36, 35-41.
Gajardo-Caviedez, P.A., Espinosa, M.A., González, U., & Ríos, D.G. (2005). The influence of thinning and tree size on the sapwood area/leaf area ratio in coigue. Canadian Journal Forest Restoration, 35, 1679-1685.
Hay, R., & Porter, J. (2006). The physiology of crop yield (2nd ed.). Oxford: Blackwell Publishing. 313 p.
Hopkings, W.G. (2006). Photosynthesis and respiration. New York: Chelsea House publishers. 169 p.
Hoover, C.H. (2008). Field measurements for forest carbon monitoring: A landscape-scale approach. Berlin: Springer. 236 p.
Landsberg, J.J., & Gower, S.T. (1997). Applications of physiological ecology to forest management. New York: Academic Press. 354 p.
Larcher, W. (2003). Physiology plant ecology: Ecophysiology and stress physiology of functional groups (4th ed.). Berlín: Springer. 513 p.
Lowman, M.D., & Rinker, H.B. (2004). Forest canopies (2nd ed.). Oxford: Elsevier Academic Press. 517 p.
Lüttge, U. (2008). Physiological ecology of tropical plants (2nd ed.). Berlin: Springer 458 p.
Lüttge, U., Beck, E., & Bartels, D. (2011). Plant desiccation tolerance: Analysis and synthesis, Vol. 215, Ecological studies. Berlin: Springer. 386 p.
McGarvey, R.C., Martin, T.A., & White, T.L. (2004). Integrating within-crown variation in net photosynthesis in loblolly and slash pine families. Tree Physiology, 24, 1209-1220.
Morison, J.I.L., & Morecroft, M.D. (2006). Plant growth and climate change. Oxford: Blackwell Publishing. 213 p.
Nouvellon, Y., Laclau, J.P., Epron, D., Kinana, A., Mabiala, A., Roupsard, O., Bonnefond, J.M., le Maire, G., Marsden, C., Bontemps, J.D., & Saint-Andre, L. (2010). Within-stand and seasonal variations of specific leaf area in a clonal Eucalyptus plantation in the Republic of Congo. Forest Ecology and Management, 259, 1796-1807.
Pallardy, S.G. (2008). Physiology of woody plants (3rd ed.). San Diego: Academic Press, Elsevier. 454 p.
Pessarakli, M. (2002). Handbook of plant and crop physiology (2nd ed.). New York: Marcel Decker Inc. 973 p.
Pretzsch, H. (2009). Forest dynamics, growth and yield. From measurement to model. Berlin: Springer. 664 p.
Ravindranth, N.H., & Oswald, M. (2008). Carbon inventory methods: Handbook for greenhouse gas inventory, carbon mitigation and roundwood production projects. Berlin: Springer. 315 p.
Riaño N.M., Tangarife, G., Osorio, O.I., Girald, J.F., Ospina, C.M., Obando, D., Gómez, L., & Jaramillo, L.F. (2005). Modelo de crecimiento y captura de carbono para especies forestales en el trópico. Bogotá: Cenicafé - Ministerio de Agricultura y Desarrollo Rural - Conif - Federación Nacional de Cafeteros de Colombia. 51 p.
Rodríguez, N. (2010). Determinación del carbono capturado por cinco especies arbóreas y arbustivas del bosque seco altoandino en el Parque Ecológico la Poma (Tesis inédita de pregrado). Ibagué: Universidad del Tolima. 85 p.
Sack, L., & Frole, K. (2006). Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees. Ecology, 87, 483-491.
Sack, L., & Holbrook, N.M. (2006). Leaf hydraulics. Annual Review of Plant Biology, 57, 361-381.
Salisbury, F.B., & Ross, C.W. (1992). Plant physiology (4th ed.). Belmont: Wadsworth Publishing Company. 682 p.
Sokal, R.R., & Rohlf, F.J. (2009). Introduction to biostatistics. New York: John Wiley and Sons, Inc. 768 p.
Taiz, L., & Zeiger, E. (2006). Plant physiology (4th ed.). New York: Sinauer Associates. 690 p.
Terashima, I., & Hikosaka, K. (2005). Comparative ecophysiology of leaf and canopy photosynthesis. Plant Cellular Environment, 18, 1111-1128.
Wareing, P.F., & Phillips, I.D.J. (2006). Growth and differentiation in plants (4th ed.). Oxford: Pergamon Press. 343 p.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
 | Melo-Cruz, O., Rodríguez-Santos, N., & Rojas-Ramírez, F. (2012). Patrones de arquitectura foliar asociados al crecimiento funcianal de cinco especies leñosas nativas de la cordillera oriental utilizadas en restauración ecológica en la sabana de Bogotá. Colombia Forestal, 15(1), 119-130. |
PATRONES DE ARQUITECTURA FOLIAR ASOCIADOS AL CRECIMIENTO FUNCIONAL DE CINCO ESPECIES LEÑOSAS NATIVAS DE LA CORDILLERA ORIENTAL UTILIZADAS EN RESTAURACIÓN ECOLÓGICA EN LA SABANA DE BOGOTÁ
Foliar architecture patterns of five native woody species utilized for ecological restoration in Bogotá
Padrões de arquitetura foliar associados ao crescimento funcional de cinco espécies lenhosas nativas da Cordilheira Oriental utilizadas em restauração ecológica na savana de Bogotá
Omar Melo-Cruz1, Nathaly Rodríguez-Santos2 & Ferney Rojas-Ramírez3
1Grupo de investigación en Biodiversidad y Dinámica de Ecosistemas Tropicales, Universidad del Tolima, Ibagué. Grupo de investigación en Ecofisiología Agraria, Universidad Nacional de Colombia, Sede Medellín. omelo@ut.edu.co
2Grupo de investigación en Biodiversidad y Dinámica de Ecosistemas Tropicales, Facultad de Ingeniería Forestal, Universidad del Tolima. Cra 11B 4C No17-19, Barrio San Diego, Ibagué (Tolima). natha8902@hotmail.com. Autor para correspondencia.
