
DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2015.1.a06Publicado:
01-01-2015Número:
Vol. 18 Núm. 1 (2015): Enero-JunioSección:
Artículos de investigación científica y tecnológicaDiversidad de árboles y arbustos en fragmentos de bosque seco tropical en río Hato, Panamá
Diversity of trees and shrubs in tropical dry forest fragments in Río Hato, Panama
Palabras clave:
dry arch of Panama, tropical dry forests, Garcinia madruno, Sloanea terniflora (en).Palabras clave:
arco seco de Panamá, bosque seco tropical, Garcinia madruno, Sloanea terniflora (es).Descargas
Referencias
Bawa, K.S. (1990). Plant-pollinator interactions in tropical rain forests. Annual Review of Ecology and Systematics, 21, 399-422.
Cascante, A., Quesada, M., Lobo, J.J, & Fuchs, E.A. (2002). Effects of dry tropical forests fragmentation on the reproductive success and genetic structure of the tree Samanea saman. Conservation Biology, 16(1), 137-147.
Colwell, R.K. (2009). Biodiversity: concepts, patterns, and measurement. En S.A. Levin, S.R., Carpenter, H.J., Charles, A.P., Godfray, Kinzig, M., Loreau, J.B., Losos, B., Walker & D.S. Wilcove (eds.). The Princeton Guide to Ecology(pp. 257-263). Princeton. 848 p.
Colwell, R.K. (2013). EstimateS: Statistical estimation of species richness and shared species from samples. Version 9 and earlier. User’s Guide and application. Published at http://purl.oclc.org/estimates
Cooke, R.G., & Ranere, A.J. (1992). Prehistoric human adaptations to the seasonally dry forests of Panama. World Archaeology, 24, 114-133.
D’Antonio, C.M., & Vitousek, P.M. (1992). Biological invasions by exotic grasses, the grass-fire cycle, and global change. Annual Review of Ecology and Systematics, 23, 63-87.
García-Villacorta, R. (2009). Diversidad, composición y estructura de una hábitat altamente amenazado: los bosques estacionalmente secos de Tarapoto, Perú. Revista Peruana de Biología, 16(1). Recuperado de http://www.scielo.org.pe/scielo.php?script=sci_arttext&pid=S1727-99332009000100010&lng=es&nrm=iso>
Graves, S. (2014). Contribution of tropical agricultural trees to species diversity and carbon: a new landscape perspective enabled by high- resolution hyperspectral and lidar images (Tesis de Maestría). Gainsville: University of Florida. 84 p.
Guariguata, R.M. (1998). Consideraciones ecológicas sobre la regeneración natural aplicada al manejo forestal. Colección Manejo Diversificado de Bosques naturales. Publicación No. 14. Costa Rica-Turrialba: CATIE. 27 p.
Heckadon-Moreno, S. (2007). Cuando se acaban los montes. Ciudad de Panamá: Editorial Universitaria- Instituto Smithsonian de Investigaciones Tropicales. 252 p.
Heckadon-Moreno, S. (2009). De selvas a potreros. Ciudad de Panamá: Editorial Exedra. 300 p.
Holdridge, L.R. (1967). Life zone ecology. San José de Costa Rica: Tropical Science Center. 140 p.
Instituto Nacional de Estadística y Censo. (2010). Contraloría Nacional de la República de Panamá. Sección Clima (121-Meteorología).Recuperado de http://www.contraloria.gob.pa/INEC/
Janzen, D.H. (1988). Tropical dry forests: the most endangered major tropical ecosystem. En E.O. Wilson (ed.). Biodiversity (pp. 130-137). Washington, D.C.: National Academy Press.
Linares, P.R., & Martínez, H. (1991). La regeneración natural temprana del bosque de cativo en chocó, Colombia. Bogotá: Corporación Nacional de Investigación y Fomento Forestal (CONIF). pp. 1-27.
López, O.R. (2012). The success of introduced alien plant species in the tropics: the Panama case. The Open Ecology Journal, 5, 84-89.
Lezcano, J., & López, O.R. (2014). Historia natural de la vegetación costero-litoral del istmo de Panamá. En F. Rodríguez & A. O’Dea (eds.). Historia Natural del Istmo de Panamá. ISBN 978-9962-05-682-9, en prensa.
Marín, W.A., & Flores, E.M. (2002). Copaifera aromática Dwyer. En J. A. Vozzo (ed.). Tropical tree seed manual (pp. 405-407). Washington D.C: Department of Agriculture, Forest Service.
