
DOI:
https://doi.org/10.14483/2256201X.20878Publicado:
01-01-2024Número:
Vol. 27 Núm. 1 (2024): Enero-junioSección:
Artículos de investigación científica y tecnológicaDiagramas de manejo de densidad para Pinus cooperi var. ornelasi (Martínez) Blanco en Durango, México
Density Management Diagrams for Pinus cooperi var. ornelasi (Martínez) Blanco in Durango, Mexico
Palabras clave:
Aclareos, ajuste simultáneo, densidad máxima, índice de espaciamiento relativo, volumen residual (es).Palabras clave:
Thinnings, simultaneous fitting, maximum density, relative spacing index, residual volume (en).Descargas
Referencias
Acevedo-Benítez, F. D. C., González-Tagle, M. A., Himmelsbach, W., Aguirre-Calderón, O. A., & Jiménez-Pérez, J. (2018). Diagrama de manejo de densidad para el género Pinus en el ejido Pueblo Nuevo, Durango. Revista Mexicana de Ciencias Forestales, 9(50), 54-73. https://doi.org/10.29298/rmcf.v9i50.262 DOI: https://doi.org/10.29298/rmcf.v9i50.262
Álvarez-González, J. G. (1999). Estudio de la densidad: determinación de los valores máximo y mínimo. Universidad de Santiago de Compostela. España.
Assmann, E. (1970). The principles of forest yield study. Pergamon Press.
Barrio, M. (2003). Crecimiento y producción de masas naturales de Quercus robur L. en Galicia [Tesis doctoral, Universidad de Santiago de Compostela].
Barrio-Anta, M., & Álvarez-González, J. G. (2005). Development of a stand density management diagram for even-aged pedunculate oak stands and its use in designing thinning schedules. Forestry, 78(3), 209-216. https://doi.org/10.1093/forestry/cpi033 DOI: https://doi.org/10.1093/forestry/cpi033
Borders, B. E. (1989). Systems of equations in forest stand modeling. Forest Science, 35(2), 548-556. https://doi.org/10.1093/forestscience/35.2.548
Cabrera-Pérez, R. S., Corral-Rivas, S., Quiñonez-Barraza, G., Nájera-Luna, J. A., Cruz-Cobos, F., & Calderón-Leal, V. H. (2019). Density management diagram for mixed-species forests in the El Salto region, Durango, Mexico. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 25(1), 17-29. https://doi.org/10.5154/r.rchscfa.2018.03.026 DOI: https://doi.org/10.5154/r.rchscfa.2018.03.026
Castaño-Santamaría, J., Barrio-Anta, M., & Álvarez-Álvarez, P. (2013). Regional-scale stand density management diagrams for Pyrenean oak (Quercus pyrenaica Willd.) stands in north-west Spain. iForest-Biogeosciences and Forestry, 6(3), 113. https://doi.org/10.3832/ifor0880-006 DOI: https://doi.org/10.3832/ifor0880-006
Castedo-Dorado, F., Álvarez-Álvarez, P., Crecente-Campo, F., & Barrio-Anta, M. (2007). Consideración de la estabilidad de la masa en los diagramas de manejo de la densidad. Cuadernos de la Sociedad Española de Ciencias Forestales, 23, 63-74. https://doi.org/10.31167/csef.v0i23.9606
Castedo-Dorado, F., Crecente-Campo, F., Álvarez-Álvarez, P., & Barrio-Anta, M. (2009). Development of a stand density management diagram for radiata pine stands including assessment of stand stability. Forestry, 82(1), 1-16. https://doi.org/10.1093/forestry/cpm032 DOI: https://doi.org/10.1093/forestry/cpm032
Condés, S., Del Rio, M., & Sterba, H. (2013). Mixing effect on volume growth of Fagus sylvatica and Pinus sylvestris is modulated by stand density. Forest Ecology and Management, 292, 86-95. https://doi.org/10.1016/j.foreco.2012.12.013 DOI: https://doi.org/10.1016/j.foreco.2012.12.013
Corral-Rivas, S., Álvarez-González, J. G., Corral-Rivas, J. J., Wehenkel, C., & López-Sánchez, C. A. (2015). Diagramas para el manejo de la densidad en bosques mixtos e irregulares de Durango, México. Bosque, 36(3), 409-421. http://dx.doi.org/10.4067/S0717-92002015000300008 DOI: https://doi.org/10.4067/S0717-92002015000300008
Daniel, T. W., Helms, J. A., & Baker, S. F. (1982). Principios de Silvicultura. Mcgraw-Hill.
