
DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2016.1.a05Publicado:
01-01-2016Número:
Vol. 19 Núm. 1 (2016): Enero-JunioSección:
Artículos de investigación científica y tecnológicaEnraizamiento in vitro de embriones cigóticos de Acrocomia aculeata (Jacq.) Lodd ex Mart
Rooting in vitro of zygotic embryos of Acrocomia aculeata (Jacq.) Lodd ex Mart.
Palabras clave:
benzylaminopurine, oily crop, in vitro rooting, latency (en).Palabras clave:
Cultivo oleaginoso, desarrollo de plántulas, enraizamiento in vitro, latencia (es).Descargas
Referencias
Azcón-Bieto, J., & Talón, M. (ed.) (2000). Fundamentos de Fisiología Vegetal. Madrid: McGraw-Hill. Interamericana. 522 p.
Azofeifa, A. (2009). Problemas de oxidación y oscurecimiento de explantes cultivados in vitro. Agronomía Mesoamericana, 20(1), 153–175.
Besnier, R. (1989). Semillas: Biología y tecnología. Madrid: Ediciones Mundi-Prensa. 637 p.
Bidwell, R, G. (1993). Fisiología vegetal. México D. F: A. G. T Editor S. A. 784 p.
Bohn, E. (2009). ‘’Tablero de comando’’ para la promoción de los biocombustibles en Paraguay. Publicación de la Naciones Unidas. CEPAL – Colección de documentos y proyectos. Bundesministerium für wirtschaftliche Zusammenar beit und Entwicklung.GTZ. Santiago de Chile. 112 p.
Borges, G, M., Destrade, B, R., Meneses, R, S., Gómez, K, R., Malaurie, B., Hamon, P., &Demenorval, L, C. (2011).Optimización de un medio de cultivo para plantas micropropagadas de Dioscoreaalata L. Revista Colombiana de Biotecnología, 13(2), 221–228.
Cáceres, A, K. (2004). Propagación in vitro de los porta injertos de cerezo (Prunus avium L.) GISELA 5 Y Prunus cerasus. Universidad Católica de Valparaíso. (Tesis de posgrado, Ciencias en Agronomía). Valparaíso, 69 p.
Centro Internacional de Agricultura Tropical -CIAT-. (1991). Cultivo de tejidos en la agricultura: fundamentos y aplicaciones. Cali: Roca, W, M., Mroginski, L, A. 970 p.
Chorfi, B, L. (2013). Avaliação de populações naturais, estimativas de parâmetros genéticos e seleção de genótipos elite de macaúba (Acrocomia aculeata). (Tesis de posgrado, Agricultura Tropical y Subtropical, Genética, Mejoramiento y Biotecnología Vegetal). Campinas: Instituto Agronómico. 150 p.
Díaz de la Guardia, M. (2010). Fisiología de las plantas. 2da ed. Granada: Grupo Editorial Universitario. 278 p.
Domínguez, R., Alpuche, S, A., Vasco. M, N., & Pérez, M, B. (2008). Efecto de citocininas en la propagación in vitro de Agaves mexicanos. Rev. Fitotec. Mex., 31(4), 317–322.
Esau, K. (1985). Anatomía vegetal. 3ra ed. Barcelona: Ediciones Omega. 779 p.
Ferreira, M, E. (2007). Embriogénese somática de macauba: induçao, regeneraçao e caracterização anatômica. (Trabajo de posgrado, Genética y Mejoramiento). Minas Gerais: Universidade Federal de Viçosa. 66 p.
Francescangeli, N &Zagabria, A. (2012). Citoquininas para modificar laarquitectura de planta de petunia. Información Técnica Económica Agraria, 106 (1), 46–52.
Ganduglia, F., León, J, G., Gasparini, R., Rodríguez, M, E.,Huarte, G, J., Estrada, J., &Filgueiras, E. (2009). Manual de Biocombustibles. San José: IICA 206 p.
González, D, A. (2010). Efectos del Carbón Activado (CA) y el Ácido ascórbico en condiciones de luz y oscuridad sobre la oxidación de embriones zigóticos del mbokaja (Acrocomia aculeata) (Jacq.) en la germinación in vitro. (Trabajo de pregrado, Ingeniería Agronómica). Asunción: Universidad Nacional de Asunción. 91 p.
Hammer, Ø., Harper, D. A. T., Ryan, P. D. (2001). PAST: Paleontological Statistics software package for education and data analysis. Paleontología Electrónica, 4 (1), 9.
Instituto Interamericano de Cooperación Asociación Regional de Empresas de Petróleo y para la Agricultura–IICA-. (2007). El estado del arte de los biocombustibles en el Paraguay. Asunción: Editorial Grafir S.A. 83 p.
Jordan, M., &Casaretto, J. (2006).Hormonas y Reguladores del Crecimiento: Auxinas, Giberelinas y Citocininas. En Squeo, F, A., & Cardemil, L. (eds.). Fisiología Vegetal (pp 1 – 28). La Serena: Ediciones Universidad La Serena.
Mantilla, A. (2000). Germinación y dormición de las semillas. En Azcon-Bieto, J., Talon, M. (eds). Fisiologia vegetal (pp 445 -449). Madrid: McGraw-Hill. Interamericana.
Moreira de C, N., &Nakagawa, J. (1988). Semillas: Ciencia, tecnología y producción. 1ra ed. Montevideo: Hemisferio sur S. R. L. 406 p.