Marulanda, L.O., Uribe, A., Velásquez, P., Montoya, M.A., Idarrága, A., López, M.C., & López, J.M. (2003). Estructura y composición de la vegetación de un fragmento de bosque seco en San Sebastián, Magdalena (Colombia). I. Composición de plantas vasculares. Actualidad Biológica, 25(78), 17-30.
Mendoza, C.H. (1999). Estructura y riqueza florística del bosque seco tropical en la región Caribe y el valle del río Magdalena, Colombia. Caldasia, 21, 70-94.
Murphy, P.G., & Lugo, A.E. (1986). Ecology of tropical dry forest. Annals Review of Ecology and Systematics, 17, 67-68.
Perez, R., & Condit, R. (2014). Tree Atlas of Panama. Ciudad de Panamá: Center for Tropical Forest Science, Smithsonian Tropical Research Institute. Recuperado de [http://ctfs.arnarb.harvard.edu/webatlas/maintreeatlas.php]
Portillo-Quintero, C.A., & Sánchez-Azofeifa, G.A. (2010). Extent and conservation of tropical dry forests in the Americas. Biological Conservation, 143(1), 144-155.
Quesada, M., Sanchez-Azofeifa, G.A., Alvarez, M., Stoner, K., Avila-Cabadilla, L.D., Calvo-Alvarado, J.C., Castillo, A., Espírito-Santo, M.M., Fagundes, M., Fernandes, G.W., Gamon, J., Lopezaraiza-Mikel, M., Lawrence, D., Morellato, L.P.C., Powers, J., Neves, F.S., Rosas-Guerrero, V., Sayago, R., & Sanchez-Montoya, G. (2009). Succession and management of tropical dry forests in the Americas: Review and New Perspectives. Forest Ecology and Management, 258, 1014-1024.
Rodríguez M, G.M., Banda-R, K., Reyes B, S.P., & Estupiñán González, A.C. (2013). Lista comentada de las plantas vasculares de bosques secos prioritarios para la conservación en los departamentos de Atlántico y Bolívar (Caribe colombiano). Biota Colombiana, 13(2), 7-39.
Sánchez-Azofeifa, G.A., Kalacska, M. Quesada, M., Calvo, J., Nassar, J., & Rodríguez, J.P. (2005). Need for integrated research for a sustainable future in tropical dry forests. Conservation Biology, 19(2), 1285-286.
Vieira, D.L.M., & Scariot, A. (2006). Principles of natural regeneration of tropical dry forests for restoration. Restoration Ecology, 14(1), 11-20.
Wang H., Sork, V.L., Wua, J., & Ge, J. (2010). Effect of patch size and isolation on mating patterns and seed production in an urban population of Chinese pine (Pinus tabulaeformis Carr.). Forest Ecology and Management, 260, 965-974.
Zamora, N. (1993). Flora arborescente de Costa Rica. Cartago: Editorial Tecnológica. 262 p.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
 |
López, O., Pérez, R. & Mariscal, E.(2015). Diversidad de árboles y arbustos en fragmentos de bosque seco tropical en Rio Dato, pacífico de Panamá. Colombia Forestal, 18(1), 105-115 |
Artículo de investigación
DIVERSIDAD DE ÁRBOLES Y ARBUSTOS EN FRAGMENTOS DE BOSQUE SECO TROPICAL EN RÍO HATO, PANAMÁ
Diversity of trees and shrubs in tropical dry forest fragments in Río Hato, Panama
Omar R. López1, Rolando Pérez2, Emilio Mariscal3
1 Centro de Biodiversidad y Descubrimiento de Drogas, Instituto de Investigaciones Científicas y Servicios de Alta Tecnología. Apartado Postal 0843-01103, Ciudad del Saber, Clayton. Panamá, República de Panamá.prioria@yahoo.com Autor para correspondencia.
2 Smithsonian Tropical Research Institute. Apartado Postal 0843-03092, Balboa, Ancón, Panamá, República de Panamá
3 Departamento Forestal. Autoridad Nacional del Ambiente. Albrook, Edificio 804, Apartado Postal 0843-00793, Balboa, Ancón Panamá, República de Panamá.