3Cámara de Comercio de Bogotá, Corporación Ambiental Empresarial - Programa hojas verdes. dtecnicophojasverdes@ccb.org.co
Recepción. Julio 6 de 2011 / Aprobación. Abril 26 2012
RESUMEN
Esta investigación se realizó en el Parque Ecológico la Poma que hace parte de los paisajes de la Sabana de Bogotá y corresponde a un bosque seco Montano Bajo (bs-Mb), con presencia recurrente de heladas, lo que genera un ambiente crítico para el crecimiento de la vegetación. Se evaluó y se caracterizó el componente foliar, los patrones que definen la arquitectura de dosel y los parámetros de crecimiento funcional, para poblaciones de las especies Abatia parviflora (Ap), Dodonaea viscosa (Dv), Escallonia paniculata (Ep), Baccharis macrantha (Bm)y Quercus humboldtii (Qh), con edades entre uno y diez años, bajo la hipótesis que plantea la existencia de tendencias particulares de crecimiento asociadas a un patrón arquitectural específico. Se evaluaron variables de tipo morfométrico foliar, variables para la caracterización de la arquitectura de dosel y variables de crecimiento funcional para las especies mencionadas. Para cada variable se realizó un análisis de varianza (p < 0.05) y en el test de rango múltiple se utilizó la prueba Tukey para la separación de las medias. Como resultado se encontró que las especies con mayor eficiencia fotosintética, y por consiguiente, con mayor capacidad de acumulación de materia seca expresada en la biomasa fueron Qh y Dv, las cuales presentaron un dosel de tipo erectófilo que generó la mayor absorción lumínica. Las demás especies (Ap, Ep y Bm) tienen doseles planófilos y plagiófilos que se pueden ligar a una menor capacidad productiva. Este trabajo vislumbra el potencial de las especies leñosas arbustivas para ser utilizadas en ambientes donde los arboles tienen restringido su crecimiento.
Palabras clave: arbustos leñosos, área foliar, biomasa forestal, cambio climático global, ecofisiología de plantas andinas, diversidad foliar, dosel.
ABSTRACT
This research was carried out in La Poma Ecological Park, which is located on the Sabana de Bogotá and corresponds to an area of Lower Montane dry forest. Frequent incidences of frost means that it is a challenging environment for vegetation growth. Leaf components, patterns of canopy architecture and functional growth parameters were evaluated and characterized for populations of Abatia parviflora (Ap), Dodonaea viscosa (Dv), Escallonia paniculata(Ep), Baccharis macrantha (Bm) and Quercus humboldtii (Qh) that ranged in age from one to ten years. We hypothesized that particular growth tendencies were associated with a specific architectural pattern. Morphometric leaf, canopy architecture and functional growth variables were evaluated for these species. An Anova (P <0.05) was performed for each variable along with a multiple range Tukey test that separated the means. We found that the species with higher photosynthetic efficiency and higher dry matter accumulation capacity expressed in biomass were Qh and Dv. These species have an erectophil canopy with the highest light absorption. The other species (Ap, Ep, and Bm) have planophil and plagiophil canopies that have a lower productive capacity. This work provides information on the potential use of woody shrub species in environments where trees have restricted growth.
Key words: woody shrub, leaf area, forest biomass, global climate change, Andean plant ecophysiology, leaf diversity, canopy.
RESUMO
Esta pesquisa foi realizada no Parque Ecológico La Poma que faz parte do cenário da Savana de Bogotá e corresponde a um bosque seco Montano Bajo (bs-Mb), com presença recorrente de geada, resultando em um ambiente crítico para o crescimento da vegetação. Avaliou-se e caracterizou-se o componente da folha, os padrões que definem a arquitetura da copa e os parâmetros de crescimento funcional para as populações de espécies Abatia parviflora (Ap), Dodonaea viscosa (Dv), Escallonia paniculata (Ep), Baccharis macrantha (Bm) e Quercus humboldtii (Qh), com idades entre um a dez anos, sob a hipótese que supõe a existência de tendências particulares de crescimento associadas a um padrão de arquitetura específico. Foram avaliadas as variáveis de tipo morfométrico foliar, para a caracterização da arquitetura da copa, e variáveis de crescimento funcional para as espécies mencionadas. Para cada variável realizou-se uma análise da variância (p < 0.05) e no teste de atuação múltipla, utilizou-se a prova Tukey para a separação das médias. Como resultado verificou-se que as espécies com maior eficiência fotossintética e, portanto com maior capacidade de acumulação de matéria seca expressada na biomassa foram Qh e Dv quem apresentou um dossel do tipo erectófilo que gerou maior absorção de luz. As demais espécies (Ap, Ep e Bm) têm copa, planófilos e plagiófilos que podem estar vinculadas a uma menor capacidade produtiva. Este trabalho vê o potencial de espécies arbustivas lenhosas para utilização em ambientes onde as árvores têm restringido seu crescimento.
Palavras chave: arbustos lenhosos, área foliar, biomassa floresta, mudança climática global, ecofisiologia das plantas andinas, diversidade de folhagem, dossel ou copa.
INTRODUCCIÓN
El conocimiento de los patrones de la arquitectura foliar de las plantas leñosas es de gran importancia para entender el efecto que tiene la disposición y el arreglo del follaje en el proceso de absorción, transmisión y reflexión de la energía radiante, lo que permite la generación de hipótesis sobre los procesos ecofisiológicos relacionados con la captura de carbono y la distribución diferencial de los fotoasimilados, de acuerdo con el entorno ambiental de crecimiento (Fageria et al., 2006). Asimismo, evaluar la arquitectura foliar permite identificar mecanismos funcionales que vinculan la expansión foliar a factores ambientales, como la disponibilidad de luz, temperatura y agua (Pessarakli, 2002), en especial, en entornos ambientales con restricciones hídricas (Ahuja et al., 2008). Igualmente, determinar el área foliar de las plantas tiene gran importancia en los estudios relacionados con su crecimiento y desarrollo, dado que en las hojas se sintetizan los carbohidratos que van a repartirse en los diferentes órganos, lo que permite efectuar la estimación de la cobertura foliar y el índice de área foliar (IAF), que son parámetros básicos para estudios ecofisiológicos y de bioproductividad de las plantas (Larcher, 2003; Hay & Porter, 2006).