Mogollón, J, G., Díaz, J, G., & Hernández, N. (2004). Multiplicación clonal y enraizamiento in vitro de Ananas comosusL. “Queen Australia”. Rev. Farm. Agron. (LUZ), 1, 15 – 21.
Ortega, D, F., Tamayo, A, C., Calderón, J., & Galván, R. (2012). Establecimiento aséptico en la micropropagación in vitro de Banano Williams (AAA, Subgrupo Cavendish). Tierra Tropical, 7 (2), 205–220.
Pedroza, M, J. (2009). Efecto del carbón activado, Acido indolacético (AIA) y Bencilaminopurina (BAP) en el desarrollo de protocormos de Epidendrum elongatum Jacq. bajo condiciones in vitro. Revista Colombiana de Biotecnología, 10 (1), 17–32.
Quiñones, M.; Miguel, M.; Aleixandre, A. (2012). Los polifenoles, compuestos de origen natural con efectos saludables sobre el sistema cardiovascular. Nutrición Hospitalaria, 27 (1), 76–89.
Rocha, M, S, de E. (2002). Princípios da cultura de tecidos vegetais. 1ra ed. Planaltina: Embrapa Cerrados. 16 p.
Rodríguez, M, N., Fuentes, F, V., Rodríguez, S, O, L., & Álvarez, B, M. (1997). Cultivo in vitro de embriones maduros e inmaduros de aguacatero (Persea americana Mill.). Agricultura Técnica (Chile), 57 (2), 154–158.
Rache, C, L., & Pacheco, M, J. (2010). Propagación in vitro de plantas adultas de Vaccinium meridionale (Ericaceae). Acta bos. bras., 24 (4), 1086–1095.
Rugna, A., Ricco, R., Gurni, A., & Wagner, M. (2007). Efectos de la Radiación Solar sobre la Producción de Polifenoles en Ejemplares Femeninos de Smilax Campestris Griseb. -Smilacaceae-. Latin American Journal of Pharmacy, 26 (3): 420–423.
Sorol, C, B., Haupenthal, B, D., & Reckziegel, M, E. (2012). Caracterización de la germinación, la plántula y el crecimiento de Acrocomia aculeata (Jacq.) Lodd ex. Mart. Rojasiana, 11 (1 – 2), 21–30.
Souto, G. (2008). Agricultural insurance. Paraguay taps its potential for biofuel production. Fourth year/Second phase.Asunción.Information and Communication, IICA. 50 p.
Schmidt, B, F. (2008).Cultivo in vitro e embriogénese somática de embrioes zigóticos de macaúba (Acrocomia aculeata (Jacq.) Loddiges). (Trabajo de posgrado, Ciencia Forestal).Minas Gerais: Universidade Federal de Viçosa. 92 p.
Villegas, F, Z., Giménez, A, C., Vílchez, P, J., Moreno, C, M., Sandoval, L., & Colmenares, E, M. (2008). Oxidación en la inducción de la embriogénesis somática a partir de flores masculinas inmaduras de Gran Enano (Musa AAA). Rev. Fac. Agron., 25: 570–588.
Zárate, R., Aparicio, A., Cantos, M., & Troncoso, A. (1997).
Echinospartum algibicum (Leguminosae) regeneración de plantas mediante organogénesis adventicia. Acta Botánica Malacitana, 22, 35–42.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
 |
Fiori F, C., Díaz L, M. & González S, L. (2016) Enraizamiento in vitro de embriones cigóticos de Acrocomia aculeata (JACQ.) LODD EX MART. Colombia Forestal, 19(1), 67-78. |
Artículo de investigación
ENRAIZAMIENTO IN VITRO DE EMBRIONES CIGÓTICOS DE Acrocomia aculeata (Jacq.) Lodd ex Mart
Rooting in vitro of zygotic embryos of Acrocomia aculeata (Jacq.) Lodd ex Mart
Carlos Fiori Fernández1, Maura Isabel Díaz Lezcano2 & Luis Roberto González Segnana3
1Universidad Nacional de Asunción. San Lorenzo, Paraguay. c_fiori88@hotmail.com. Autor para correspondencia.
2Universidad Nacional de Asunción. San Lorenzo, Paraguay. maura.diaz@agr.una.py
3Universidad Nacional de Asunción. San Lorenzo, Paraguay. biotecno@agr.una.py
RESUMEN
Acrocomia aculeata es una especie potencial para la industria energética; sin embargo, la latencia de sus semillas retrasa la germinación, afectando al rendimiento. El experimento se realizó en el Laboratorio de Biología de la Facultad de Ciencias Agrarias de la Universidad Nacional de Asunción, el objetivo fue establecer un protocolo de embriogénesis cigótica y enraizamiento de esta especie. Se utilizó un diseño completamente al azar, aplicándose el test Kruskal-Wallis y Mann-Whitney con 5% de probabilidad de error, previa comprobación de supuestos de distribución. Los tratamientos fueron medios MS (Murashige y Skoog) con bencilaminopurina (BAP) en 1, 2 y 4 ppm, medios MS con 2 g/L de carbón activado (CA) y MS simples. Se utilizaron 45 embriones cigóticos cultivados inicialmente en medios MS con 2g/L de CA, expuestos a oscuridad inicial por 20 días; posteriormente subcultivados en los tratamientos. La variable medida fue la longitud radicular, evaluada en dos periodos de 20 días. No existió efectos significativos en los tratamientos con BAP, el CA suprimió la oxidación en explantes; mientras que, los mejores resultados presentaron los tratamientos 1 (MS+1 ppm de BAP) y 4 (MS+CA 2g/L) con 6 y 5 cm de longitud radicular respectivamente, luego de 40 días de incubación.