Recepción: 23 de octubre de 2014 / Aprobación: 9 de diciembre de 2014
RESUMEN
En fragmentos de bosque seco tropical en Río Hato, Panamá, estudiamos la composición florística según los árboles y arbustos. Establecimos 61 parcelas de 100 m2 distribuidas de forma aleatoria, totalizando 0.61 ha. En cada unidad de muestreo medimos todos los individuos presentes con un DAP >5.0 cm. Encontramos un total de 52 especies de árboles representadas en veinticinco familias, de las cuales Fabaceae, Burseraceae, Myrtaceae y Rubiaceae aportan mayor número de especies (~35%). La especie Sloanea terniflora se encontró en más del 60.6% de los cuadrantes estudiados y resultó además ser la especie con mayor dominancia relativa (23%), seguida de Anacardium excelsum, que con solo quince individuos de grandes diámetros presentó una dominancia relativa del 10%. La diversidad promedio reveló índices intermedios (3.13 índice de Shannon). A pesar del bajo número de especies en comparación con otros bs-T de la región, los fragmentos en Río Hato conservan elementos florísticos de importancia ecológica y para la conservación de los bosques secos. Garcinia madruno, una de las especies más abundante es clave en la producción de frutos para la fauna. Otras especies de importancia son Manilkara sapota y Copaifera aromatica, las cuales aún son utilizadas como recurso maderero. Es prioritario para Panamá establecer estrategias de conservación que salvaguarden estos fragmentos como fuente de especies del bosque seco tropical y refugio para la vida silvestre.
Palabras clave: arco seco de Panamá, bosque seco tropical, Garcinia madruno, Sloanea terniflora.
ABSTRACT
The floristic composition of trees and shrubs in tropical dry forest fragments in Río Hato, Panama was studied. We established 61 plots of 100 m2 distributed randomly, totaling 0.61 hectares. In each sampling unit we measured all individuals above 5.0 cm DAP. We found a total of 52 tree species represented in 25 families, of which Fabaceae, Burseraceae, Myrtaceae and Rubiaceae account for 35% of the species. The species Sloanea terniflora was found in more than 60.6% of the quadrants. This species also proved to be the one with the highest relative dominance (23%) followed by Anacardium excelsum, with only 15 individuals of large diameters, showing a relative dominance of 10%. Mean diversity indices revealed intermediate values (Shannon index 3.13). While the number of tree species is relatively low when compared to other tropical dry forests in the region, the fragments in Río Hato preserve floristic elements of ecological importance for the conservation of tropical dry forests. Garcinia madruno, the most abundant species is a key species in fruit production for wildlife and other species have been overexploited as timber resources, e.g., Manilkara sapota and Copaifera aromatica. Establishing conservation strategies to safeguard these fragments as a source of tropical dry forest species and as wildlife refuges should be a priority for Panama.
Key words: dry arch of Panama, tropical dry forests, Garcinia madruno, Sloanea terniflora.
INTRODUCCIÓN
El bosque seco tropical (bs-T) representa la formación tropical más amenazada del planeta. En el neotrópico centroamericano el bs-T ha sido el ecosistema predilecto para el desarrollo de asentamientos humanos proporcionando una larga historia de uso y trasformación (últimos diez mil años; Cooke & Ranere, 1992). Actualmente, las actividades humanas continúan alterando y diezmando la extensión del bs-T en Centroamérica, aun cuando desconocemos bastante en lo que respecta a su diversidad y los procesos ecológicos que lo regulan (Vieira & Scariot, 2006; Quezada et al., 2009). Lo anterior se da principalmente por la conversión del bs-T a pastizales para la ganadería y agricultura extensiva (Heckadon, 2007; 2009), la introducción y establecimiento exitoso de especies alóctonas mejores adaptadas a la perturbación (e.g., fuego; D'Antonio & Vistousek, 1992; López, 2012), así como el desarrollo inmobiliario y turístico en áreas costeras (Lezcano & López, 2014). Por consiguiente, es prioritaria la conservación y regeneración del bs-T en la región ya que solo cerca del 1% de bs-T se encuentra protegido (Sanchez-Azofeifa et al., 2005; Janzen, 1988).