Las hojas de las plantas leñosas varían enormemente tanto en estructura como en composición, en consecuencia, se generan diferencias funcionales que permiten la coexistencia entre las diferentes especies (Hopkings, 2006).Las hojas de los árboles varían enormemente en su estructura y composición, con lo cual contribuyen a la generación de diferencias funcionales que conducen la coexistencia de las especies. La diversidad en los rasgos de las hojas surge de la filogenia y la adaptación, la cual es limitada por la correlación entre los rasgos. Por ejemplo, los rasgos de la hoja relacionados con la economía del carbono están estrechamente coordinados dentro y por medio de los biomas, de manera que la masa foliar por unidad de área, está positivamente correlacionada con el lapso de vida de la hoja y negativamente relacionada con las tasas de asimilación de nitrógeno y fotosintética por unidad de masa foliar, lo mismo que con características hidráulicas de la hoja de gran importancia para la funcionalidad de la planta (Landsberg & Gower, 1997; Sack & Holbrook, 2006; Ahuja et al., 2008).
En general, para las coberturas boscosas heterogéneas y estratificadas, la absorción de la luz por el dosel del bosque se realiza en forma diferencial, de manera que cerca del 80% de esta, se captura en las copas de los árboles superiores, donde se absorben longitudes de onda principalmente de color azul. En el interior del bosque, entre el sub-dosel y el sotobosque, se absorbe alrededor del 9% de la luz, en longitudes de onda del color amarillo y naranja. Cerca de la superficie del suelo se absorbe tan solo un 2% de la radiación total, lo cual se expresa particularmente en longitudes de onda del rojo y el rojo lejano. El color verde y algunas fracciones del amarillo son reflejados hacia la atmósfera en gran parte por la clorofila de las hojas, en consecuencia, alcanza valores cercados al 10% de la radiación incidente (Begon et al., 2006; Taiz & Zeiger, 2006; Pallardy, 2008). Igualmente, en la planta individual, la forma como incide la radiación y como esta es absorbida es diferencial. Las hojas de la periferia de la copa del árbol tienen la posibilidad de absorber mayor cantidad de luz, mientras que las hojas ubicadas en el interior de la copa solo reciben una baja cantidad de radiación; por esta condición, las plantas presentan heterogeneidad en sus tipos de hojas, desarrollando tanto hojas de luz como de sombra, dentro de la copa del mismo individuo. De igual forma, el arreglo angular de las hojas en la arquitectura de la copa permite la absorción, la transmisión y la reflexión de la radiación, lo que determina su eficiencia fotosintética (Salisbury & Ross, 1992; Sack & Frole, 2006).
Independientemente del patrón de asignación de carbono fijado en la fotosíntesis a los diferentes órganos (hojas, tallos y raíces), el crecimiento y el funcionamiento de la planta depende sobre todo de la adquisición de carbono. Tanto la síntesis como la ganancia de carbono en toda la planta dependen de una serie de propiedades del individuo como un todo, entre ellas la disposición de las hojas, el área foliar total, la dinámica foliar y la forma como se encuentra repartida la biomasa en la planta. Las tasas fotosintéticas también dependen de la relación que exista entre las propiedades de la hoja, las propiedades de la planta y el entorno ambiental de crecimiento (Fageria et al., 2006; Wareing & Phillips, 2006).
Las especies leñosas y en especial los árboles han sido utilizados como elementos mitigadores del cambio climático global por la captura del CO2 atmosférico que ellos realizan. Para disminuir las concentraciones de este gas de efecto invernadero, que en los últimos cien años se ha incrementado de 280 a 375 partes por millón, se han aumentado las áreas de plantación, puesto que los árboles forestales son los que más tiempo retienen el carbono capturado (Morison & Morecroft, 2006). Esta actividad se ha generalizado en ambientes óptimos para su desarrollo, donde los individuos pueden maximizar sus tasas de crecimiento y de acumulación de biomasa (Blaser et al., 2011). Sin embargo, cuando se establecen coberturas vegetales en paisajes con restricciones ambientales, con factores limitantes referidos a la disponibilidad hídrica, la luz o la temperatura, tal como sucede en la mayoría de las regiones secas del territorio colombiano, gran parte de las especies de árboles colapsan y sus poblaciones disminuyen significativamente tanto la supervivencia como la productividad esperada, como consecuencia de la no adaptación al entorno de crecimiento (Lüttge et al., 2011).
Por lo anterior se hace necesario experimentar con otros grupos funcionales de plantas, como las especies arbustivas leñosas, que han mostrado mayor posibilidad de adaptación a ambientes adversos en comparación con muchas especies de árboles forestales. Además, estas especies son capturadoras de carbono y generadoras de otros servicios ambientales (Rodríguez, 2010). El conocimiento de las especies arbustivas en cuanto a su funcionamiento y posibles patrones morfológicos que estas generan para interactuar con el entorno ambiental, se convierten en prioritarios para cubrir los vacíos de información que actualmente existe sobre este grupo de plantas.
El presente trabajo se desarrolló con el propósito de contribuir a la identificación de herramientas y oportunidades de uso, manejo y conservación para un grupo de cinco especies leñosas arbustivas de la cordillera oriental. Por lo general, estas especies vienen siendo utilizadas en la restauración de coberturas degradadas y en entornos ambientales críticos, que presentan restricciones para el crecimiento normal de los árboles forestales. El objetivo principal se enfoca en determinar los patrones que describen la arquitectura del dosel, asociados al crecimiento funcional de las especies Abatia parviflora (Ruiz & Pav), Dodonaea viscosa (L.) Jacq, Escallonia paniculata (Ruiz & Pav), Baccharis macrantha (Kunth) y Quercus humboldtii (Bonpl), como indicadores de la productividad y adaptación de estas a climas adversos y su papel potencial como capturadoras de carbono en un clima cambiante.
