
DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2012.2.a06Publicado:
01-07-2012Número:
Vol. 15 Núm. 2 (2012): Julio-DiciembreSección:
Artículos de investigación científica y tecnológicaINDICADORES MORFOMÉTRICOS FOLIARES PARA LA DETECCIÓN TEMPRANA DEL ESTRÉS HÍDRICO DE Gmelina arborea Roxb. EN SISTEMAS AGROFORESTALES, SANTAFÉ DE ANTIOQUIA
Foliar morphometric indicators for early detection of water stress in Gmelina arborea Roxb. in agroforestry systems of Santafé de Antioquia
Palabras clave:
asimetría foliar, bosque seco tropical, fenotipo foliar, inestabilidad del desarrollo. (es).Palabras clave:
leaf asymmetry, tropical dry forest, leaf phenotype, developmental instability. (en).Descargas
Referencias
Alados, C.L. (2009). Inestabilidad en el desarrollo y cambio global en plantas. Revista Real Academia de Ciencias. Zaragoza, 64, 35-49.
Alados, C.L., & Aich, A.E. (2008). Stress assessment of argan (Argania spinosa L. Skeels) in response to land uses across an aridity gradient: Translational asymmetry and branch fractal dimension. Journal of Arid Environments, 72, 338-349.
Alados, C.L., Giner, M.L., Dehesa, L., Escós, J., Barroso, F.G., Emlen, J.M., & Freeman, D.C. (2002). Developmental instability and fitness in Periploca laevigata experiencing grazing disturbance. International Journal of Plant Sciences, 163(6), 969-978.
Aldana, J.A. (2008). Evaluación y selección fenotípica de árboles superiores en rodales de Gmelina arborea en el valle geográfico del Rio Magdalena (Tesis de pregrado, Ingeniería Forestal). Ibagué: Universidad del Tolima. 125 p.
Arias, D., Murillo, O., & Rojas, F. (2004). Manual para productores de melina (Gmelina arborea) en Costa Rica. Cartago: Fonafifo. 120 p.
Burrows, G.E. (2001). Comparative anatomy of the photosynthetic organs of 39 xeromorphic species from subhumid New South Wales, Australia. International Journal of Plant Sciences, 162(2), 411-430.
Dale, V.H., & Beyeler, S.C. (2001). Challenges in development and use of ecological indicators. Ecological Indicators, 1, 3-10.
Emlen, J.M., Freeman, D.C., & Graham, J.H. (1993). Nonlinear growth dynamics and the origin of fluctuating asymmetry. Genetica, 89, 77-96.
Fageria, N.K., Baligar, V.C., & Clarck, R.B. (2006). Physiology of crop production. London: FPP. 345 p.
Fair, J.M., & Breshears, D.D. (2005). Drought stress and fluctuating asymmetry in Quercus undulata leaves: confounding effects of absolute and relative amounts of stress? Journal of Arid Environments, 62, 235-249.
Freeman, D.C., Brown, M.L., Duda, J.J., Graham, J.H., Emlen, J.M., Krzysik, A.J., Balbach, H.E., Kovacic, D.A., & Zak, J.C. (2004). Photosynthesis and fluctuating asymmetry as indicators of plant response to soil disturbance in the fall-line sandhills of Georgia: a case study using Rhus copallinum and Ipomoea pandurata. International Journal of Plant Sciences, 165(5), 805-816.
Freeman, D.C., Brown, M.L., Duda, J.J., Graham, J.H., Emlen, J.M., Krzysik, A.J., Balbach, H., Kovacic, D.A., & Zak, J.C. (2005). Leaf fluctuating asymmetry, soil disturbance and plant stress: a multiple year comparison using two herbs, Ipomoea pandurata and Cnidoscolus stimulosus. Ecological Indicators, (5), 85-95.
Kozlov, M.V., Niemela, P., & Malkonen, E. (2002). Effects of compensatory fertilization on pollution-induced stress in Scots pine. Water Air and Soil Pollution, 134, 307-318.
Larcher, W. (2003). Physiological plant ecology. New York: Springer Verlag. 520 p.
Lorenzo, N., Mantuano, D.G., & Mantovani, A. (2010). Comparative leaf ecophysiology and anatomy of seedlings, young and adult individuals of the epiphytic aroid Anthurium scandens (Aubl.) Engl. Environmental and Experimental Botany, 68, 314-322.
Mogie, M., & Cousins, M. (2001). Are sample sizes usually at least an order of magnitude too low for reliable estimates of leaf asymmetry? Journal of Theoretical Biology, 211, 181-185.
Møller, A.P. (2006). A review of developmental instability, parasitism and disease Infection genetics and evolution. Infection, Genetics and Evolution, 6, 133-140.
Páez, Q.E., & Barreiro, H.D. (1986). Análisis del régimen de lluvias de la estación experimental “El Espinal” Centro Cotové. (Trabajo de pregrado, Ingeniería Agrónoma). Universidad Nacional de Colombia, sede Medellín. Medellín: Facultad de Ciencias Agropecuarias. 120 p.
Palmer, A.R., & Strobeck, C. (1986). Fluctuating asymmetry: measurement, analysis, patterns. Annual Review of Ecology and Systematic, 17, 391-421.
Pérez-Contreras, T., Soler, J. J., & Soler, M. (2008). Needle asymmetry, pine vigour and pine selection by the processionary moth Thaumetopoea pityocampa. Acta Oecologica, 33, 213-221.
Rettig, J.E., Fuller, R.C., Corbett, A.L., & Getty, T. (1997). Fluctuating asymmetry indicate levels of competition in an even-aged poplar clone. Oikos, 80, 123-127.
Sack, L., & Frole, K. (2006). Leaf structural diversity is related to hydraulic capacity trees. Ecology, 87(2), 483-491.
Stuckens, J., Verstraeten, W.W., Delalieux, S., Swennen, R., & Coppin, P. (2009). A dorsiventral leaf radiative transfer model: Development, validation and improved model inversion techniques. Remote Sensing of Environment, 113, 2560-2573.