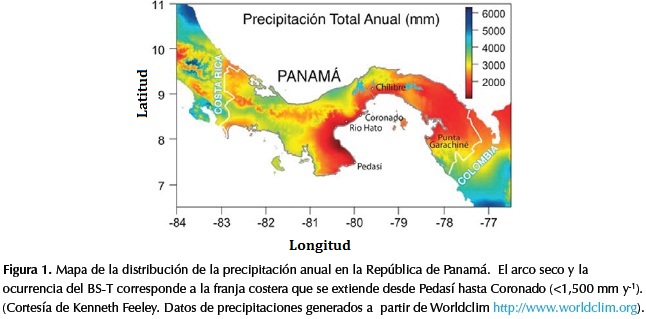
En Panamá los bosques secos se encuentran asociados al litoral Pacífico, específicamente en lo que se conoce como el "arco seco" de Panamá, que es una franja costera que se ubica allí, el cual recibe una precipitación menor a los 1500mm al año y que se extiende desde el poblado de Pedasí, en la costa sureste de la península de Azuero hasta Coronado, en la provincia de Panamá (Figura 1). Otras áreas de bs-T son observables sobre formaciones geológicas calcáreas en el área central de Panamá (Chagres) y en Punta Garachiné (Figura 1). Aun cuando Panamá cuenta con una extensión de 2 128 km2 de bosques secos (2.7% del territorio), el bs-T se encuentra débilmente protegido (Portillo & Sánchez, 2009). El Parque Nacional Sarigua, que abarca unos 8 km2, es el único parque nacional que contempla la formación de bs-T, pero que en su mayoría comprende áreas de marismas con suelos altamente degradados y remanentes de bs-T entremezclados. Otros remanentes de bs-T, como lo es el bosque de El Colmón (una de las primeras áreas protegidas del continente decretada como bosque comunal en 1918, así como lo es el refugio de vida silvestre Pablo Arturo Barrios en la provincia de Los Santos), no cuentan con el nivel de respaldo gubernamental que garanticen su protección y conservación.
El presente trabajo busca fortalecer el conocimiento acerca del bosque tropical seco en la región centroamericana y en particular en Panamá, donde se cuenta con pocos estudios de este tipo de bosque. Presentamos el resultado de un inventario florístico realizado en fragmentos de bs-T en el área de Río Hato, en el Pacífico panameño. Se analiza la diversidad, estructura y similitud de la flora en dichos fragmentos y se destaca el valor de dichos fragmentos en la conservación del bs-T en Panamá.
MATERIALES Y MÉTODOS
Área de estudio
Los fragmentos de bs-T de Río Hato se encuentran ubicados en la provincia de Coclé (8°25'42" N y 80°07'17" W; Figura 1). El área se halla dentro de la zona de vida de bosque seco tropical (bs-T) según Holdridge (1967) y abarca una superficie total de aproximadamente veinticuatro ha con una área efectiva de bosque de 10.5 ha. Los fragmentos de estudio representan remanentes de bs-T que han sufrido cierto grado de perturbación ya que buena parte del área fue utilizada para el pastoreo y sometida a fuegos recurrentes. La precipitación promedio anual es de 1 280.6 mm (n=15 años; Instituto Nacional de Estadística y Censo, 2010). Muestras de suelo tomadas en el interior del bosque indican que este se caracteriza por presentar una textura de tipo Arenoso Franco (AF) y Franco-Arcilloso (FA), el pH promedio es de 6.0, su coloración típica es pardo, con ligeras variaciones entre pardo-pardo pálido y pardo grisáceo
Metodología de los inventarios
Se establecieron 61 parcelas de 100 m2 de forma aleatoria dentro de cinco parches de bosques, totalizando 0.61 ha. Se levantó información florística de todos los individuos mayores de 5 cm de diámetro a la altura del pecho (DAP). Se realizaron identificaciones in situ de las especies encontradas y registraron los nombres comunes empleados por los campesinos de la región. Todas las identificaciones en campo fueron realizadas por R.P., botánico con más de 30 años de experiencia (Pérez & Condit, 2014). En el caso de desconocidos (dos en nuestro caso) se tomó al menos tres muestras por cada espécimen, se anotó la localidad, las coordenadas geográficas empleando un GPSmap 60CSx modelo GARMIN. Los espécimenes fueron recolectados (preferiblemente con flores y frutos), prensados y secados en un horno a una temperatura promedio de 46°C por un período de tres a cinco días. Seguidamente se identificó el material empleando para ello las claves taxonómicas publicadas en los tratados de: Flora de Panamá, Flora Neotrópica, Flora de Nicaragua, Flora Mesoamericana, además de libros y artículos de diferentes revistas referentes a la revisión de las familias y los géneros. Las muestras de los especímenes se entregaron en el Herbario Nacional ubicado en la Universidad de Panamá (PMA).
Análisis
En primer lugar evaluamos el esfuerzo del muestreo sobre la base del comportamiento de la relación especies-área muestreada (cuadrantes) utilizando el número de especies observadas (Sobs) y estimadas (Sest) y el estimador de riqueza de especies basado en cobertura, Chao1. Adicionalmente, estimamos los índices de diversidad de Shannon y Sørensen. En dichos análisis utilizamos el programa EstimateS 9.1.0 (Colwell, 2013). Para los análisis estructurales utilizamos los descriptores más comúnmente mencionados en la literatura con base en las medidas de diámetro a partir de 5 cm de DAP. Para cada especie estimamos la abundancia relativa (AR, %) definida como el número de individuos de una especie en proporción al número total de individuos de toda la muestra. De manera similar, calculamos la dominancia relativa (DR, %) como el área basal de una especie en proporción al área basal total y la frecuencia relativa (FR, %) como el número de cuadrantes en que una especie se encuentra presente a razón del total de cuadrantes en toda la muestra.