MATERIALES Y MÉTODOS
ÁREA DE ESTUDIO
Esta investigación se realizó en el Parque Ecológico la Poma (PEP), ubicado en el sur-oriente de la Sabana de Bogotá, en el municipio de Soacha y con gran influencia del municipio de Sibaté. El PEP cuenta con un área de 140 ha, en la que se han realizado trabajos de restauración ecológica a partir de plantaciones mixtas de especies nativas tanto arbóreas como arbustivas, que se han adaptado exitosamente a este entorno ambiental (Rodríguez, 2010). El área de estudio se encuentra entre los 2600 y 2800 m de altitud, con una precipitación media de 568.4 mm al año; la temperatura promedio alcanza los 12.8°C, la humedad relativa es inferior al 75%, el brillo solar promedio diario es de 1.8 horas de sol, la evapotranspiración potencial se estima en 949 mm anuales, lo que genera un balance hídrico deficitario en más de 490 mm anuales. El climadiagrama tiene un comportamiento bimodal con presencia de heladas en los dos periodos de estiaje. El área de estudio se clasifica dentro de la formación vegetal bosque seco Montano Bajo (bs-MB; Castellanos, 2001).
MUESTREOS
Las especies objeto de trabajo para esta investigación corresponden a cuatro especies leñosas arbustivas (A. parviflora, D. viscosa, E. paniculata, B. macrantha) y una especie arbórea (Q. humboldtii), que se encuentran establecidas en rodales mixtos. Para cada especie se seleccionó una muestra de cien individuos distribuidos uniformemente en cinco categorías de edad comprendidas entre uno y diez años, a los que se les registró la variable dasométrica altura total utilizando un hipsómetro ACE-laser. Posteriormente, utilizando la metodología propuesta por Hoover (2008), se cuantificó la biomasa expresada en peso de materia seca para el individuo promedio por categoría de edad y por componente funcional (hojas, ramas, fuste, raíces). Estos valores se utilizaron para determinar los índices de crecimiento funcional y la generación de los patrones arquitecturales para el componente foliar (Fageria et al., 2006).
Morfometría foliarDe acuerdo con la metodología propuesta por Pallardy (2008), la copa de los árboles seleccionados para cosecha por categoría de edad y por especie se dividió en tres tercios o estratos: apical (1/3), medio (2/3) y basal (3/3); para cada uno de estos se cosecharon uniformemente muestras de 500 g de hojas, a las que se les determinó el área foliar (A), perímetro foliar (P) y el factor de forma de las hojas (SF), utilizando tecnología Li-Cor (Lowman & Rinker, 2004). El área foliar total (AFT) y por estrato en la copa se determinó por relación directa entre el área foliar de las muestras y el peso fresco total de todo el componente foliar del individuo (Ravindranth & Oswald, 2008).
Se organizaron arreglos factoriales tomando como referencia las especies, las clases de edad y cinco repeticiones para una muestra total evaluada de 125 árboles. Las variables se sometieron a pruebas para determinar su normalidad y homogeneidad de varianzas. El efecto de los tratamientos sobre la magnitud y distribución de cada una de las variables se determinó mediante análisis de varianza (p < 0.05). La separación de medias se realizó utilizando la prueba de Tukey (Sokal & Rohlf, 2009) y se generaron modelos de regresión lineal a partir del área foliar y la edad de los individuos, al igual que histogramas para visualizar el efecto del tiempo en la organización del dosel.
Arquitectura de dosel
Las clases de inserción foliar se evaluaron teniendo en cuenta los arreglos planófilos (0° - 30°), erectófilos (60° - 90°) y plagiófilos (30° - 60°); para esto se realizaron lecturas angulares de las ramas sobre el fuste y de los peciolos de las hojas sobre las ramas, cuyas frecuencias fueron graficadas en histogramas de valores relativos (Nouvellon et al., 2010; Riaño et al., 2005; McGarvey et al., 2004). Para determinar el coeficiente de extinción lumínica (k), se realizaron lecturas para cada uno de los estratos de copa, por clase de edad y por especie, con un analizador de frondas LAI-2200 - Plant Canopy Analyzer (Cournac et al., 2002). El coeficiente k se determinó aplicando la ley de Beer y Lambert, que relaciona la intensidad de luz entrante en un medio con la intensidad saliente después de que en dicho medio se produzca absorción. La relación entre ambas intensidades se puede expresar mediante la siguiente relación (Ec. 1; Brantley & Young, 2007):
Donde:
Io: es el valor de la radiación fotosintéticamente activa (RFA) sobre el dosel del árbol.
Iz: es el valor de RFA bajo el dosel o en cualquier estrato dentro de este.
k: [Ln(Io)-Ln(Iz)]/IAF.
IAF: índice de área foliar.
e: es la base de los logaritmos neperianos.
Crecimiento funcional
El crecimiento funcional de las especies se determinó con los siguientes parámetros propuestos por Lüttge (2008):
Tasa de crecimiento de la especie (TCE; Ec. 2): expresado en kg/ha/año.
TCE = (P2 - P1) / A (t2 - t1) Ec. 2
Donde:
A = área donde la materia seca de la especie fue registrada.
P1 = peso de la materia seca cosechada para los individuos de la clase de edad 1.
P2 = peso de la materia seca cosechada para los individuos de la clase de edad 2.
t1 = valor en años de la clase de edad 1.
t2 = valor en años de la clase de edad 2.
Tasa de asimilación neta (TAN; Ec. 3): expresada como gramos de materia seca por m2 por día.
TAN = (PS2 - PS1)(Ln AF2 - Loge AF1) / (AF2 - AF1)(t2 - t1) Ec. 3
Donde:
Ln = logaritmo natural.