Tamas, J., & Hably, L. (2009). Unidirectional shift in leaf morphology of coexisting species: a possible indicator of palaeoclimatic differences. Palaeogeography, Palaeoclimatology, Palaeoecology, 27, 185-195.
Velickovic, M.V. (2005). Does the coefficient of variation reflect developmental instability in Plants? Archives of Biological Science Belgrade, 57 (3), 7-8.
Villar, R., Ruiz, J., Quero, J.L., Poorter, H., Valladares, F., & Marañón, T. (2004). Tasas de crecimiento en especies leñosas: aspectos funcionales e implicaciones ecológicas. En: Valladares, F. (2004). Ecología del bosque mediterráneo en un mundo cambiante (pp. 191-227). Madrid: Ministerio de Medio Ambiente, EGRAF S.A.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
 | Melo-Cruz, O., Martínez-Bustamante, E., & Correa-Londoño, G. (2012). Indicadores morfométricos foliares para la detección temprana del estrés hídrico de Gmelina arborea Roxb. en sistemas agroforestales, Santafé de Antioquia. Colombia Forestal, 15(2), 215-226. |
INDICADORES MORFOMÉTRICOS FOLIARES PARA LA DETECCIÓN TEMPRANA DEL ESTRÉS HÍDRICO DE Gmelina arborea Roxb. EN SISTEMAS AGROFORESTALES, SANTAFÉ DE ANTIOQUIA
Foliar morphometric indicators for early detection of water stress in Gmelina arborea Roxb. in agroforestry systems of Santafé de Antioquia
Omar Melo-Cruz1, Enrique Martínez-Bustamante2 & Guillermo Correa-Londoño3
1Grupo de investigación en Biodiversidad y Dinámica de Ecosistemas Tropicales, Universidad del Tolima, Ibagué. Grupo de investigación en Ecofisiología Agraria, Universidad Nacional de Colombia, sede Medellín. Ibagué, Colombia. omelo@ut.edu.co. Autor para correspondencia
2Departamento de Ciencias Agronómicas, Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, sede Medellín. Medellín, Colombia. enmartin@unal.edu.co
3Departamento de Ciencias Agronómicas, Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, sede Medellín. Medellín, Colombia. gcorrea@unal.edu.co
Recepción. 23 de octubre de 2012 / Aprobación. 15 de noviembre de 2012
RESUMEN
En esta investigación se evaluó la asimetría foliar (ASF) en individuos de Gmelina arborea Roxb., establecidos en sistemas agroforestales (SAF), en áreas de bosque seco tropical, en el municipio de Santafé de Antioquia. Estos individuos crecen bajo diferentes condiciones hídricas del suelo. La ASF se correlacionó con nuevos parámetros de tipo morfométrico como el ángulo de inserción del peciolo en la lámina foliar (AIP) en individuos maduros y la razón entre el número de dientes del borde de la lámina (RDF) en las hojas de individuos juveniles, propuestos como nuevos indicadores del estado temprano de estrés en la especie. Los modelos generados presentaron valores de R2 superiores al 75 %, lo que soporta su utilización como indicadores tempranos del estrés hídrico en G. arborea. Adicionalmente, se evaluó la diversidad foliar entre los dos tipos de hojas (HM y HJ), con variables morfométricas convencionales, tales como la longitud de la nervadura principal (L), perímetro de la hoja (P), área foliar específica (AFE) y materia seca (DW), puesto que son caracteres relacionados con la forma y funcionalidad foliar. De lo anterior se dedujo que existen marcadas diferencias en forma y comportamiento entre los dos tipos de hojas. Los resultados de este trabajo permitirán la optimización de estrategias de manejo de G. arborea en el SAF, gracias a la identificación temprana de la presencia de este tipo de estrés. Esto permitiría mantener permanentemente el dosel de los árboles y así mejorar el ambiente de productividad de los cultivos agrícolas asociados.
Palabras clave: asimetría foliar, bosque seco tropical, fenotipo foliar, inestabilidad del desarrollo.
ABSTRACT
Leaf asymmetry was assessed in individuals of Gmelina arborea Roxb. growing under different soil water conditions in a dry forest agroforestry system (AFS), in Santafé de Antioquia. Leaf asymmetry was correlated with additional morphometric parameters, such as angle of insertion of the petiole in the leaf in mature individuals and the ratio of the number of teeth on the edge of the blade in juvenile leaves to propose new indicators of early stage stress. The models generated had R2 values of above 75 %, which support their use in identifying developmental instability as an early indicator of water stress. Similarly, leaf diversity was assessed between the two phenotypes of leaves (ML and JL), with conventional morphometric variables (length of the midrib, leaf perimeter, specific leaf area and dry matter), because they are characters related to leaf form and function. There were marked differences in form and behavior between the two types of leaf indicating a further source of instability evident from leaf ontogeny. The results of this work will allow the optimization of management strategies of G. arborea in the AFS as an early indicator of water stress.
Key words: leaf asymmetry, tropical dry forest, leaf phenotype, developmental instability.
INTRODUCCIÓN
La inestabilidad del desarrollo es un término que se utiliza para identificar la incapacidad de un organismo para soportar perturbaciones aleatorias durante el proceso de su desarrollo, por lo cual su cuantificación se convierte en un indicador potencial del estado temprano de estrés y permite inferir las relaciones de los individuos con su entorno ambiental (Alados & Aich, 2008). Por lo anterior, al evaluar la inestabilidad del desarrollo dentro de una población, se pueden identificar los individuos que tienen la capacidad de adaptarse mejor a un ambiente particular (Møller, 2006).