RESULTADOS
Esfuerzo del muestreo
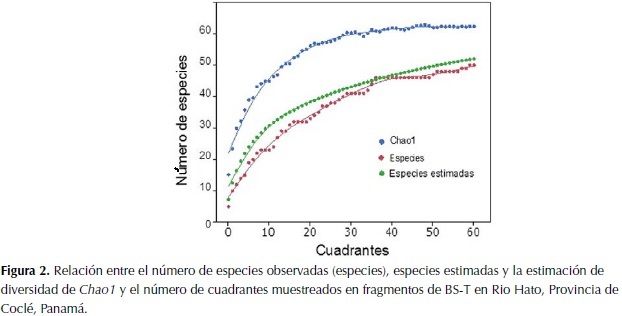
La Figura 2 presenta la relación entre Sobs, Sest y Chao1 y el número de cuadrantes muestreados. Cabe notar que la estimación del índice de diversidad de Chao1 es sensible al número de especies raras encontradas en el muestreo (especies que solo cuentan con un individuo "singletons" o dos individuos en toda la muestra "doubletons") (Colwell, 2009). En nuestro muestreo encontramos un total de once especies que fueron registradas una sola vez, y siete especies de las que se encontraron solo dos individuos, lo que representa cerca de un 35% de especies raras en la muestra.
Composición florística y estructural
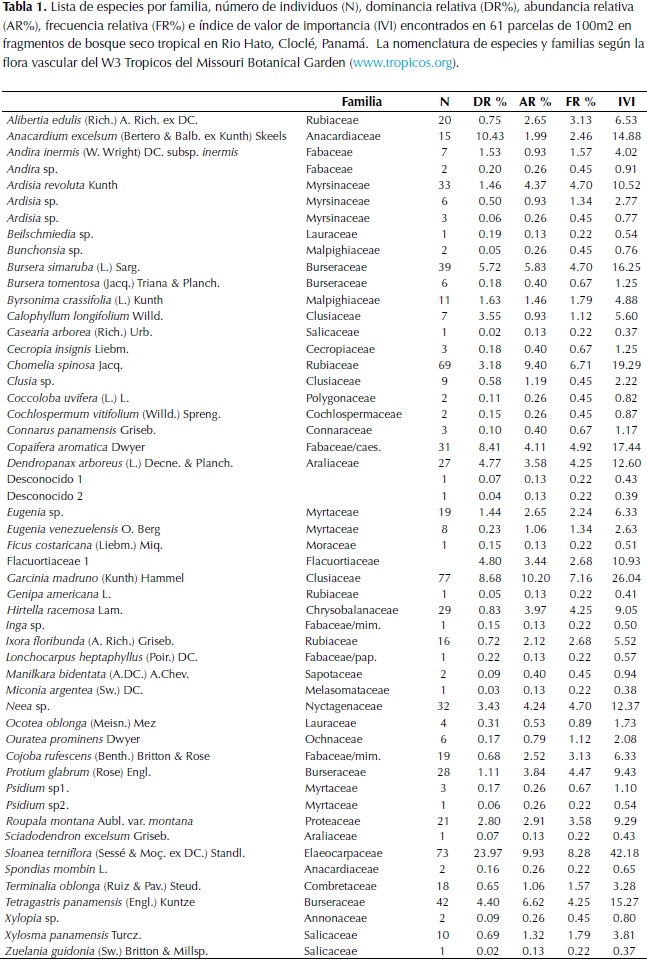
En totalse registraron 52 especies distribuidas en 25 familias. Tomando en cuenta el tamaño de la unidad de muestreo (0.001 ha), en promedio se encontraron siete especies por parcela con un rango entre dos y dieciséis especies por unidad de muestreo. Las Familias Fabaceae, Burseraceae, Myrtaceae y Rubiaceae fueron las más numerosas conteniendo 35% de todas las especies (Tabla 1). En general, Garcinia madruno (Kunth) Hammel, fue la especie más numerosa con una AR de 10.2 %, mientras que Sloanea terniflora (Sessé & MoÃ�. ex DC.) Standl. la más frecuente (FR), presente en más de 60.6% de los cuadrantes estudiados. No obstante, S. terniflora resultó ser la especie más dominante con 23% de DR, seguida de Anacardium excelsum (Bertero & Balb. ex Kunth) Skeels, que con tan solo quince individuos (AR del 2%), presentó una DR del 10%. Ambas especies registraron varios individuos con DAP superior a los 40 cm, lo que contribuye sustancialmente a su dominancia con base en el área basal. Como resultado de la alta frecuencia y dominancia con que S. terniflora se mostró en los fragmentos muestreados, esta especie resultó ser la especie con el mayor índice de valor de importancia (IVI) seguida de G. madruno.