PS = peso seco de las muestras en t1 y t2.
AF = área foliar en el periodo t1 y t2.
Relación de Área Foliar (RAF; Ec. 4): expresada en cm2 por gramo de materia seca.
Donde:
PS = peso seco total.
AF = área foliar de la planta.
Área Foliar Específica (AFE; Ec. 5): expresada en cm2 por gramo.
Donde:
AF: área Foliar.
PSAF= peso seco del área foliar.
Índice de Área Foliar (IAF; Ec. 6): es el área foliar por unidad de superficie de suelo.
Donde:
AFT = área foliar total.
S = área de suelo ocupada.
RESULTADOS
MORFOMETRÍA FOLIAR
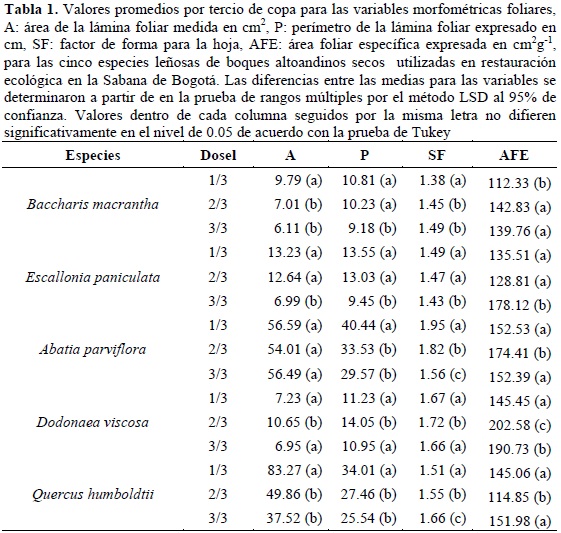
La tabla 1 muestra los valores promedios para las variables morfométricas foliares A, P y SF, las cuales se evaluaron para cada tercio de copa por medio de un análisis de varianza (p < 0.05). En cuanto a la morfometría de la lámina foliar, todas las especies presentan diferencias estadísticamente significativas al 95%, de acuerdo con la posición de las hojas en el dosel. En general, las hojas de mayor tamaño (A y P) se ubican en el primer tercio de copa que corresponde a la sección apical. Este patrón difiere en Dv, donde dicha situación se presenta en la parte media de la copa. Para el caso del SF, las hojas de todas las especies en general presentan hojas más redondeadas hacia el ápice de la copa y más rectangulares hacia la base de estas, a excepción de Dv; cuyas hojas más redondeadas se ubican en la parte media de la copa. Bm y Ep, se caracterizan por generar hojas oval redondas, mientras que Ap, se caracteriza por hojas rectangular oblongas. Dv y Qh, generan hojas del tipo oval oblongo.
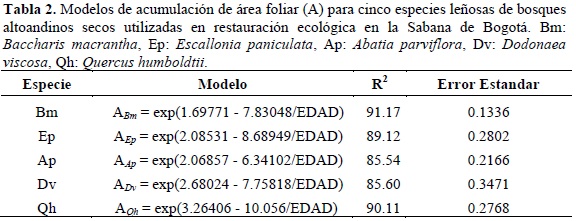
En la tabla 2 se muestran los modelos de acumulación de área foliar para las cinco especies objeto de estudio. El modelo utilizado como base es de tipo sigmoideo, que expresa la tendencia biológica de crecimiento para los organismos vivos. En general todos los modelos muestran altos valores de ajuste con R-cuadrado superior al 85%. Los datos más homogéneos se presentaron en Bm con un error estándar de 0.1336, mientras que Dvmostró mayor heterogeneidad con un error estándar de 0.3471.
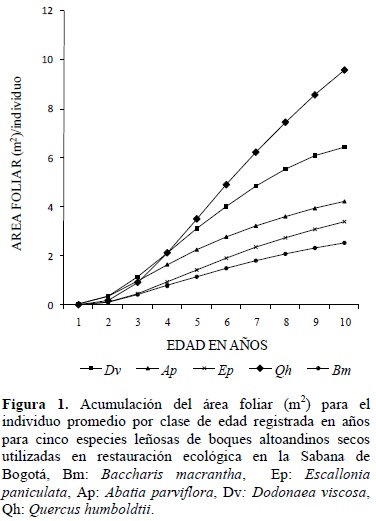
La tendencia de los modelos de acumulación de área foliar se visualizan en la figura 1, para los cuales la proyección de crecimiento es de diez años. La gráfica muestra el área foliar expresada en m2, acumulada para el individuo promedio de las respectivas clases de edad. A los diez años de edad la especie que presenta mayor acumulación de área foliar es Qh, mientras que las especies Bm, Ap y Ap, en conjunto presentan los menores valores. Dv presenta un comportamiento intermedio.
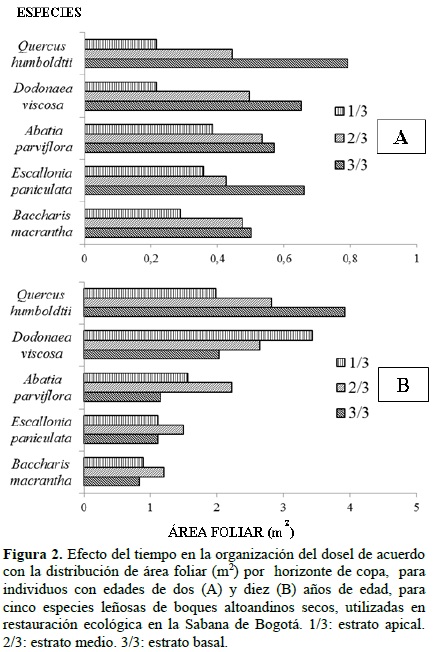
En la figura 2 se muestra el efecto del tiempo en la forma como de distribuye el área foliar de acuerdo con la estratificación del dosel. Para lo cual se compara la organización de la copa en estados tempranos de desarrollo a los dos años de edad, frente a individuos de diez años. La copa de todas las especies en estados tempranos tiene una distribución piramidal del área foliar. A los diez años de edad solamente Qh conserva esta distribución. Para Bm y Ep, las copas de sus individuos a los 10 años de edad presentan tanto en el estrato apical como el basal aproximadamente la misma área foliar, que es inferior al área foliar presentada por el estrato medio de la copa. En la copa de Ap, el estrato media presenta la mayor área foliar y el estrato apical es considerablemente menor al estrato basal. En Dvla copa tiene una forma invertida frente al dosel de los estados tempranos, con los mayores valores de área foliar en el estrato apical y el menor valor en el estrato basal.