Para cuantificar la inestabilidad del desarrollo referida a la estimación del estrés ambiental en muchas especies, se ha utilizado la fluctuación asimétrica que corresponde a pequeñas desviaciones aleatorias de la simetría bilateral que hace referencia a las dos mitades (izquierda y derecha), en las que se puede dividir un individuo o uno de sus órganos al trazar un eje central. Estas fluctuaciones asimétricas se han utilizado como herramientas para el monitoreo ecológico del estrés, entre y dentro de poblaciones (Rettig et al., 1997; Burrows, 2001). Lo anterior requiere de trabajos exhaustivos y detallados de gran magnitud para evitar el ruido estadístico cuando se definen las caras o mitades objetos de evaluación (Mogie & Cousins, 2001). Por esto se hace necesario explorar nuevos indicadores más sencillos, que correlacionen las fluctuaciones asimétricas con parámetros morfométricos de los órganos, con el fin de lograr una estimación adecuada de la inestabilidad del desarrollo (Velickovic, 2005). Sin embargo, es claro que no hay consenso sobre el tipo de indicador que se puede desarrollar o los puntos de referencia a tener en cuenta para ese fin (Alados & Aich, 2008). No obstante, se debe partir de la premisa de que si la asimetría foliar (ASF) es el resultado de las fluctuaciones de los valores que caracterizan la lámina foliar cuando se comparan con su valor estándar, lo cual genera una distribución normal para los valores absolutos de dichas diferencias y cuya media es cero (Palmer & Strobeck, 1986), se esperaría que cualquier parte de la hoja o parámetro morfométrico registrado en esta manifieste dicha asimetría y, por consiguiente, la inestabilidad del desarrollo, lo que facilitaría su identificación y cuantificación.
Por lo anterior, se supone que dichos mecanismos, para el caso de las plantas, actúan con gran dependencia de las condiciones ambientales y están enfocados principalmente hacia a la adaptación a la sequía. Sin embargo, los cambios en las condiciones ambientales podrían conducir a una interacción entre los mecanismos expuestos o incluso a una supuesta diferenciación paulatina a lo largo del gradiente ambiental (Alados & Aich, 2008). Por esto se ha propuesto que la ASF se incrementa al disminuir la disponibilidad de humedad en el suelo, mientras que dichas fluctuaciones morfométricas disminuyen bajo condiciones ambientales más estables (Lorenzo et al., 2010). De esta manera, lo que se esperaría a lo largo de un gradiente de humedad es que la ASF aumente proporcionalmente con la disminución de la humedad del suelo.
El presente estudio se llevó a cabo en poblaciones de Gmelina arborea, con individuos en diferente estado de desarrollo tanto a nivel de vivero, como de árboles establecidos en campo, que hacen parte de un sistema agroforestal (SAF) y cuya función principal es la de generar sombra sobre el componente agrícola (Theobroma cacao), para estimular los procesos reproductivos que conduzca a la máxima producción de grano. El déficit hídrico en G. arborea genera defoliación total de los árboles lo que afecta las condiciones reproductivas para T. cacao al incrementar tanto la temperatura, como el índice de radiación sobre el cultivo. Por esto se ha propuesto generar y validar nuevos indicadores morfométricos foliares para la detección temprana del estrés hídrico en G. arborea y poder contribuir así a la optimización del uso del agua en un entorno ambiental donde su disponibilidad es crítica, tal como son los bosques secos tropicales. Los interrogantes de investigación se enfocan en resolver si existe evidencia de asimetría foliar en G. arborea y si la ASF que se identifique está asociada con la disponibilidad de humedad en el suelo. Con lo anterior se espera contribuir a la identificación de herramientas y oportunidades de manejo de sistemas productivos más amigables con el entorno ambiental afectados por un clima cambiante.
MATERIALES Y MÉTODOS
ÁREA DE ESTUDIO
Esta investigación se realizó en el Centro Agropecuario Cotové, cuya extensión es de 121.16 ha y se encuentra adscrito a la Universidad Nacional de Colombia, sede Medellín. El Centro se encuentra ubicado en la vereda El Espinal, municipio de Santafé de Antioquia, a 76 km al occidente de la ciudad de Medellín. Sus coordenadas geográficas son 6°33�32� latitud norte - 77°04�51� longitud oeste, lo que lo ubica en la zona media del valle geográfico del río Cauca. Ecológicamente, se encuentra en la zona de vida bosque seco tropical (Bs-T), con una altitud aproximada de 540 m, una temperatura promedio anual de 27°C y una precipitación media anual de 1031 mm (Páez & Barreiro, 1986).
POBLACIONES DE TRABAJO
Las evaluaciones se realizaron sobre individuos juveniles tanto en nivel de vivero, que crecen bajo condiciones óptimas de humedad suministradas por riego por microaspersión, como en individuos establecidos inicialmente en campo y que se encuentran sometidos a cambios ambientales típicos del trasplante. A nivel de plantación se evaluaron individuos de dos años de edad que constituyen el componente arbóreo de un sistema agroforestal (SAF) cuyo cultivo principal es T. cacao. A este sistema se le ha suministrado agua a través de un sistema autónomo de riego, esto ha reducido el déficit hídrico al que naturalmente se encuentran sometidas las especies vegetales de los bosques secos tropicales, al permanecer el suelo a capacidad de campo. Igualmente, se evaluaron individuos G. arborea con edades equivalentes que hacen parte de un experimento de monitoreo y acumulación de biomasa, en un área cuya oferta hídrica es suministrada por el entorno natural y que, por consiguiente, está sujeta a procesos de defoliación como respuesta al estrés por sequía. Los G. arborea pertenecen a la familia Verbenaceae y es un árbol tropical originario del sudeste asiático, el cual presenta un acelerado crecimiento asociado a buenas características en la calidad de su madera. Esto lo ha convertido en una especie de gran importancia económica para el establecimiento de plantaciones comerciales a gran escala o asociado en arreglos agroforestales en toda la región tropical (Arias et al., 2004).
En las zonas secas, G. arborea desarrolla una fenología de tipo caducifolio, con una copa cónica y amplia en sitios abiertos como los sistemas agroforestales, pero en monocultivos su copa es densa y compacta. G. arborea presenta diversidad foliar ya que las hojas de los individuos juveniles tienen láminas simples de borde dentado y usualmente acorazonadas, decoloradas, el haz verde y glabras. En contraste cuando los individuos superan la etapa de establecimiento en campo, las hojas desarrollan borde entero y tienden a ser más coriáceas, el envés se vuelve verde pálido y aterciopelado, genera una venación reticulada, con nervaduras secundarias arregladas entre 4 y 10 pares y no presentan estípulas (Aldana, 2008).