En el Anexo 1 se presentan los valores de abundancia de individuos por especie >5 cm DAP para cada una de las 61 parcelas de estudio. En promedio, el índice de similitud de Sørenesen mostró un valor relativamente bajo de 0.27 (donde 1 indica todas las especies compartidas), ya que cerca del 50% de las parcelas no comparten una sola especie (Anexo 1). Adicionalmente, dado el alto número de especies raras y el tamaño de la muestra es posible que la similitud entre las parcelas muestreadas sea subestimado. El índice de Shannon mostró un valor promedio de diversidad de 3.13, una diversidad relativamente mediana en comparación con bosques húmedos, a pesar de la falta de equidad en el área muestreada. El índice de diversidad de Fisher alfa se mostró bastante robusto a través de los diferentes cuadrantes muestreados con un valor promedio de 12.6.
DISCUSIÓN
En general, el número de especies encontradas en los fragmentos del bs-T de Río Hato, unas 52 especies agrupadas en 44 géneros y 25 familias en 0.61 ha, es bajo en comparación con otros bosques secos de la región. Por ejemplo, Mendoza (1999), considerando los individuos >2.5 cm de DAP, reporta un promedio de 60 especies por 0.1 ha en los bosques secos de la región del Caribe y el valle seco del río Magdalena, Colombia. De manera similar, el número de especies encontradas en Río Hato es inferior al de otros bosques secos de la región (p.ej., Costa Rica y Venezuela, reportados en Mendoza, 1999). No obstante, el número de especies y estructura de los fragmentos de bs-T de Río Hato, Panamá, es relevante dentro del contexto de perturbación, principalmente fuego y sobrepastoreo, que los bs-T en la costa del Pacífico panameño experimentan. A nivel de familia, la familia Fabaceae fue la que más contribuyó en especies. Este resultado es consistente con la mayoría de inventarios en la región (Mendoza, 1999; Marulanda et al., 2003; Rodríguez et al., 2013). Sin embargo, Río Hato difiere de muchos otros bs-T de la región en donde no se encontraron especies de la familia Bignonaceae y Capparidaceae, familias típicas de bs-T (Murphy & Lugo, 1986).
En términos de estructura, S. terniflora mostró ser la especie de mayor importancia según el IVI. G. madruno, localmente conocido como "satro", con 77 individuos, es una de las especies de importancia ecológica en estros fragmentos dado que sus frutos son consumidos por diversos elementos de la fauna (E.M. observación personal). Por otro lado, aunque en menor proporción fueron registradas especies maderable tales como; Copaifera aromatica ("cabimo"), A. excelsum ("espavé"), Calophyllum longifolium ("María") y la especie Manilkara bidentata ("níspero"). No obstante la ocurrencia de estas especies, en particular C. aromatica y A. excelsum, podría estar ligada a factores edafo-climáticos del sitio, ya que Cabimo es una especie característica de bosques más húmedos (Marín & Flores, 2002), al igual que Espavé, una especie que aunque con amplia distribución, es frecuente en bosques de galería y dominante en planicies inundables (Zamora, 1993; Linares & Martínez, 1991). En efecto, es posible que estas especies sean más frecuentes en unidades de muestreo con suelos más húmedos.
En comparación con otros bs-T de la región, los fragmentos de Río Hato muestran valores de diversidad inferiores. García-Villacorta (2009) reporta índices de Fisher's alfa que varían entre 14.1 y 21.6 para 10 parcelas de 0.1 ha en el bs-T de Tarapoto, Perú (vs. 12.6 para Río Hato). De igual manera, Marulanda et al. (2003) registraron 155 especies utilizando la misma unidad de muestreo (100 m2) que en nuestro estudio. No obstante, en ambos casos el DAP mínimo fue de 2.5 cm, lo que podría explicar la mayor diversidad en relación con nuestro sitio de estudio en Río Hato, Panamá. La disparidad de áreas muestreadas y diámetros mínimos empleados dificulta establecer comparaciones objetivas entre distintos bosques secos tropicales en la región. La unificación de criterios y mínimo de área muestreada, así como también la posibilidad de cuantificar la dinámica de estos bosques a largo plazo podría contribuir al establecimiento de mejores estrategias de conservación e identificación de especies en peligro.