Palabras clave: bencilaminopurina, cultivo oleaginoso, desarrollo in vitro, latencia.
ABSTRACT
Acrocomia aculeata is a potential specie for the energy industry, however, the latency in its seeds delay germination, affecting performance. The experiment was performed in the Biology Laboratory of the Faculty of Agricultural Sciences of the National University of Asunción; the objective was to establish a protocol for zygotic embryogenesis and rooting of this species. A completely randomized design was used, applying Kruskal-Wallis and Mann-Whitney test with 5% error probability, subject to verification of assumptions of distribution The treatments consisted of MS medium (Murashige and Skoog) supplemented with benzylaminopurine (BAP) of 1, 2 and 4 ppm, MS medium with 2 g/L of active carbon (AC) and simple MS mediums. 45 zygotic embryos were initially cultivated on MS medium supplemented with 2 g/L of AC and exposed to initial darkness for 20 days, then subcultured in the mentioned treatments. Root length was the measured variable, assessed in two periods of 20 days. There was no significant effects in the treatments with BAP, the CA suppressed explants oxidation; whereas the best results showed treatments 1 (MS + 1 ppm BAP) and 4 (MS + CA 2g/L) with 6 and 5 cm of root length respectively, after 40 incubation days.
Keywords: benzylaminopurine, oily crop, in vitro rooting, latency.
INTRODUCCIÓN
Acrocomia aculeata (Jacq.) Lodd ex Mart. comúnmente conocida en Paraguay como “mbokaja” es una palmera que se distribuye a lo largo de América tropical y subtropical, nativa de la vegetación de Paraguay y Brasil (Sorol et al., 2012; Chorfi, 2013), sirve de alimento para animales domésticos y como sustento familiar proporciona ingresos por la venta de frutos de los que se extraen aceite que se emplea para la elaboración de jabones; así mismo, los troncos partidos se utilizan para construcción de techos y paredes. También se comercializa la almendra ya sea en forma triturada o no, utilizándose gastronómicamente. Entre todos los valores citados se destaca el aceite tanto de la almendra (55% a 60%) como de la pulpa (4% a 5% de la fruta), importante para la industria aceitera (IICA, 2007; Ganduglia et al., 2009).
Paraguay presenta un importante potencial para convertirse en productor y exportador de este tipo de combustible, debido a sus características como país agropecuario y forestal por su clima y suelo favorable para la producción de este rubro. Sumado a ello se encuentran sus condiciones locales (clima, suelo, mano de obra, tierra disponible) que favorecen al cultivo, del cual se obtiene buen rendimiento y presenta condiciones para ser utilizado como materia prima en la generación de combustibles de origen biológico (Bohn, 2009).
Souto (2008) indica que del fruto pueden extraerse aproximadamente 4000 litros de aceite por hectárea, producción mucho mayor comparado con otros cultivos también aceiteros como el aguacate (Persea americana), ricino (Ricinus communis), colza (Brassica napus), maní (Arrachis hipogeae), girasol (Helianthus annuus), tung (Aleurites fordii) y soja (Glicine max).
Esta especie presenta problemas para la germinación de las semillas por la lentitud, el bajo porcentaje y, en muchos casos, la pérdida de viabilidad por deshidratación. Este problema de germinación se conoce como latencia y es debida a características naturales presentes en la semilla constituyendo así un factor limitante y restrictivo en la producción del rubro (Schmidt, 2008).
Una solución viable y útil mediante una técnica que no requiera enormes inversiones y gastos múltiples es el método de propagación in vitro. Esta técnica consiste en aislar una porción de la planta y proporcionarle las condiciones físicas y químicas necesarias para su desarrollo (CIAT, 1991).
El proceso de enraizamiento consiste en uno de los eventos fisiológicos más importantes realizado durante el cultivo in vitro, pues determinará el establecimiento en el invernadero y posteriormente a campo, donde finalmente se desarrollarán las plantas originadas en el laboratorio. El inicio del desarrollo de la semilla que sirve primeramente de anclaje para la futura planta y posterior medio para la absorción de agua y nutrientes es la emergencia de la radícula cuyo proceso está definido por reguladores de crecimiento (por lo general las Auxinas) que de manera endógena se encuentran en los vegetales (Bidwell, 1993; Azcón-Bieto & Talon, 2000).
El objetivo del presente trabajo fue evaluar el desarrollo radicular de embriones cigóticos de A. aculeata (mbokaja) en cultivo in vitro en diferentes medios de cultivo y tiempos de subcultivo.
MATERIALES Y MÉTODOS
El proceso experimental se llevó a cabo en el Laboratorio de Biología de la Facultad de Ciencias Agrarias de la Universidad Nacional de Asunción, situada en la ciudad de San Lorenzo, Paraguay, entre los meses de junio y agosto del 2012.