TAMA�O DE LA MUESTRA
Las cuantificaciones biogeométricas se realizaron sobre 4 hojas funcionales de G. arborea, cosechadas en el tercio medio de la copas de 100 individuos, en el que se seleccionaron cuatro ramas ubicadas en posición cardinal y de la parte media de estas se cosechó una hoja funcional sin herbivoría, daño mecánico o ataque patógeno. El valor representativo del individuo lo constituyó el promedio de las 4 hojas para la variable asimetría foliar (ASF), esta se determinó como el promedio de las diferencias entre los anchos del lado izquierdo y derecho de la lámina foliar medida en milímetros; la ASF se expresa en términos absolutos como |I-D| (Alados & Aich, 2008). Los 100 individuos se clasificaron en dos grupos: 50 individuos juveniles (25 ubicados a nivel de vivero y 25 recientemente establecidos en campo), los cuales se caracterizan por generar hojas de borde dentado y para el caso del experimento se identificaron como hojas juveniles (HJ). Los restantes 50 individuos corresponden al estado establecido y adulto que ha superado la etapa de adaptación en campo y que para la fecha del experimento tenían dos años de edad. Estos individuos se caracterizan por generar hojas de borde entero, coriáceas, de color opaco y con presencia de tricomas por el envés y para el caso del experimento se denominarón hojas maduras (HM). Para esta categoría se evaluaron 25 individuos bajo condición permanente de humedad de suelo a capacidad de campo y los restantes en el área con oferta hídrica del entorno natural y que se encuentran sometidos a los procesos naturales de defoliación. Esto generó un gradiente de humedad que va desde un entorno de vivero con condiciones de humedad controladas, hasta plantaciones en SAF sin subsidio hídrico. Un primer SAF suministró riego al suelo a capacidad de campo, mientras que los individuos que crecen en el segundo SAF permanecen sometidos al régimen hídrico natural del Centro Cotové con estrés hídrico. Para el primer caso, la disponibilidad hídrica estuvo regida por los sistemas de riego tanto del vivero, como del cultivo (SSH) y, para el segundo caso, sólo a la oferta natural del entorno ambiental del área de estudio (CSH).
VARIABLES
Cada tipo de hoja se caracterizó a partir de la evaluación de variables morfométricas que con frecuencia se utilizan en este tipo de trabajo para generar las comparaciones respectivas entre estas y las correlaciones con la ASF, con el propósito de identificar un indicador de la inestabilidad del desarrollo (Alados & Aich, 2008). Las variables morfométricas evaluadas por tipo de hoja y condición de humedad fueron: longitud de la nervadura principal (L), la cual se tomó como eje principal para dividir la lámina foliar en las dos mitades (derecha-izquierda), en la determinación de la fluctuación asimétrica (Møller, 2006). Ancho de la hoja en la base, a 1/3, 1/2 y 2/3 de la longitud total de la nervadura principal, medidas en la cara abaxial, en centímetros con aproximación al milímetro, para lo cual se empleó un calibrador digital de lectura directa. Esta información se utilizó como base para la determinación del ASF (Stuckens et al., 2009). Como nuevas variables para este tipo de estudio y no reportadas en la literatura, se propusieron el ángulo de inserción del peciolo (AIP) para el tipo HM, registrado como el ángulo externo generado entre la proyección de la nervadura principal, la base de la hoja y el peciolo, para lo cual se utilizó un trasportador de unidad angular. Para el tipo de hoja HJ se registró el número de dientes en el borde de la lámina foliar tanto para el lado izquierdo, como para el lado derecho, lo cual generó la razón entre el menor y mayor número de dientes (RDF).
También se evaluaron variables funcionales típicas de la hoja que se relacionan principalmente con la fisiología de la producción de plantas sometidas a estrés (Fageria et al., 2006). Entre estas se registraron el perímetro foliar (P) y área foliar (ARF), medidas con los analizadores LI-3000C Portable Area Meter y CI-200 Laser Leaf Area Meter, cuyas lecturas se registraron en cm y cm2, respectivamente. Peso de la materia seca de la lámina foliar (DWF), registrada en balanza digital con aproximación a miligramos; área foliar específica (AFE), generada a partir de la relación entre el área de la lámina foliar y su respectivo peso de materia seca expresada en metros cuadrados por kilogramo (Fair & Breshears, 2005).
ESTADÍSTICAS
Para determinar la presencia de la inestabilidad del desarrollo en G. arborea se propuso una prueba de t-student (α = 0.05), para comparación múltiple de medias en la variable ASF. Su hipótesis nula indica que los valores de las medias para la variable objeto de evaluación en los diferentes tipos de hojas (HM y HJ), con diferentes condiciones de humedad del suelo (SSH y CSH), son iguales a cero y la hipótesis alternativa propone que, por lo menos, una de las medias es diferente a cero.
Para determinar si la condición de humedad del suelo (SSH y CSH) influye sobre la ASF se propuso un ANOVA de múltiples factores, con un nivel de significancia del 95 % (α = 0.05) y prueba de rango múltiple de medias por el método LSD (95 %). Esto se hizo con el propósito de validar en G. arborea la dependencia entre el estrés hídrico y la fluctuación asimétrica.
Para confrontar la diversidad foliar entre HM y HJ en G. arborea para las condiciones del experimento se generaron modelos individuales de regresión polinomial, que describen la biogeometría de la lámina foliar al correlacionar las variables P, ARF y DWF con L (Mogie & Cousins, 2001). El variación morfológica de la lámina foliar se visualizó al graficar la relación entre el área foliar total (AFT), tanto para HM, como para HJ respecto a L. Se comparó la funcionalidad de las hojas a partir de una prueba de medias para la variable área foliar específica (AFE), la cual influye en el crecimiento de las plantas al favorecer la eficiencia fotosintética en el uso del nitrógeno y la demanda de luz (Larcher, 2003).