El fraccionamiento, la sobreexplotación y complejos sistemas reproductivos podrían diezmar significativamente la capacidad regenerativa de algunas especies del bs-T. El distanciamiento entre individuos reproductivos de una especie con síndromes de polinizacion auto-incompatibles sufren desproporcionadamente una reducción en su capacidad reproductiva (Bawa, 1990; Guariguata, 1998). A nivel de paisaje, el fraccionamiento, como es el caso de los remanentes de bs-T de la costa Pacífica de Centroamérica, estaría incrementando las distancias de flujo de material genético y las tasas de predación de las semillas producidas por individuos aislados. Cascante et al. (2002) han demostrado cómo la fragmentacion y el aislamiento (>500 m) afecta el porcentaje de germinación y el vigor de las semillas de Samanea saman (Jacq.) Merrill, una especie común del bs-T. Aunado a estas limitaciones reproductivas, la tala selectiva reduce la producción de semillas tanto a nivel de individuos, así como también las distancias genéticas de potenciales "donantes" de polen más cercanos. Lo anterior podría ser el caso de M. bidentata, una especie maderable y con frutos atractivos para la fauna, que en condiciones de aislamiento tienden a producir frutos sin semillas (ORL observación personal; Wang et al., 2010).
CONCLUSIONES
Desde una perspectiva de conservación el bs-T requiere la conformación de áreas de conservación funcionales que salvaguarden los elementos propios de este tipo de bosque tropical en Panamá. A pesar de su alto grado de fraccionamiento, este estudio evidencia que los fragmentos en Río Hato aún conservan una diversidad importante de especies de valor ecológico del bs-T. En otros casos, por ejemplo M. bidentata "Níspero", de la cual se encontraron pocos individuos, es imperante evaluar su vulnerabilidad genética y reproductiva a manera de establecer estrategias de conservación para esta especie. Es importante además considerar estos fragmentos de bosque como parte de las estrategia de mitigación al cambio climático (e.g. Programa REDD de Naciones Unidas), ya que especies con grandes volúmenes (p.ej., A. excelsum y C. aromática) podrían jugar un papel importante en el secuestro de carbono a escala de paisaje (Graves, 2014). Se requiere ampliar los inventarios florísticos y estudios genéticos en remanentes y áreas continuas de bs-T en Panamá con el propósito de determinar las especies más vulnerables al fraccionamiento y al aislamiento reproductivo.
REFERENCIAS BIBLIOGRÁFICAS
Bawa, K.S. (1990). Plant-pollinator interactions in tropical rain forests. Annual Review of Ecology and Systematics, 21, 399-422.
Cascante, A., Quesada, M., Lobo, J.J, & Fuchs, E.A. (2002). Effects of dry tropical forests fragmentation on the reproductive success and genetic structure of the tree Samanea saman. Conservation Biology, 16(1), 137-147.
Colwell, R.K. (2009). Biodiversity: concepts, patterns, and measurement. En S.A. Levin, S.R., Carpenter, H.J., Charles, A.P., Godfray, Kinzig, M., Loreau, J.B., Losos, B., Walker & D.S. Wilcove (eds.). The Princeton Guide to Ecology(pp. 257-263). Princeton. 848 p.
Colwell, R.K. (2013). EstimateS: Statistical estimation of species richness and shared species from samples. Version 9 and earlier. User's Guide and application. Published at http://purl.oclc.org/estimates
Cooke, R.G., & Ranere, A.J. (1992). Prehistoric human adaptations to the seasonally dry forests of Panama. World Archaeology,24, 114-133.
D'Antonio, C.M., & Vitousek, P.M. (1992). Biological invasions by exotic grasses, the grass-fire cycle, and global change. Annual Review of Ecology and Systematics, 23, 63-87.
García-Villacorta, R. (2009). Diversidad, composición y estructura de una hábitat altamente amenazado: los bosques estacionalmente secos de Tarapoto, Perú. Revista Peruana de Biología, 16(1). Recuperado de http://www.scielo.org.pe/scielo.php?script=sci_arttext&pid=S1727-99332009000100010&lng=es&nrm=iso
Graves, S. (2014). Contribution of tropical agricultural trees to species diversity and carbon: a new landscape perspective enabled by high- resolution hyperspectral and lidar images (Tesis de Maestría). Gainsville: University of Florida. 84 p.