Material vegetal
Se utilizaron embriones originarios de frutos maduros de A. aculeata como fuente de explantes para el cultivo in vitro, colectados entre los meses de febrero y marzo del 2012 de una finca localizada en la ciudad de San Lorenzo, Departamento Central, donde fueron seleccionadas cinco plantas madres que exhibieron mayor fructificación. De cada planta madre fueron cosechados 10 frutos, totalizando de esa forma 50 frutos, los cuales fueron mezclados para su posterior secado a temperatura ambiente durante 20 días para la eliminación del pericarpio.
Luego de la eliminación del pericarpio, se procedió a la ruptura del endocarpo con la ayuda de una prensa manual, para que de esa manera se produzca la liberación de las almendras (semilla) que contienen los embriones cigóticos. Aquellas almendras dañadas o afectadas de algún modo por lesiones mecánicas, infecciones fúngicas, síntomas de infecciones bacterianas o ataques de insectos fueron eliminadas por cuestiones de bioseguridad.
La esterilización de los materiales como placas de Petri, mango de bisturí, pinzas de punta fina, agua destilada y frascos fue realizada en autoclave a una temperatura de 121°C a 1 atmósfera de presión durante 50 minutos.
Desinfestación de las almendras
Previamente a la escisión, las almendras se sometieron a unos tratamientos previos compuestos de dos etapas:
1. una inmersión en una solución constituida por alcohol al 90%, durante 4 minutos acompañada de leves agitaciones para favorecer la acción desinfectante del alcohol en las superficies de todas las almendras.
2. consistió en una desinfestación con inmersión en solución de 20% de hipoclorito de sodio durante 20 minutos, acompañado de suaves agitaciones por la razón citada en la primera etapa.
Concluida la segunda etapa se efectuó el triple enjuague de las almendras con agua destilada esterilizada para eliminar los restos de soluciones que pudieran arrastrar las almendras. Todo el proceso de desinfestación se llevó a cabo dentro de la cámara de flujo laminar para aumentar las medidas de bioasepsia.
Escisión de los embriones
Las almendras fueron depositadas en cajas de Petri esterilizadas. Con ayuda de un bisturí y una pinza de punta fina, para la sujeción, se realizaron dos incisiones a ambos lados de la almendra, en la zona media, tomando como referencia el punto de inserción del embrión, el cual es levemente prominente, al momento de realizar la segunda incisión se acompañó con una leve torsión para facilitar la fisión de la almendra y así poder extraer el embrión.
De manera inicial, para la germinación, los embriones extraídos fueron cultivados en el primer medio de cultivo con carbón activado (CA) a razón de 2 g/L y sometidos a oscuridad inicial durante 20 días para de esa manera lograr la supresión de la oxidación de los explantes. Posteriormente, los mismos fueron expuestos a fotoperiodo constante de 16 horas luz en la sala de crecimiento a 25°C.
Tratamientos
Una vez germinados, los embriones fueron subcultivados y sometidos a cinco tratamientos consistentes en T1: medios MS suplementados con 1 ppm de BAP, T2: MS + 2 ppm de BAP, T3: MS+4 ppm de BAP, T4: MS + 2 g/L de CA y T5: medios MS sin concentraciones de BAP ni CA. Las mismas fueron expuestas a fotoperiodo constante de 16 horas luz.
Población y tamaño de muestra
Se utilizaron 50 frutos, de las mismas se extrajeron al azar 45 embriones cigóticos para el cultivo in vitro, cada repetición estuvo compuesta de 3 embriones cultivados en frascos individuales, basados en estudios realizados por Schmidt (2008) y González (2010), los mismo enfrentaron en gran medida la oxidación así como también la presencia de embriones indiferenciados en cuanto a sus estructuras tanto aéreas como radiculares. No se puede dejar de mencionar la contaminación masiva de embriones, y como lo sostiene Rocha (2002), la misma ocasiona una pérdida total de los explantes cultivados in vitro. Los frutos no utilizados fueron tenidos en cuenta como reservas ante cualquier tipo de imprevistos.
Variables
La variable analizada en el trabajo fue la longitud de raíces generadas por los embriones cigóticos sembrados in vitro, a los 20 y 40 días posteriores al subcultivo. La medición se realizó con la ayuda de una regla milimetrada y los valores expresados en centímetros.
Las evaluaciones de medición se realizaron luego de veinte días de la siembra de los embriones cigóticos en el primer medio de cultivo con carbón activado (CA) a razón de 2 g/L para de esa manera lograr la supresión de oxidación para posteriormente ser subcultivados en los diferentes tratamientos.
Diseño experimental
El trabajo consistió en un diseño completamente al azar, con cinco tratamientos y tres repeticiones. Se aplicaron las pruebas de Shapiro-Wilk para corroborar aspectos de normalidad en la distribución de datos, la prueba de Kruskal-Wallis para diferir efectos entre tratamientos y la prueba de Mann-Whitney para observar diferencias entre las comparaciones pareadas. En la totalidad de los procedimientos descritos se utilizó un nivel de significancia de (p<0.005). Las operaciones fueron llevadas a cabo con la ayuda del software estadístico PAST3.