Como propuesta de nuevos indicadores para identificar la inestabilidad del desarrollo en G. arborea, se evaluaron el ángulo de inserción del peciolo (AIP) para HM y la razón de dientes foliares (RDF) para HJ, los cuales se correlacionaron con la ASF a partir de modelos lineales simples y su evaluación se realizó con base en los valores de R2 y el error estándar. Además, se realizó un ANOVA de múltiples factores, con un nivel de significancia del 95 % (α = 0.05), tanto para la variable AIP, como para RDF, con el propósito de determinar la sensibilidad de las nuevas variables propuestas frente a la condición de humedad del suelo (SSH y CSH). Para todos los procesos estadísticos se utilizó el software Statgraphics Centurion Versión 16.15.
RESULTADOS
De acuerdo con los resultados que se muestran en la figura 1, queda claro que la inestabilidad del desarrollo se manifiesta ampliamente en los dos tipos de hojas generadas tanto en las plántulas (HJ), como en los árboles (HM) de G. arborea, lo cual se expresa en términos de asimetría foliar (ASF). Igualmente, en esta figura se observa que tanto en condiciones de humedad del suelo a capacidad de campo (SSH), como en las condiciones sometidas a la oferta ambiental del entorno natural (CSH), se manifiesta la inestabilidad del desarrollo, pero con diferentes niveles de magnitud. Por lo anterior, de acuerdo con lo planteado en la prueba de t-student (α = 0.05), se rechaza la hipótesis nula y se acepta la hipótesis alternativa, puesto que todos los valores promedios para la ASF en los factores objeto de evaluación fueron diferentes a cero. Sin embargo, es de resaltar que las hojas que se desarrollan en las plántulas a nivel de vivero (HJ), bajo condiciones relativamente controladas de humedad (SSH), son las que menor variación presentan en la ASF (2.37 � 1.13 mm), mientras que las HM que se desarrollan en condición de humedad CSH mani-fiestan los mayores valores (13.52 � 4.75 mm) e incluso presentan valores extremos que superan los 25 mm.
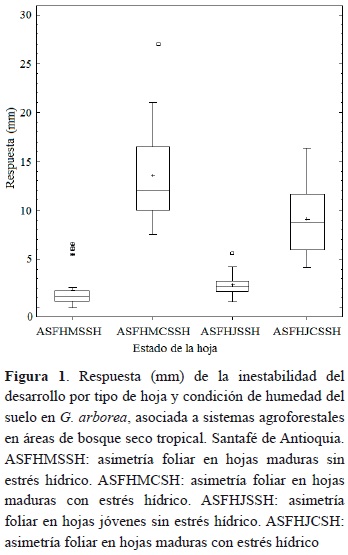
Los resultados estadísticos, que se muestran en la tabla 1 dejan de manifiesto que existe una influencia directa de la condición de humedad del suelo sobre los valores de la ASF para los dos tipos de hoja (HM y HJ), puesto que se presentan diferencias estadísticas significativas al 95 % (α = 0.05), entre las hojas desarrolladas por individuos que crecen bajo la condición de humedad del suelo SSH y CSH. Igualmente, llama la atención que las mayores variaciones en la asimetría se dan bajo la condición de humedad de suelo CSH con valores del CV % de 32.7 y 35.2 para las HJ y HM, respectivamente, mientras que los individuos que crecen bajo una mejor condición de humedad presentan hojas con una mayor estabilidad en la ASF. En general, todas las hojas de G. arborea presentan ASF, ellas manifiestan mayor sensibilidad cuando la condición de humedad del suelo disminuye.
La tabla 2 muestra los modelos de regresión polinomial que relacionan la biogeometría de la lámina foliar de los dos tipos de hoja (HM y HJ) generados por G. arborea. Los modelos generales que relacionan a P, ARF y DW con L se corrieron para cada tipo de hoja dando como resultados ajustes que van desde el 86 hasta el 98 %; sin embargo, las tendencias de los modelos son inversas entre los tipos de hojas. Esto se aprecia en los valores de los interceptos de los modelos, los cuales son positivos para las HM, mientras que asumen un valor negativo para las HJ, lo cual vislumbra un comportamiento diferencial entre estas, que caracterizaría la diversidad foliar de G. arborea.

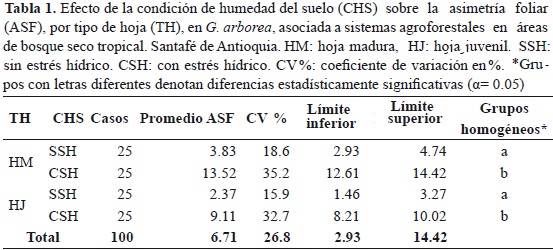
La figura 2 contrasta la variación morfológica de la lámina foliar en G. arborea tanto para HM, como para HJ, al relacionar el área total de la lámina foliar (AFT) con la longitud de la nervadura principal (L). Esta es una expresión del factor de forma foliar que indica, para este caso, que a medida que las HM aumentan de tamaño tienden a formas redondeadas, mientras que las HJ más grandes tienden a ser más elípticas y rectangulares. Cuando las hojas de G. arborea son de tamaño pequeño la relación es inversa.
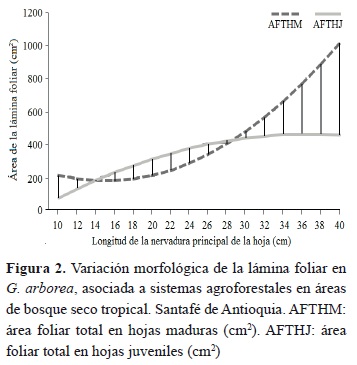
El comportamiento funcional, a partir del análisis del AFE de los dos tipos de hojas (HM y HM), se describe en la figura 3. Este rasgo aparentemente simple de la planta, que corresponde al cociente entre la superficie y el peso de su hoja, puede explicar en ocasiones hasta el 80 % de la variación en sus tasas de crecimiento. Las mayores frecuencias del AFE para las HM varían entre 12 y 15 m2 kg-1 y en las HJ dichos valores se ubican entre 21 y 25 m2 kg-1. Lo anterior demuestra que las HM son más eficientes desde el punto de vista funcional al requerir menos área foliar para generar una unidad de materia seca foliar, mientras que las HJ requieren casi de un 75 % más de área foliar para producir la misma unidad de materia seca.