Guariguata, R.M. (1998). Consideraciones ecológicas sobre la regeneración natural aplicada al manejo forestal. Colección Manejo Diversificado de Bosques naturales. Publicación No. 14. Costa Rica-Turrialba: CATIE. 27 p.
Heckadon-Moreno, S. (2007). Cuando se acaban los montes. Ciudad de Panamá: Editorial Universitaria- Instituto Smithsonian de Investigaciones Tropicales. 252 p.
Heckadon-Moreno, S. (2009). De selvas a potreros. Ciudad de Panamá: Editorial Exedra. 300 p.
Holdridge, L.R. (1967). Life zone ecology. San José de Costa Rica: Tropical Science Center. 140 p.
Instituto Nacional de Estadística y Censo. (2010). Contraloría Nacional de la República de Panamá. Sección Clima (121-Meteorología).Recuperado de http://www.contraloria.gob.pa/INEC/
Janzen, D.H. (1988). Tropical dry forests: the most endangered major tropical ecosystem. En E.O. Wilson (ed.). Biodiversity (pp. 130-137). Washington, D.C.: National Academy Press.
Linares, P.R., & Martínez, H. (1991). La regeneración natural temprana del bosque de cativo en chocó, Colombia. Bogotá: Corporación Nacional de Investigación y Fomento Forestal (CONIF). pp. 1-27.
López, O.R. (2012). The success of introduced alien plant species in the tropics: the Panama case. The Open Ecology Journal, 5, 84-89.
Lezcano, J., & López, O.R. (2014). Historia natural de la vegetación costero-litoral del istmo de Panamá. En F. Rodríguez & A. O'Dea (eds.). Historia Natural del Istmo de Panamá. ISBN 978-9962-05-682-9, en prensa.
Marulanda, L.O., Uribe, A., Velásquez, P., Montoya, M.A., Idarrága, A., López, M.C., & López, J.M. (2003). Estructura y composición de la vegetación de un fragmento de bosque seco en San Sebastián, Magdalena (Colombia). I. Composición de plantas vasculares. Actualidad Biológica, 25(78), 17-30.
Mendoza, C.H. (1999). Estructura y riqueza florística del bosque seco tropical en la región Caribe y el valle del río Magdalena, Colombia. Caldasia,21, 70-94.
Murphy, P.G., & Lugo, A.E. (1986). Ecology of tropical dry forest. Annals Review of Ecology and Systematics,17, 67-68.
Perez, R., & Condit, R. (2014). Tree Atlas of Panama. Ciudad de Panamá: Center for Tropical Forest Science, Smithsonian Tropical Research Institute. Recuperado de http://ctfs.arnarb.harvard.edu/webatlas/maintreeatlas.php
Portillo-Quintero, C.A., & Sánchez-Azofeifa, G.A. (2010). Extent and conservation of tropical dry forests in the Americas. Biological Conservation, 143(1), 144-155.
Quesada, M., Sanchez-Azofeifa, G.A., Alvarez, M., Stoner, K., Avila-Cabadilla, L.D., Calvo-Alvarado, J.C., Castillo, A., Espírito-Santo, M.M., Fagundes, M., Fernandes, G.W., Gamon, J., Lópezaraiza-Mikel, M., Lawrence, D., Morellato, L.P.C., Powers, J., Neves, F.S., Rosas-Guerrero, V., Sayago, R., & Sanchez-Montoya, G. (2009). Succession and management of tropical dry forests in the Americas: Review and New Perspectives. Forest Ecology and Management, 258, 1014-1024.
Rodríguez M, G.M., Banda-R, K., Reyes B, S.P., & Estupiñán González, A.C. (2013). Lista comentada de las plantas vasculares de bosques secos prioritarios para la conservación en los departamentos de Atlántico y Bolívar (Caribe colombiano). Biota Colombiana, 13(2), 7-39.
Sánchez-Azofeifa, G.A., Kalacska, M. Quesada, M., Calvo, J., Nassar, J., & Rodríguez, J.P. (2005). Need for integrated research for a sustainable future in tropical dry forests. Conservation Biology, 19(2), 1285-286.
Vieira, D.L.M., & Scariot, A. (2006). Principles of natural regeneration of tropical dry forests for restoration. Restoration Ecology, 14(1), 11-20.
Wang H., Sork, V.L., Wua, J., & Ge, J. (2010). Effect of patch size and isolation on mating patterns and seed production in an urban population of Chinese pine (Pinus tabulaeformis Carr.). Forest Ecology and Management, 260, 965-974.
Zamora, N. (1993). Flora arborescente de Costa Rica. Cartago: Editorial Tecnológica. 262 p.




Licencia
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