Day, K. (1998). Stocking standards for uneven-aged interior douglas fir. En A. Vyse, C. Hollstedt & D. Huggard (Eds.), Managing the Dry Douglas-Fir Forests of the Southern Interior: Workshop Proceedings (pp. 37-52). Crown Publications. https://citeseerx.ist.psu.edu/document?repid=rep1&type=pdf&doi=dd1cd9aa62f1a01b0a39db2e8e38f4d2818c102a#page=47
Diéguez-Aranda, U., Rojo-Alboreca, A., Castedo-Dorado, F., Álvarez-González, J., Barrio-Anta, M., Crecente-Campo, F., González-González, J. M., Pérez-Cruzado, C., Rodríguez-Soalleiro, R., López-Sánchez, C. A., Balboa-Murias, M. A., Gorgoso-Varela, J. J., & Sánchez-Rodríguez, F. (2009). Herramientas selvícolas para la gestión forestal sostenible en Galicia. Dirección Xeral de Montes, Consellería do Medio Rural, Xunta de Galicia.
Ducey, M. J., & Knapp, R. A. (2010). A stand density index for complex mixed species forests in the northeastern United States. Forest Ecology and Management, 260(9), 1613-1622. https://doi.org/10.1016/j.foreco.2010.08.014 DOI: https://doi.org/10.1016/j.foreco.2010.08.014
Escobedo-Soto, A. S. (2014). Diagrama de manejo de densidad para rodales naturales del ejido El Largo, Chihuahua [Tesis de Maestría, Universidad Autónoma de Nuevo León]. http ://eprints.uanl.mx/4294/1/1080253739.pdf
Gadow, K. V., & Hui, G. (1999). Modelling stand development. Kluwer Academic Publishers. https://doi.org/doi:10.1007/978-94-011-4816-0 DOI: https://doi.org/10.1007/978-94-011-4816-0
Ginrich, S. F. (1967). Measuring and evaluating stocking and stand density in upland hardwood forests in the Central States. Forest Science, 13(1), 38-53. https://doi.org/10.1093/forestscience/13.1.38
González-Elizondo, M. S., González-Elizondo, M., & Márquez-Linares, M. A. (2007). Vegetación y ecorregiones de Durango. CIIDIR Unidad Durango.
González-Elizondo, M. S., González-Elizondo, M., Tena-Flores, J. A., Ruacho-González, L., & López-Enríquez. I. L. (2012). Vegetación de la Sierra Madre Occidental, México: una síntesis. Acta Botánica Mexicana, 100, 351-403. https://doi.org/10.21829/abm100.2012.40 DOI: https://doi.org/10.21829/abm100.2012.40
Graciano-Ávila, G., Alanís-Rodríguez, E., Aguirre-Calderón, O. A., González-Tagle, M. A., Treviño-Garza, E. J., Mora-Olivo, A., & Buendía-Rodríguez, E. (2019). Estimación de volumen, biomasa y contenido de carbono en un bosque de clima templado-frío de Durango, México. Revista Fitotecnia Mexicana, 42(2), 119-127. https://doi.org/10.35196/rfm.2019.2.119-127 DOI: https://doi.org/10.35196/rfm.2019.2.119-127
Instituto Nacional de Geografía y Estadística (INEGI) (2010). Compendio de información geográfica municipal 2010. Pueblo Nuevo, Durango. https://www.inegi.org.mx/contenidos/app/mexicocifras/datos_geograficos/10/10023.pdf
Koenker, R. (2015). quantreg: Quantile Regression. R package version 5.11. https://mran.microsoft.com/snapshot/2015-07-07/web/packages/quantreg/index.html
Koenker, R., & Bassett, G. (1978). Regression quantiles. Econometrica: Journal of the Econometric Society, 33-50. https://doi.org/10.2307/1913643 DOI: https://doi.org/10.2307/1913643
Long, J. N., & Shaw, J. D. (2005). A density management diagram for even-aged ponderosa pine stands. Western Journal of Applied Forestry, 20(4), 205-215. https://doi.org/10.1093/wjaf/20.4.205 DOI: https://doi.org/10.1093/wjaf/20.4.205
López-Sánchez, C., & Rodríguez-Soalleiro, R. (2009). A density management diagram including stand stability and crown fire risk for Pseudotsuga menziesii (Mirb.) Franco in Spain. Mountain Research and Development, 29(2), 169-176. https://doi.org/10.1659/mrd.1070 DOI: https://doi.org/10.1659/mrd.1070