ARQUITECTURA DE DOSEL
En la figura 3 se muestra el comportamiento de la frecuencia de inserción foliar para las especies objeto de estudio. Las especies Qh y Dv, presentan un dosel dominado en su mayoría por inserciones angulares de tipo erectófico, cuya frecuencia supera el 80%. Las especies Ep y Bm, presentan un dosel de tipo planófilo, con una frecuencia de inserción foliar alrededor del 50%. La especie Ap, es plagiófila en un 60%.

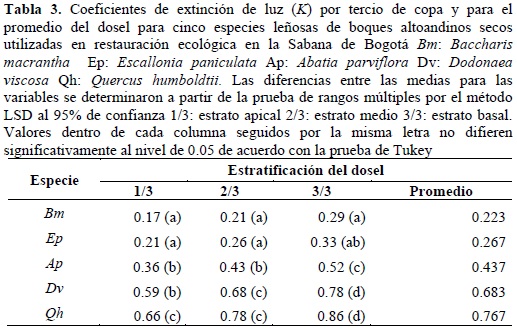
El comportamiento del coeficiente de extinción de luz (k) por tercio de copa y para el promedio del dosel, se muestra en la tabla 3. En general el análisis de varianza (p < 0.05) mostró diferencias estadísticamente significativas tanto para las especies como para la estratificación del dosel, a excepción de Ep que se comporta homogéneamente con Bm. No se presentan diferencias significativas en el valor de k, con el factor edad expresada en años. Se presentan tres grupos con diferencias estadísticas entre sus valores promedios, de acuerdo con la prueba de separación de medias por el método de Tukey (0.05). Bp y Ep no difieren entre sí y los valores de k son 0.223 y 0.267 respectivamente. Ap se encuentra en una posición intermedia con 0.437. Dv y Qh presentan los mayores valores de k con 0.683 y 0.767, respectivamente.
CRECIMIENTO FUNCIONAL
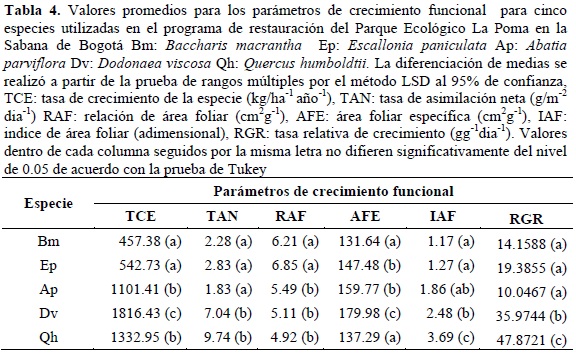
La tabla 4 muestra los valores promedios para los parámetros de crecimiento funcional de las especies objeto de evaluación. Para cada variable los valores promedios por especie dentro de cada columna seguidos por la misma letra no difieren significativamente del nivel de 0.05, de acuerdo con la prueba de Tukey. Para la TCE que expresa la tasa de acumulación de biomasa en términos de materia seca, Dv es la especie con el mayor valor promedio para las clases de edad con 1816.43 kg/ha-1 año-1, mientras que Bm y Ep son las especies que menor valor presentan con 457.38 y 542.73 kg/ha-1 año-1, Qh y Ap constituyen el grupo de especies con valores de TCE intermedios. En cuanto a la eficiencia fotosintética de la especies (TAN), la misma tabla muestra dos grupos de especies el primero conformado por Ap, Ep y Bm, con valores entre 1.83 y 2.83 g/m-2 dia-1, el segundo grupo constituido por Qh y Dv presenta la mayor eficiencia fotosintética con valores de asimilación de 12.74 y 8.04 g/m-2 dia-1. En cuanto a la magnitud del aparato fotosintético de las especies (RAF), se generan dos grupos el primero constituido por Bm y Ep, que requieren más de 6 cm2 de área foliar para construir un gramo de biomasa expresada como materia seca, en contraste con las demás especies que requieren menos de 5.5 cm2 de área foliar para construir la misma cantidad de biomasa. El AFE caracteriza las especies de acuerdo con el grosor de la lámina foliar; asimismo, indica que las hojas más gruesas corresponden a Dv, en contraste con Bm y Qh, que presentan las hojas más delgadas, y con Ep y Ap, que poseen hojas de grosor intermedio. En cuanto a la magnitud del área foliar por unidad de superficie (IAF), Qh genera más de 3 metros cuadrados de área foliar por m2 de superficie, en contraste con Bm y Ap que generan valores de IAF de 1.17 y 1.27 respetivamente, las demás especies generan IAF entre 1.86 y 2.48.
DISCUSIÓN
MORFOMETRÍA FOLIAR
Todas las especies evaluadas manifestaron para las variables A, P y SF diferencias estadísticas significativas que indican la presencia de diversidad foliar entre las poblaciones en general y dentro del dosel de los individuos en particular, lo que está acorde con lo propuesto por Taiz & Seiger (2006), quienes consideran este comportamiento como una estrategia adaptativa al régimen de luz cambiante. Por esto, la mayoría de las especies evaluadas desarrollaron hojas de mayor tamaño en el estrato apical de la copa y más redondeadas que las hojas que crecen en el estrato basal, las cuales tienden a ser rectangulares y pequeñas (Tabla 1). Lo anterior permite vislumbrar que dicho comportamiento no obedece a procesos aleatorios sino a un comportamiento determinístico, como consecuencia de la disponibilidad de luz en el entorno de crecimiento. En cuanto a la funcionalidad de la lámina foliar expresada por los valores del AFE, la mayoría de las especies arbustivas (Dv, Bm y Ep), presentan las hojas de mayor grosor en el estrato medio de la copa, lo que se convierte en un nuevo patrón que contrasta con lo propuesto por Terashima & Hikosaka (2005), quienes consideran que este tipo de hojas siempre se ubicaban en la porción apical de la copa.