RESULTADOS
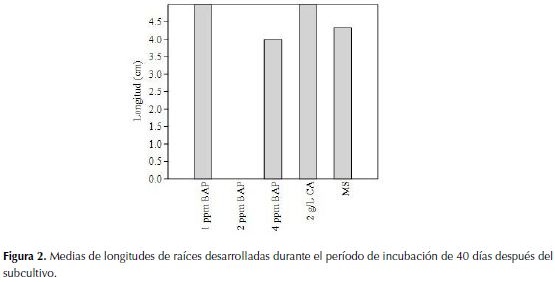
Los embriones sometidos al primer medio de cultivo con CA a razón de 2 g/L durante 20 días, fueron traspasados a frascos que contenía medios enriquecidos con BAP (1, 2 y 4 ppm) CA (2 g/) y MS en donde luego de 20 días de esa operación no presentaron diferencias significativas entre tratamientos. Los resultados observados mediante el test de Shapiro-Wilk (p>0.005) demostraron que los datos no seguían una distribución normal respecto a la media por lo que se procedió a aplicar Kruskal-Wallis para las mediciones efectuadas 20 y 40 días después del subcultivo en los diferentes tratamientos con BAP, MS y CA corroborándose que tampoco existió diferencias significativas (p=0.129>0.005) y (p=0.067>0.005) respectivamente. De forma que al observar los datos arrojados de manera individual en cada tratamiento y repetición, se procedió a aplicar la prueba de Mann-Whitney (Tabla 1) donde no se observó diferencias entre comparaciones pareadas entre tratamientos luego de 20 días de incubación de fotoperiodo constante de 16 horas luz, verificándose de esta manera que ningún tratamiento resultó significativo y por ende ninguna dosis de BAP.
Pudo observarse que las dosis de CA (2g/L) logró suprimir la oxidación en explantes en ambos periodos de incubación posibilitando de esa manera la obtención y desarrollo de la totalidad de los embriones cultivados durante al primer cultivo y luego del subcultivo.
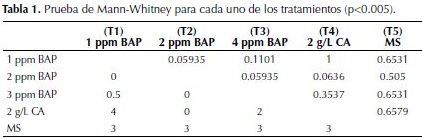
Resulta importante destacar, en términos de medias, que las mejores respuestas de medidas se desarrollaron en los tratamientos 1 (1 ppm de BAP) y 4 (2 g/L CA) con medias de 4.66 cm y 3.33 cm respectivamente, como se observa en la figura 1. En el tratamiento 2 (2ppm de BAP) no se observan datos ya que la misma no generó respuesta alguna.
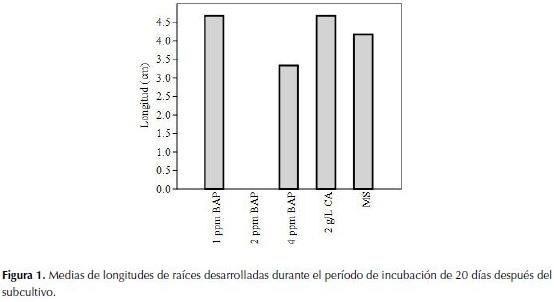
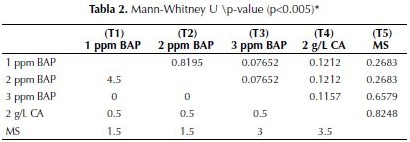
De la misma manera se representa en la figura 2 las mejores medias de los tratamientos producidos durante 40 días de incubación a fotoperiodo constante de 16 horas luz, donde se observa un leve aumento en las longitudes de raíces en los mismos tratamientos, 1 (1 ppm de BAP) con una media de 6 cm y el 4 (2 g/L de CA) con 5 cm de longitud. Como se observa en la tabla 2, estos resultados no arrojaron diferencias significativas entre tratamiento luego de 40 días.
DISCUSIÓN
Entre los principales beneficios de la utilización del CA en los medios de cultivo se cita la supresión de la oxidación pues la misma actúa como soporte catalítico en la oxidación del fenol; así mismo, las condiciones de oscuridad inicial suprimen rutas metabólicas de cierto compuestos en especial el ciclo sikimico (Quiñones et al., 2012), dependiente de la luz del cual derivan los compuestos fenólicos que por medio de oxidación de sus partes químicas se convierten en quinonas muy reactivas capaces de necrosar los tejidos y causar muerte celular (Cáceres, 2004; Rugna et al., 2007; Villegas et al., 2008 mencionados en González, 2010).
Azofeifa (2009) menciona que el efecto benéfico del CA se atribuye a su capacidad para remover sustancias inhibitorias o tóxicas del medio de cultivo que son producidas durante el autoclavado del medio o liberadas por el explante. Dentro de las sustancias producidas durante el autoclavado, se ha reportado la presencia del 5-(hidroximetil)-2-furaldehído (HMF), compuesto inhibitorio, formado primeramente a partir de la fructuosa, ya sea agregada al medio de cultivo o formada por hidrólisis de sacarosa durante el proceso de autoclavado. El empleo del CA agregado al medio de cultivo es frecuente, especialmente en especies de la familia Arecaceae. El cultivo in vitro de diferentes tipos de palmeras presentan problemas de oxidación y la inclusión de CA al medio de cultivo tiene un papel preponderante en la disminución del mismo. Las concentraciones empleadas que se observan en la literatura varían entre 0.5 y 10 g/L, siendo más frecuentes las dosis de 2.0 y 3.0 g/L.
Sumados a los beneficios de mantener en oscuridad a los explantes como medida para atenuar la oxidación por lo anteriormente mencionado, se debe resaltar que la incorporación de concentraciones iguales a 2g/L de CA ofrecerían condiciones de oscuridad, debido a que por naturaleza las moléculas carbonosas son oscuras según Schmitd (2008) Figura 3.