Magaña-Torres, O.S., Torres-Rojo, J. M., Rodríguez-Franco, C., Aguirre-Díaz, H., & Fierros-González, A. M. (2008). Predicción de la producción y rendimiento de Pinus rudis Endl. en Aloapan, Oaxaca. Madera y Bosques, 14(1), 5-19. https://doi.org/10.21829/myb.2008.1411214 DOI: https://doi.org/10.21829/myb.2008.1411214
Martínez-Antúnez, P., Wehenkel, C., Hernández-Díaz, J. C., González-Elizondo, M., Corral-Rivas, J. J., & Pinedo-Álvarez, A. (2013). Effect of climate and physiography on the density of tree and shrub species in Northwest Mexico. Polish Journal Ecology, 61(2), 283-295. https://www.cabdirect.org/cabdirect/abstract/20133392896
McCarter, J. B., & Long, J. N. (1986). A lodgepole pine density management diagram. Western Journal of Applied Forestry, 1(1), 6-11. https://doi.org/10.1093/wjaf/1.1.6 DOI: https://doi.org/10.1093/wjaf/1.1.6
Nájera, L. J. A., & Hernández, H. E. (2009). Acumulación de biomasa aérea en un bosque coetáneo de la región de El Salto, Durango. Ra Ximhai, 5, 225-230. http://uaim.edu.mx/webraximhai/Ej-14articulosPDF/07BIOMASA.pdf DOI: https://doi.org/10.35197/rx.05.02.2009.07.jn
Návar, C. J. J., González, N., & Graciano, J. (2005). Carbon stocks and fluxes in reforestated sites of Durango, Mexico. Madera y Bosques, 11, 15-34, https://doi.org/10.21829/myb.2005.1121254 DOI: https://doi.org/10.21829/myb.2005.1121254
Newton, P. F., & Weetman, G. F. (1993). Stand density management diagram and their development and utility in black spruce management. Forestry Chronicle, 69, 421-430. https://doi.org/10.5558/tfc69421-4 DOI: https://doi.org/10.5558/tfc69421-4
Pérez-Cruzado, C. & Rodríguez-Dacosta, L. (2009). Diagramas de manejo de la densidad para plantaciones de Eucalyptus nitens Maid. en el norte de España con evaluación del carbono fijado en biomasa arbórea. En Sociedad Española de Ciencias Forestales y Junta de Castilla y León (Eds.), 5º Congreso Forestal Español. Montes y Sociedad: saber qué hacer (pp. 2-13). Sociedad Española de Ciencias Forestales. http://doi.org/10.1007/s13398-014-0173-7.2
Quiñonez-Barraza, G., Tamarit-Urias, J. C., Martínez-Salvador, M., García-Cuevas, X., Santos-Posadas, H. M., & Santiago-García, W. (2018). Densidad máxima y diagrama de manejo de la densidad para bosques mezclados de Durango, México. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 24(1), 73-90. https://doi.org/10.5154/r.rchscfa.2017.09.056 DOI: https://doi.org/10.5154/r.rchscfa.2017.09.056
Rogers, R. (1983). Guides for thinning shortleaf pine. En E. P. Jones (Ed.), Proceedings of the Second Biennial Southern Silvicultural Research Conference (pp. 217-225). US Department of Agriculture, Forest Service.
R Core Team (2016). RStudio: Integrated Development Environment for R. RStudio, Inc. http://www.rstudio.com/
Sánchez, O., Vega, E., Peters, E., & Monrroy, V. O. (2003). Conservación de ecosistemas templados de montaña en México. Instituto Nacional de Ecología-Secretaría de Medio Ambiente y Recursos Naturales.
Santiago-García, W., De los Santos-Posadas, H. M., Ángeles-Pérez, G., Valdez-Lazalde, J. R., Del Valle-Paniagua, D. H., & Corral-Rivas, J. J. (2013). Auto-aclareo y guías de densidad para Pinus patula mediante el enfoque de regresión de frontera estocástica. Agrociencia, 47(1), 75-89.
SAS Institute Inc. (2009). SAS/ETS 9.2 User’s Guide. SAS Institute Inc.