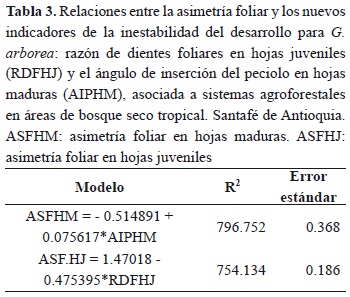
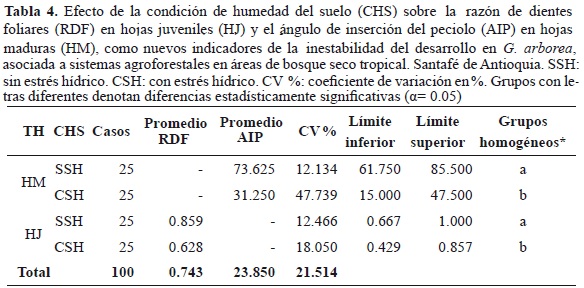
Los nuevos indicadores morfológicos foliares AIP y RDF, propuestos para identificar la inestabilidad del desarrollo en G. arborea, se correlacionaron en forma independiente con la ASF (Tabla 3); de esta forma muestran valores de R2 que varían entre el 75 y el 80 % y que indican la alta correlación para las nuevas variables planteadas; es decir, que tanto el AIP, para HM; como la RDF, para las HJ, son descriptores de la inestabilidad del desarrollo en G. arborea. El efecto de la condición de humedad del suelo (CHS) sobre los dos nuevos indicadores propuestos se muestra en la tabla 4. Los análisis estadísticos muestran que existen diferencias significativas al 95 % de confianza (α = 0.05), cuando las hojas se desarrollan en individuos sometidos a condiciones de humedad del suelo contrastantes. Igualmente, tanto para AIP en HM como para RDF en HJ presentan los mayores CV % bajo la condición de humedad del suelo CSH. Para el caso de AIP, las diferencias entre las medias para SSH y CSH superan el 100 %, mientras que para RDF, dichas diferencias son inferiores al 50 %. Estos resultados muestran la alta sensibilidad tanto de AIP, como de RDF frente a entornos de crecimiento con restricción hídrica.
DISCUSIÓN
La inestabilidad en el desarrollo es la incapacidad de un organismo de reducir los errores durante el crecimiento y es una medida que integra la facultad para resistir el estrés y junto con las fluctuaciones del entorno ambiental, frente a la carencia de energía suficiente para mantener el equilibrio homeostático, dan lugar al incremento de las variaciones intraindividuales de estructuras con idéntica influencia genética y ambiental. Esto ha permitido utilizar este principio para detectar el estrés ambiental en muchas especies, al alterar el desarrollo a lo largo de su trayectoria de vida en un ambiente particular (Palmer & Strobeck, 1986; Emlen et al., 1993, Alados, 2009). Para el caso de G. arborea en el presente estudio, cuyos individuos crecen en áreas de bosque seco tropical asociados a un sistema agroforestal (SAF), en suelos con diferentes condiciones de humedad, la inestabilidad en el desarrollo se manifiesta tanto en la asimetría bilateral de las láminas foliares (Figura 1, Tabla 1), como en otros rasgos morfométricos, tales como el ángulo de inserción del peciolo respecto a la base de la hoja en HM, al igual que la variación del número de dientes foliares ubicados en el borde de las HJ (Tabla 4), por lo cual se espera que dichas fluctuaciones correspondan a la respuesta de los individuos a las condiciones de estrés ambiental causadas por las restricciones hídricas de su entorno de crecimiento. Dicha afirmación se genera a partir de las marcadas diferencias que se presentan en estos indicadores morfométricos, cuando los individuos de G. arborea se someten a condiciones contrastantes de humedad del suelo. Esto es afín con los resultados encontrados para Argania spinosa y Quercus undulata, en estudios realizados por Alados & Aich (2008) y Fair & Breshears (2005), en bosques mediterráneos que presentan equivalencia ecológica con los bosques secos tropicales.
En cuanto a las variaciones del valor de la inestabilidad en el desarrollo para los indicadores morfométricos foliares utilizados en el presente estudio (Tabla 1 y Tabla 4), se encontró que los diferentes tipos de hojas evaluadas (HM y HJ) manifiestan las mayores variaciones cuando los individuos de G. arborea crecen en suelos con condición de humedad CSH. Estos resultados concuerdan con lo reportado por Alados (2009), al estudiar poblaciones naturales de A. spinosa localizadas en las zonas más áridas de su distribución natural en el norte de África, en comparación con poblaciones establecidas en zonas más húmedas sometidas a procesos agrícolas.
En cuanto a la diversidad que se manifiesta en la biogeometría foliar de G. arborea (Tabla 2, Figuras 2 y 3), esta obedece a respuestas fisiológicas de las plantas al estrés hídrico, cuya magnitud depende del rango de sequia al que las plantas estén adaptadas. De esta manera, los individuos que han tenido estrés hídrico frecuentemente son capaces de resistir mejor la sequia, en contraste con otros que no se enfrentan a tales situaciones, por lo que los primeros individuos manifiestan una mayor variación en los indicadores morfomértricos foliares en comparación a los que crecen en entornos ambientales mas estables (Sack & Frole, 2006). Es así como las plantas que crecen regularmente sometidas a estrés hídrico generan estrategias de supervivencia que reducen su crecimiento, para mantener el funcionamiento de las estructuras fundamentales para su capacidad de supervivencia y permanencia en el tiempo, en comparación con las plantas que viven en condiciones ambientales más favorables, las cuales presentan estrategias competitivas en las que invierten más energía en la producción y desarrollo y soló pueden mantener la estabilidad bajo condiciones ambientales más favorables (Alados, 2009). Para el caso de G. arborea, en el presente estudio, esto se manifiesta en la distribución diferencial del AFE para los diferentes tipos de hojas (Figura 3), lo cual tendría un significado adaptativo, de tal forma que las HJ, las cuales tienen los más altos valores de AFE (25 m2 kg-1), tendrían ventajas en hábitats productivos a costa de una menor longevidad foliar, mientras que las HM con baja AFE (15 m2 kg-1) son las que pueden presentar una mayor longevidad foliar y tendrían ventajas en hábitats con escasez de nutrientes y agua. Esto coincide con lo demostrado por Villar et al. (2004) para un grupo de 24 especies leñosas típicas de la península ibérica, en las que vinculan las morfometría foliar con el desarrollo funcional.