Los resultados sobre acumulación y distribución del área foliar (A) en el dosel (Figura 1 y Tabla 2) permiten vislumbrar que las especies arbustivas (Dv, Ap, Ep y Bm), requieren de un periodo inferior a ocho años para construir su aparato fotosintético. Igualmente, se observa que a partir de este tiempo la biomasa generada se distribuye diferencialmente en otros compartimentos de la planta. Lo anterior permite fijar más temprano el carbono en órganos de la planta de mayor permanencia, como las ramas y la raíz. Esto coincide con lo encontrado por Brantley & Young (2007), para Morella cerifera que es una especie arbustiva nativa de la vegetación de la Costa Este de Norteamérica.
Igualmente, en la figura 2, se ratifica dicho comportamiento, puesto que las copas de las especies arbustivas cambian rápidamente de formas piramidales características de estados tempranos de desarrollo, a formas más compactas de tipo aparasolada, elípticas o en forma de cáliz, que corresponden a doseles de individuos maduros, como es descrito por Lowman & Rinker (2004) y Larcher (2003). Lo anterior contrasta para el caso de Qh, cuya tendencia de acumulación del área foliar se proyecta para un periodo superior a veinte años, lo que implica que estos árboles, tanto en la parte temprana como media de su ciclo de vida, lo dedican a construir su aparato fotosintético constituido principalmente de hojas y ramas de pequeñas dimensiones. Estos resultados coinciden con lo encontrado por Pretzsch (2009) y Gajardo-Caviedez et al. (2005), en estudios realizados en fabáceas de zonas templadas de Norteamérica con Quercus alba y con Nothofagus dombeyi en Suramérica.
ARQUITECTURA DE DOSEL
Las especies arbustivas leñosas no se cobijan bajo un solo patrón arquitectural de inserción foliar para el dosel, ya que manifiestan los tres tipos generales de arreglo tanto erectófilo, planófilo, como plagiófilo, lo que manifiesta el amplio rango de plasticidad funcional que les permiten adaptarse al entorno de crecimiento, como es propuesto por Pallardy (2008). Llama la atención que, en general, y sin importar el tipo de arreglo foliar, el dosel de todas las especies arbustivas presentan una alta absorción lumínica en el estrato basal a diferencia de lo que sucede en el dosel de los árboles forestales (Qh). Este comportamiento no se encuentra reportado en la literatura revisada y puede ser considerado como específico para las especies arbustivas estudiadas.
CRECIMIENTO FUNCIONAL
En cuanto a la productividad y la eficiencia fotosintética de las especies, tomando como base la tasa de crecimiento relativo (TCR), que relaciona la TAN y la RAF, de acuerdo, con los valores presentado en la tabla 4, se observa que el mayor valor lo genera Dv, con 0.726, es decir, que esta especie arbustiva por cada unidad de masa inicial, anualmente la incrementa por lo menos en un 72%, valores que superan la mayoría de los arboles forestales, incluso al mismo Qh, bajo el mismo entorno de crecimiento. Al comparar estos valores con resultados de trabajos similares desarrollados por Farmer (2006), en otras especies del genero Quercus de zonas templadas tales como Q. alba (17.4%) y Q. rubra (25.2%), además de algunas arbustivas del género Vaccinium (21.2%), en general, todas las especies arbustivas evaluadas en el presente estudio superan estos valores, lo que las identifica como altamente eficientes en los procesos productivos, aun en entornos ambientales críticos.
CONCLUSIONES
El presente estudio permitió evaluar el comportamiento ecofisiológico de cinco especies andinas nativas de la cordillera oriental colombiana (A. parviflora, D. viscosa, E. paniculata, B. macrantha y Q. humboldtii) de las cuales las cuatro primeras son arbustos leñosos, que se encuentran creciendo en un entorno ambiental con altas restricciones para el desarrollo. Para lograr lo anterior, estas especies arbustivas han generado una alta diversidad foliar como estrategia adaptativa al régimen de luz cambiante, basada principalmente, en el tamaño y grosor de las hojas, las cuales son dispuestas bajo un patrón arquitectural diferente al generado por los árboles forestales. Esto hace que requieran de un periodo más corto que el utilizado por los árboles, para construir su aparato fotosintético y acumular permanentemente el carbono capturado en compartimentos de larga permanencia. También se concluye que estas especies arbustivas no se cobijan bajo un solo patrón arquitectural de inserción foliar para el dosel, si no que, por el contrario, manifiestan los diferentes tipos de arreglos conocidos y, además, organizan las copas de manera que puedan capturar más eficientemente la luz, convirtiéndolas en especies altamente eficientes en los procesos funcionales productivos. Por lo anterior queda claro el gran potencial que tienen estas especies arbustivas, como de alto valor de uso en áreas de restauración con entornos ambientales críticos, lo que las convierte en especies clave para la mitigación del cambio climático global, bajo estas condiciones de crecimiento.
AGRADECIMIENTOS
Este trabajo fue realizado gracias a la financiación y apoyo del Programa Hojas Verdes de la Corporación Ambiental Empresarial, Filial de la Cámara de Comercio de Bogotá, al Parque Ecológico La Poma, por el apoyo de la logística de campo, al Laboratorio LASEREX de la Universidad del Tolima, por su apoyo científico y al laboratorio de Ciencias Forestales de la Universidad del Tolima por prestar los espacios de investigación científica para este proyecto.