La adición de CA en los medios de cultivo MS ayuda a promover un enraizamiento efectivo de embriones, estos resultados coinciden con los mencionados por Rodríguez et al, (1997) en sus trabajos realizados con embriones maduros de Persea americana Mill, del mismo modo Pedroza (2009) sostiene que la adición de CA en los medios de cultivo incidió de manera positiva en el desarrollo tanto radicular como caulinar y foliar en protocormos de Epidendrum elongatum Jacq. cultivados in vitro. Rache & Pacheco (2010) sostienen que es posible suprimir el uso de reguladores de crecimiento utilizando, para el enraizamiento, concentraciones próximas a los 3 g/L de CA basados en sus experiencias de propagación in vitro de Vaccinium meridionale.
Así mismo, sustancias como carbohidratos, proteínas y lípidos nutren el crecimiento del eje embrionario hasta el punto en que la plántula resultante haya desarrollado un sistema radical capaz de retirar sustancias que la planta necesita (Moreira & Nakagawa, 1988). La mayor parte de los trabajos de movilización y degradación de sustancias de reserva se llevó a cabo en monocotiledóneas. En estas semillas el proceso está controlado por el embrión (Mantilla, 2000). Por lo general todo proceso de germinación se inicia con la salida de la radícula, evento final del proceso de iniciación (Besnier, 1989). El embrión consta de un eje, el hipocotilo, que lleva en un extremo, el meristemo radical y, en el otro, el cotiledón o cotiledones y el meristemo del primer brote (Esau, 1985).
Es importante destacar que el inicio y desarrollo radicular no fue inducido, es decir, no estuvo controlado por la adición exógena de auxina, aunque las mismas presentes de manera endógena y en interacción con las concentraciones de BAP adicionadas más las dosis de CA de manera aparente pudieron haber generado este inicio lo que coincide con los trabajos realizados por Borges et al., (2011) durante su proceso de optimización de medios de cultivo para plantas micropropagadas de Dioscorea alata L.
Así mismo, con los de Zárate et al, (1997) durante sus experiencias en la regeneración de Echinospartum algibicum, estos mismo autores también obtuvieron resultados de enraizamiento en medios con BAP en sus trabajos de inducción de brotes múltiples con Atropa baetica atribuyendo un efecto residual del BAP en el desarrollo de los explantes. Mismos efectos atribuidos al poder residual del BAP en el desarrollo de in vitro de meristemas de Banano Williams fueron mencionados por Ortega et al., (2012).
El poder residual del BAP no afecta la capacidad de enraizamiento de los explantes, sino que se ven favorecidos con el inicio y desarrollo de raíces como se observó en los embriones cigóticos de A. aculeata luego de 40 días de exposición a fotoperiodo constante de 16 horas de luz, misma afirmación fue propuesta por Domínguez et al. (2008), al notar efectos similares en agaves mexicanos expuestos en BAP.
Las citoquininas son un grupo de fitohormonas que juegan un importante en el crecimiento y desarrollo de la planta, resultando su actividad en la morfogénesis y en el metabolismo muy dependiente de factores ambientales (Francescangeli & Zagabria, 2010). En el trabajo realizado por Mogollón et al., (2004), sostiene que una menor incorporación de auxinas en el medio y más elevada concentración de citoquininas, en este caso BAP resulta positivo para el desarrollo de explantes en cuanto a órganos esenciales, entre ellas las raíces.
En general, la acción de las auxinas no son aisladas y otras hormonas como etileno, giberelinas y citoquininas también influyen. Las citoquininas son las hormonas claves para inducir la formación de novo de brotes en diversos explantes in vitro (hojas, raíces, medula, cotiledones). Junto a auxinas, promueven la producción de tejidos no organizados denominados callos, de los cuales es también posible inducir la formación de brotes y/o raíces (Jordan & Casaretto, 2006). A parte de los efectos producidos en el alargamiento de raíces y tallos influyen de forma decisiva en procesos como división celular del cambium, la diferenciación vascular, la dominancia apical y la formación del futo (Azcón-Bieto & Talon, 2000).
Además, el efecto de elongación que ejerce la auxina natural AIA (ácido indolacético) sobre la planta, no solo es evidente desde el punto de vista del crecimiento primario del vástago, sino también desde el punto de vista del crecimiento del sistema radicular porque diversos estudios han demostrado que las auxinas estimulan la rizogénesis en el cultivo de tejidos vegetales bajo condiciones in vitro (Pedroza, 2009).
La auxina trabaja conjuntamente con otro grupo de hormonas vegetales como las citoquininas que permiten el desarrollo de cloroplastos en oscuridad, reemplazando parcialmente la demanda de luz. Una mayor permanencia de clorofilas activas implica para la hoja y planta la conservación de síntesis de proteínas y consiguiente transcripción de varios genes. Esto se ha demostrado con la expresión de varias bandas de proteínas que no son visualizadas cuando el tejido envejece. La presencia de citoquininas provoca un efecto “sumidero” (sink) en el transporte de varias “materias primas” (como aminoácidos) hacia tejidos donde se encuentra la hormona y donde estos recursos serán usados para la síntesis de nuevas proteínas (Jordan & Casaretto, 2006).
En la parte aérea de una planta la auxina emigra desde el ápice en que se producen hacia las zonas de crecimiento que está debajo. Este transporte se produce, a veces, en zonas sin diferenciar los elementos vasculares y por tanto ocurre de una célula a otra a una velocidad aproximada de 1 cm/h. Este transporte tiene la particularidad de que ocurre solo en una dirección, es decir del ápice hacia la base en lo que se denomina un transporte polar (Díaz de la Guardia, 2010).