Actualmente, el empleo de nuevos indicadores ecológicos está generando un gran interés en la comunidad científica, probablemente a causa de la tendencia a la degradación del medio ambiente en todo el mundo. El calentamiento global y los cambios antropogénicos generalizados son las dos principales fuerzas que actúan sobre la dinámica del ecosistema, con implicaciones de gran importancia para la conservación de este último, donde la temperatura y el agua son los factores clave que regulan los procesos biológicos (Dale & Beyeler, 2001; Freeman et al., 2004; Alados, 2009). Es por esto que los estudios sobre inestabilidad en el desarrollo de plantas cobran gran importancia y están siendo utilizados para identificar los efectos de la actividad antrópica sobre la vegetación, los efectos de la sequía sobre el crecimiento de las plantas, la selección de fenotipos con capacidad de adaptación a ambientes con restricciones ambientales, indicadores de contaminación e incluso para la clasificación y separación de grupos funcionales de plantas fósiles en estudio de paleogeografía (Alados et al., 2002).
Al proponer el AIP y la RDF como nuevos indicadores morfométricos foliares para la detección temprana del estrés hídrico en HM y HJ para G. arborea y comprobar su alta correlación con la asimetría foliar (Tabla 3), ellos se convierten en medidas de identificación para la inestabilidad en el desarrollo y constituyen una nueva herramienta para la detección temprana del estrés hídrico para la especie en estas áreas de bosque seco tropical. De igual forma, Kozlov et al. (2002) propuso diferenciales de materia seca en acículas de Pinus sylvestris, para evaluar el efecto del estrés por contaminación sobre sus poblaciones naturales. Tamas & Hably (2009) estudiaron los cambios unidireccionales en la morfología foliar como un posible indicador de las diferencias paleoclimáticas de especies de plantas coexistentes en el oligoceno tardío y mioceno temprano. Pérez-Contreras et al. (2008) evaluaron las fluctuaciones del área foliar en acículas de pinos como un indicador de resistencia al ataque de Thaumetopoea pityocampa, un lepidóptero defoliador de la península ibérica. Finalmente, Freeman et al. (2005) propusieron la fluctuación de la longitud de los lóbulos foliares en Cnidoscolus stimulosus y el ángulo de apertura de la base foliar en Ipomoea pandurata, como indicadores de la inestabilidad en el desarrollo de estas dos plantas, para identificar el efecto de las perturbaciones edáficas en zonas de pastoreo, en el este de los Estados Unidos.
Los resultados logrados en este estudio serían aplicados para optimizar el manejo de los SAF en los cuales G. arborea es el componente arbóreo, cuya función es generar sombra que estimule los procesos reproductivos de los cultivos agrícolas asociados, tal como es el caso en la presente investigación. Poder identificar los estados tempranos de estrés en G. arborea y corregir el nivel de humedad con estrategias de irrigación evitaría la defoliación de los árboles que es el mecanismo natural de la especie para enfrentar el déficit hídrico en el suelo. De esta manera, se mantendría la condición de sombra durante todo el año y optimizaría los rendimientos productivos de los cultivos. Igualmente, el componente arbóreo dedicaría su mayor esfuerzo al desarrollo y reproducción de los individuos al crecer en un entorno ambiental más estable, al tener garantizada su supervivencia.
CONCLUSIONES
Gmelina arborea es un árbol tropical utilizado ampliamente en plantaciones y sistemas agroforestales (SAF). Para este estudio en particular, sus individuos manifestaron diversidad foliar, la cual se expresa tanto a nivel de la morfometría de la hoja, como en la funcionalidad de esta, lo que le permite adaptarse a diferentes condiciones de humedad del suelo, esto se expresa en un diferencial en la asimetría foliar (ASF), que es un indicador de la inestabilidad en el desarrollo generada principalmente por el estrés hídrico.
Cuando la condición de sequía se incrementa como sucede bajo la oferta ambiental del entorno natural de crecimiento de los bosques secos tropicales, sus árboles generan defoliación total como medida de supervivencia. Si G. arborea es el componente forestal en un SAF, como es el caso de la presente investigación, su defoliación total afecta directamente los procesos reproductivos de los cultivos asociados, tales como T. cacao, al aumentar la radiación y la temperatura sobre el componente agrícola. Por eso, poder utilizar indicadores morfométricos foliares como indicadores tempranos del estrés hídrico para los árboles de G. arborea, tales como el ángulo de inserción del peciolo (AIP) y la razón de dientes foliares (RDF) �que correlacionan altamente con la ASF y además son muy fáciles de identificarlos en campo�, permite implementar estrategias de manejo asociadas al uso del agua en el suelo, para que el componente forestal pueda mantener su dosel en forma permanente, lo cual incrementa los niveles de productividad de los cultivos agrícolas asociados y que los árboles puedan dedicar su energía al crecimiento y desarrollo, al haber garantizado su supervivencia.
AGRADECIMIENTOS
Agradecimientos al Laboratorio de Fisiología Vegetal de la Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Sede Medellín, y al Laboratorio de Ciencias Forestales, Facultad de Ingeniería Forestal, Universidad del Tolima, Ibagué. A la vicerrectoría de investigación y a la Dirección de Investigación - Medellín (DIME), de la Universidad Nacional de Colombia, por la financiación de esta investigación
REFERENCIAS BIBLIOGRÁFICAS
Alados, C.L. (2009). Inestabilidad en el desarrollo y cambio global en plantas. Revista Real Academia de Ciencias. Zaragoza, 64, 35-49.