REFERENCIAS BIBLIOGRÁFICAS
Ahuja, L.R., Reddy, V.R., Saseendran, S.A., & Quiang, Y. (2008). Response of crops to limited water: Understanding and modeling water stress effects on plant growth processes. Madison: American Society of Agronomy Inc. 436 p.
Begon, M., Townsend, C.R., & Harper, J.L. (2006). Ecology: From Individuals to Ecosystems (4th ed.). Melden-Oxford-Carlton: Blackwell Publishing Ltd. 738 p.
Blaser, J., Sarre, A., Poore, D., & Johnson, S. (2011). Status of Tropical Forest Management 2011 (Technical Series No. 38). Yokohama: International Tropical Timber Organization. 28 p.
Brantley, S.T., & Young, D.R. (2007). Leaf-area index and light attenuation in rapidly expanding shrub thickets. Ecology, 88(2), 524-530.
Castellanos, N.M. (2001). Programa piloto de adaptación de especies nativas para la restauración de ecosistemas de bosque andino de la cordillera oriental. Parque ecológico La Poma (Tesis inédita de pregrado). Ibagué: Universidad del Tolima. 115 p.
Cournac, L., Duboist, M., Chavet, J., & Riera, B. (2002). Fast determination of light availability and leaf area index in tropical forests. Journal of Tropical Ecology, 18, 295-302.
Fageria, N.K., Baligar, V.C., & Clarck, R.B. (eds.). (2006). Physiology of crop production. London: Routledge. 345 p.
Farmer, R.E. (2006). Comparative analysis of first year growth in six deciduous trees. Canadian Journal of Forestry Restoration, 36, 35-41.
Gajardo-Caviedez, P.A., Espinosa, M.A., González, U., & Ríos, D.G. (2005). The influence of thinning and tree size on the sapwood area/leaf area ratio in coigue. Canadian Journal Forest Restoration, 35, 1679-1685.
Hay, R., & Porter, J. (2006). The physiology of crop yield (2nd ed.). Oxford: Blackwell Publishing. 313 p.
Hopkings, W.G. (2006). Photosynthesis and respiration. New York: Chelsea House publishers. 169 p.
Hoover, C.H. (2008). Field measurements for forest carbon monitoring: A landscape-scale approach. Berlin: Springer. 236 p.
Landsberg, J.J., & Gower, S.T. (1997). Applications of physiological ecology to forest management. New York: Academic Press. 354 p.
Larcher, W. (2003). Physiology plant ecology: Ecophysiology and stress physiology of functional groups (4th ed.). Berlín: Springer. 513 p.
Lowman, M.D., & Rinker, H.B. (2004). Forest canopies (2nd ed.). Oxford: Elsevier Academic Press. 517 p.
Lüttge, U. (2008). Physiological ecology of tropical plants (2nd ed.). Berlin: Springer 458 p.
Lüttge, U., Beck, E., & Bartels, D. (2011). Plant desiccation tolerance: Analysis and synthesis, Vol. 215, Ecological studies. Berlin: Springer. 386 p.
McGarvey, R.C., Martin, T.A., & White, T.L. (2004). Integrating within-crown variation in net photosynthesis in loblolly and slash pine families. Tree Physiology, 24, 1209-1220.
Morison, J.I.L., & Morecroft, M.D. (2006). Plant growth and climate change. Oxford: Blackwell Publishing. 213 p.
Nouvellon, Y., Laclau, J.P., Epron, D., Kinana, A., Mabiala, A., Roupsard, O., Bonnefond, J.M., le Maire, G., Marsden, C., Bontemps, J.D., & Saint-Andre, L. (2010). Within-stand and seasonal variations of specific leaf area in a clonal Eucalyptus plantation in the Republic of Congo. Forest Ecology and Management, 259, 1796-1807.
Pallardy, S.G. (2008). Physiology of woody plants (3rd ed.). San Diego: Academic Press, Elsevier. 454 p.
Pessarakli, M. (2002). Handbook of plant and crop physiology (2nd ed.). New York: Marcel Decker Inc. 973 p.
Pretzsch, H. (2009). Forest dynamics, growth and yield. From measurement to model. Berlin: Springer. 664 p.
Ravindranth, N.H., & Oswald, M. (2008). Carbon inventory methods: Handbook for greenhouse gas inventory, carbon mitigation and roundwood production projects. Berlin: Springer. 315 p.
Riaño N.M., Tangarife, G., Osorio, O.I., Girald, J.F., Ospina, C.M., Obando, D., Gómez, L., & Jaramillo, L.F. (2005). Modelo de crecimiento y captura de carbono para especies forestales en el trópico. Bogotá: Cenicafé - Ministerio de Agricultura y Desarrollo Rural - Conif - Federación Nacional de Cafeteros de Colombia. 51 p.
Rodríguez, N. (2010). Determinación del carbono capturado por cinco especies arbóreas y arbustivas del bosque seco altoandino en el Parque Ecológico la Poma (Tesis inédita de pregrado). Ibagué: Universidad del Tolima. 85 p.
Sack, L., & Frole, K. (2006). Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees. Ecology, 87, 483-491.
Sack, L., & Holbrook, N.M. (2006). Leaf hydraulics. Annual Review of Plant Biology, 57, 361-381.
Salisbury, F.B., & Ross, C.W. (1992). Plant physiology (4th ed.). Belmont: Wadsworth Publishing Company. 682 p.
Sokal, R.R., & Rohlf, F.J. (2009). Introduction to biostatistics. New York: John Wiley and Sons, Inc. 768 p.
Taiz, L., & Zeiger, E. (2006). Plant physiology (4th ed.). New York: Sinauer Associates. 690 p.
Terashima, I., & Hikosaka, K. (2005). Comparative ecophysiology of leaf and canopy photosynthesis. Plant Cellular Environment, 18, 1111-1128.
Wareing, P.F., & Phillips, I.D.J. (2006). Growth and differentiation in plants (4th ed.). Oxford: Pergamon Press. 343 p.




Licencia
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