En los trabajos de Ferreira (2007) y Schmidt (2008) se mencionan las dificultades en la rizogénesis, así como el escaso enraizamiento de la especie recomendando mayor número de investigaciones, para de esa manera posibilitar el éxito en la aclimatación de las vitroplantas por lo que se podría decir que el genotipo marcaría una gran brecha en cuanto al desarrollo de las plantas cultivadas in vitro pues no todas ellas producirán los efectos esperados, dependerá en gran manera de la procedencia del explante y las condiciones ambientales.
El efecto residual del BAP conjuntamente con la adición de CA para suprimir la oxidación de los explantes genera eventos de enraizamiento en embriones cigóticos provenientes de frutos maduros de A. aculeata. Los mismos aparentemente, bajo estas condiciones necesitarían 40 días para que la iniciación y desarrollo de raíces pueda ocurrir, aunque los mismos no hayan producido diferencias desde el punto de vista estadístico, resulta importante destacar y describir los resultados arrojados durante el proceso de desarrollo y culminación del experimento.
CONCLUSIONES
El CA en concentraciones de 2 g/L resultó efectivo en la supresión de la oxidación de los embriones cigóticos de A. aculeata posibilitando de esa manera un efectivo desarrollo de raíces.
En cuanto a los tratamientos con BAP, las mejores medias mostraron los tratamientos 1 (MS + 1 ppm de BAP ) y (MS + 2 g/L de CA) observándose 6 y 5 cm respectivamente requiriendo un período de 40 días de fotoperíodo constante de 16 horas luz. Concluyendo de esta manera que fue necesaria la adición de CA en concentraciones de 2 g/L en los medios de cultivo paliando la oxidación de los explantes que junto con los efectos residuales positivos del BAP posibilitaron el inicio y desarrollo de raíces en embriones cigóticos de A. aculeata, logrando vencer la latencia presente en sus semillas que retrasan la germinación y de esta forma fue posible la obtención de plantines producidos in vitro con alto desarrollo de raíces.
Se necesitarían mayores concentraciones y réplicas para futuros experimentos con embriones cigóticos de A. aculeata yde esta manera generar respuestas significativas a nivel estadístico debido a los resultados biológicos arrojados.
AGRADECIMIENTOS
A la Universidad Nacional de Asunción en cuyo Laboratorio de Biología se ha realizado este experimento. A los profesores Maura Díaz, Luis González, Mirta Vera y Líder Ayala por sus valiosas orientaciones en la elaboración del trabajo. A los compañeros de laboratorio por la colaboración y soporte durante el desarrollo del experimento.
REFERENCIAS BIBLIOGRÁFICAS
Azcón-Bieto, J., & Talón, M. (ed.) (2000). Fundamentos de Fisiología Vegetal. Madrid: McGraw-Hill. Interamericana. 522 p.
Azofeifa, A. (2009). Problemas de oxidación y oscurecimiento de explantes cultivados in vitro. Agronomía Mesoamericana, 20(1), 153-175.
Besnier, R. (1989). Semillas: Biología y tecnología. Madrid: Ediciones Mundi-Prensa. 637 p.
Bidwell, R, G. (1993). Fisiología vegetal. México D. F: A. G. T Editor S. A. 784 p.
Bohn, E. (2009). ‘’Tablero de comando’’ para la promoción de los biocombustibles en Paraguay. Publicación de la Naciones Unidas. CEPAL - Colección de documentos y proyectos. Bundesministerium für wirtschaftliche Zusammenar beit und Entwicklung.GTZ. Santiago de Chile. 112 p.
Borges, G, M., Destrade, B, R., Meneses, R, S., Gómez, K, R., Malaurie, B., Hamon, P., &Demenorval, L, C. (2011).Optimización de un medio de cultivo para plantas micropropagadas de Dioscoreaalata L. Revista Colombiana de Biotecnología, 13(2), 221-228.
Cáceres, A, K. (2004). Propagación in vitro de los porta injertos de cerezo (Prunus avium L.) GISELA 5 Y Prunus cerasus. Universidad Católica de Valparaíso. (Tesis de posgrado, Ciencias en Agronomía). Valparaíso, 69 p.
Centro Internacional de Agricultura Tropical -CIAT-. (1991). Cultivo de tejidos en la agricultura: fundamentos y aplicaciones. Cali: Roca, W, M., Mroginski, L, A. 970 p.
Chorfi, B, L. (2013). Avaliação de populações naturais, estimativas de parãmetros genéticos e seleção de genótipos elite de macaúba (Acrocomia aculeata). (Tesis de posgrado, Agricultura Tropical y Subtropical, Genética, Mejoramiento y Biotecnología Vegetal). Campinas: Instituto Agronómico. 150 p.
Díaz de la Guardia, M. (2010). Fisiología de las plantas. 2da ed. Granada: Grupo Editorial Universitario. 278 p.
Domínguez, R., Alpuche, S, A., Vasco. M, N., & Pérez, M, B. (2008). Efecto de citocininas en la propagación in vitro de Agaves mexicanos. Revista fitotecnia mexicana, 31(4), 317-322.
Esau, K. (1985). Anatomía vegetal. 3ra ed. Barcelona: Ediciones Omega. 779 p.