Alados, C.L., & Aich, A.E. (2008). Stress assessment of argan (Argania spinosa L. Skeels) in response to land uses across an aridity gradient: Translational asymmetry and branch fractal dimension. Journal of Arid Environments, 72, 338-349.
Alados, C.L., Giner, M.L., Dehesa, L., Escós, J., Barroso, F.G., Emlen, J.M., & Freeman, D.C. (2002). Developmental instability and fitness in Periploca laevigata experiencing grazing disturbance. International Journal of Plant Sciences, 163(6), 969-978.
Aldana, J.A. (2008). Evaluación y selección fenotípica de árboles superiores en rodales de Gmelina arborea en el valle geográfico del Rio Magdalena (Tesis de pregrado, Ingeniería Forestal). Ibagué: Universidad del Tolima. 125 p.
Arias, D., Murillo, O., & Rojas, F. (2004). Manual para productores de melina (Gmelina arborea) en Costa Rica. Cartago: Fonafifo. 120 p.
Burrows, G.E. (2001). Comparative anatomy of the photosynthetic organs of 39 xeromorphic species from subhumid New South Wales, Australia. International Journal of Plant Sciences, 162(2), 411-430.
Dale, V.H., & Beyeler, S.C. (2001). Challenges in development and use of ecological indicators. Ecological Indicators, 1, 3-10.
Emlen, J.M., Freeman, D.C., & Graham, J.H. (1993). Nonlinear growth dynamics and the origin of fluctuating asymmetry. Genetica, 89, 77-96.
Fageria, N.K., Baligar, V.C., & Clarck, R.B. (2006). Physiology of crop production. London: FPP. 345 p.
Fair, J.M., & Breshears, D.D. (2005). Drought stress and fluctuating asymmetry in Quercus undulata leaves: confounding effects of absolute and relative amounts of stress? Journal of Arid Environments, 62, 235-249.
Freeman, D.C., Brown, M.L., Duda, J.J., Graham, J.H., Emlen, J.M., Krzysik, A.J., Balbach, H.E., Kovacic, D.A., & Zak, J.C. (2004). Photosynthesis and fluctuating asymmetry as indicators of plant response to soil disturbance in the fall-line sandhills of Georgia: a case study using Rhus copallinum and Ipomoea pandurata. International Journal of Plant Sciences, 165(5), 805-816.
Freeman, D.C., Brown, M.L., Duda, J.J., Graham, J.H., Emlen, J.M., Krzysik, A.J., Balbach, H., Kovacic, D.A., & Zak, J.C. (2005). Leaf fluctuating asymmetry, soil disturbance and plant stress: a multiple year comparison using two herbs, Ipomoea pandurata and Cnidoscolus stimulosus. Ecological Indicators, (5), 85-95.
Kozlov, M.V., Niemela, P., & Malkonen, E. (2002). Effects of compensatory fertilization on pollution-induced stress in Scots pine. Water Air and Soil Pollution, 134, 307-318.
Larcher, W. (2003). Physiological plant ecology. New York: Springer Verlag. 520 p.
Lorenzo, N., Mantuano, D.G., & Mantovani, A. (2010). Comparative leaf ecophysiology and anatomy of seedlings, young and adult individuals of the epiphytic aroid Anthurium scandens (Aubl.) Engl. Environmental and Experimental Botany, 68, 314-322.
Mogie, M., & Cousins, M. (2001). Are sample sizes usually at least an order of magnitude too low for reliable estimates of leaf asymmetry? Journal of Theoretical Biology, 211, 181-185.
Møller, A.P. (2006). A review of developmental instability, parasitism and disease Infection genetics and evolution. Infection, Genetics and Evolution, 6, 133-140.
Páez, Q.E., & Barreiro, H.D. (1986). Análisis del régimen de lluvias de la estación experimental �El Espinal� Centro Cotové. (Trabajo de pregrado, Ingeniería Agrónoma). Universidad Nacional de Colombia, sede Medellín. Medellín: Facultad de Ciencias Agropecuarias. 120 p.
Palmer, A.R., & Strobeck, C. (1986). Fluctuating asymmetry: measurement, analysis, patterns. Annual Review of Ecology and Systematic, 17, 391-421.
Pérez-Contreras, T., Soler, J. J., & Soler, M. (2008). Needle asymmetry, pine vigour and pine selection by the processionary moth Thaumetopoea pityocampa. Acta Oecologica, 33, 213-221.
Rettig, J.E., Fuller, R.C., Corbett, A.L., & Getty, T. (1997). Fluctuating asymmetry indicate levels of competition in an even-aged poplar clone. Oikos, 80, 123-127.
Sack, L., & Frole, K. (2006). Leaf structural diversity is related to hydraulic capacity trees. Ecology, 87(2), 483-491.
Stuckens, J., Verstraeten, W.W., Delalieux, S., Swennen, R., & Coppin, P. (2009). A dorsiventral leaf radiative transfer model: Development, validation and improved model inversion techniques. Remote Sensing of Environment, 113, 2560-2573.
Tamas, J., & Hably, L. (2009). Unidirectional shift in leaf morphology of coexisting species: a possible indicator of palaeoclimatic differences. Palaeogeography, Palaeoclimatology, Palaeoecology, 27, 185-195.
Velickovic, M.V. (2005). Does the coefficient of variation reflect developmental instability in Plants? Archives of Biological Science Belgrade, 57 (3), 7-8.
Villar, R., Ruiz, J., Quero, J.L., Poorter, H., Valladares, F., & Marañón, T. (2004). Tasas de crecimiento en especies leñosas: aspectos funcionales e implicaciones ecológicas. En: Valladares, F. (2004). Ecología del bosque mediterráneo en un mundo cambiante (pp. 191-227). Madrid: Ministerio de Medio Ambiente, EGRAF S.A.




Licencia
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