Ferreira, M, E. (2007). Embriogénese somática de macauba: induçao, regeneraçao e caracterização anatõmica. (Trabajo de posgrado, Genética y Mejoramiento). Minas Gerais: Universidade Federal de Viçosa. 66 p.
Francescangeli, N & Zagabria, A. (2012). Citoquininas para modificar laarquitectura de planta de petunia. Información Técnica Económica Agraria, 106 (1), 46-52.
Ganduglia, F., León, J, G., Gasparini, R., Rodríguez, M, E.,Huarte, G, J., Estrada, J., &Filgueiras, E. (2009). Manual de Biocombustibles. San José: IICA 206 p.
González, D, A. (2010). Efectos del Carbón Activado (CA) y el ácido ascórbico en condiciones de luz y oscuridad sobre la oxidación de embriones zigóticos del mbokaja (Acrocomia aculeata) (Jacq.) en la germinación in vitro. (Trabajo de pregrado, Ingeniería Agronómica). Asunción: Universidad Nacional de Asunción. 91 p.
Instituto Interamericano de Cooperación Asociación Regional de Empresas de Petróleo y para la Agricultura-IICA-. (2007). El estado del arte de los biocombustibles en el Paraguay. Asunción: Editorial Grafir S.A. 83 p.
Jordan, M., &Casaretto, J. (2006).Hormonas y Reguladores del Crecimiento: Auxinas, Giberelinas y Citocininas. En Squeo, F, A., & Cardemil, L. (eds.). Fisiología Vegetal (pp 1 - 28). La Serena: Ediciones Universidad La Serena.
Mantilla, A. (2000). Germinación y dormición de las semillas. En Azcon-Bieto, J., Talon, M. (eds). Fisiologia vegetal (pp 445 -449). Madrid: McGraw-Hill. Interamericana.
Moreira de C, N., &Nakagawa, J. (1988). Semillas: Ciencia, tecnología y producción. 1ra ed. Montevideo: Hemisferio sur S. R. L. 406 p.
Mogollón, J, G., Díaz, J, G., & Hernández, N. (2004). Multiplicación clonal y enraizamiento in vitro de Ananas comosusL. “Queen Australia”. Rev. Farm. Agron. (LUZ), 1, 15 - 21.
Ortega, D, F., Tamayo, A, C., Calderón, J., & Galván, R. (2012). Establecimiento aséptico en la micropropagación in vitro de Banano Williams (AAA, Subgrupo Cavendish). Tierra Tropical, 7 (2), 205-220.
Pedroza, M, J. (2009). Efecto del carbón activado, Acido indolacético (AIA) y Bencilaminopurina (BAP) en el desarrollo de protocormos de Epidendrum elongatum Jacq. bajo condiciones in vitro. Revista Colombiana de Biotecnología, 10 (1), 17-32.
Quiñones, M.; Miguel, M.; Aleixandre, A. (2012). Los polifenoles, compuestos de origen natural con efectos saludables sobre el sistema cardiovascular. Nutrición Hospitalaria, 27 (1), 76-89.
Rocha, M, S, de E. (2002). Princípios da cultura de tecidos vegetais. 1ra ed. Planaltina: Embrapa Cerrados. 16 p.
Rodríguez, M, N., Fuentes, F, V., Rodríguez, S, O, L., & álvarez, B, M. (1997). Cultivo in vitro de embriones maduros e inmaduros de aguacatero (Persea americana Mill.). Agricultura Técnica (Chile), 57 (2), 154-158.
Rache, C, L., & Pacheco, M, J. (2010). Propagación in vitro de plantas adultas de Vaccinium meridionale (Ericaceae). Acta bos. bras., 24 (4), 1086-1095.
Rugna, A., Ricco, R., Gurni, A., & Wagner, M. (2007). Efectos de la Radiación Solar sobre la Producción de Polifenoles en Ejemplares Femeninos de Smilax Campestris Griseb. -Smilacaceae-. Latin American Journal of Pharmacy, 26 (3): 420-423.
Sorol, C, B., Haupenthal, B, D., & Reckziegel, M, E. (2012). Caracterización de la germinación, la plántula y el crecimiento de Acrocomia aculeata (Jacq.) Lodd ex. Mart. Rojasiana, 11 (1-2), 21-30.
Souto, G. (2008). Agricultural insurance. Paraguay taps its potential for biofuel production. Fourth year/Second phase.Asunción.Information and Communication, IICA. 50 p.
Schmidt, B, F. (2008).Cultivo in vitro e embriogénese somática de embrioes zigóticos de macaúba (Acrocomia aculeata (Jacq.) Loddiges). (Trabajo de posgrado, Ciencia Forestal).Minas Gerais: Universidade Federal de Viçosa. 92 p.
Villegas, F, Z., Giménez, A, C., Vílchez, P, J., Moreno, C, M., Sandoval, L., & Colmenares, E, M. (2008). Oxidación en la inducción de la embriogénesis somática a partir de flores masculinas inmaduras de Gran Enano (Musa AAA). Revista de la Facultad de Agronomía, 25: 570-588.
Zárate, R., Aparicio, A., Cantos, M., & Troncoso, A. (1997). Echinospartum algibicum (Leguminosae) regeneración de plantas mediante organogénesis adventicia. Acta Botánica Malacitana, 22, 35-42.




Licencia
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