Tang, X., Pérez-Cruzado, C., Vor, T., Fehrmann, L., Álvarez-González, J. G., & Kleinn, C. (2016). Development of stand density management diagrams for Chinese fir plantations. Forestry: An International Journal of Forest Research, 89(1), 36-45. https://doi.org/10.1093/forestry/cpv024 DOI: https://doi.org/10.1093/forestry/cpv024
Vargas-Larreta, B., López-Sánchez, C. A., Corral-Rivas, J. J., López-Martínez, J. O., Aguirre-Calderón, C. G., & Álvarez-González, J. G. (2017). Allometric equations for estimating biomass and carbon stocks in the temperate forests of north-western Mexico. Forests, 8(8), 269. https://doi.org/10.3390/f8080269 DOI: https://doi.org/10.3390/f8080269
Wittwer, R. F., Lynch, T. M., & Huebschmann, M. M. (1997). Stand density index for shortleaf pine (Pinus echinata Mill.) natural stands. En E. P. Jones (Ed.). Proceedings of the Ninth Biennial Southern Silvicultural Research Conference (pp. 590-596). US Department of Agriculture, Forest Service.
Yang, S. I., & Brandeis, T. J. (2022). Estimating maximum stand density for mixed-hardwood forests among various physiographic zones in the eastern US. Forest Ecology and Management, 521, 120420. https://doi.org/10.1016/j.foreco.2022.120420 DOI: https://doi.org/10.1016/j.foreco.2022.120420
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Recibido: 20 de mayo de 2023; Aceptado: 17 de noviembre de 2023
Resumen
Se desarrollaron diagramas de manejo de densidad (DMD) con datos de 218 sitios permanentes establecidos en rodales puros de Pinus cooperi en Durango, México. Los DMD se basan en el índice de espaciamiento relativo y en tres ecuaciones no lineales que incluyen variables de rodal. La primera relaciona el diámetro cuadrático con la densidad y la altura dominante; la segunda estima el volumen con la biomasa aérea, el diámetro cuadrático y la altura dominante; y la tercera estima la biomasa a partir del diámetro cuadrático, la altura dominante y la densidad. La densidad máxima se estimó mediante regresión cuantílica ajustando la relación densidad-altura dominante. Las ecuaciones explicaron el 92 % (biomasa) y el 99 % (volumen) de la varianza observada en los datos. Los DMD fueron aplicados en la cuantificación y proyección del volumen y la biomasa en rodales de la especie, como apoyo en la toma de decisiones multiobjetivo.
Palabras clave:
aclareos, ajuste simultáneo, densidad máxima, índice de espaciamiento relativo, volumen residual.Abstract
Density Management Diagrams (DMDs) were developed using data from 218 permanent plots established in pure Pinus cooperi stands in Durango, Mexico. These DMDs are based on the relative spacing index and three non-linear equations that incorporate stand variables. The first equation establishes a relationship between quadratic mean diameter, density, and dominant height; the second equation links volume to aboveground biomass, quadratic mean diameter, and dominant height; and the third equation estimates biomass using quadratic mean diameter, dominant height, and density. The maximum density was estimated via quantile regression by fitting the density-dominant height relationship. The equations explained 92% (biomass) and 99% (volume) of the variance observed in the data. The DMD were applied in the quantification and projection of the volume and biomass of stands of the species, as a support for multi-objective decision-making.
Keywords:
thinnings, simultaneous fitting, maximum density, relative spacing index, residual volume.INTRODUCCIÓN
Las masas forestales con presencia del género Pinus L. constituyen uno de los bosques naturales más abundantes del estado de Durango (González-Elizondo et al., 2012). Este ecosistema es sumamente importante, lo cual se debe en gran medida a las operaciones de cosecha forestal que se llevan a cabo en él, que constituyen una importante actividad económica (Sánchez et al., 2003). Sin embargo, las masas totalmente puras son escasas. Por lo tanto, el control de la densidad es clave en su ordenamiento, que se realiza con base en las condiciones de reproducción, ocupación, crecimiento (altura-diámetro), auto-poda, capacidad de liberación y competencia, según la tolerancia de las especies por ecorregiones (Daniel et al., 1982).
El manejo de la densidad del rodal es un proceso de regulación de la ocupación de un sitio a través de la manipulación ordenada de la ocurrencia y la intensidad de las interacciones de competencia entre árboles para lograr un objetivo de manejo deseado (Newton & Weetman, 1993). La densidad de un rodal influye en las interacciones de las especies y varía espacial y temporalmente al interior de los bosques (Condés et al., 2013). Es una característica importante para evaluar el contenido de volumen de madera y el secuestro de carbono de los árboles (Assmann, 1970). Para una amplia aplicabilidad dentro de una región forestal, una medida de la densidad debe ser capaz de controlar un rango de distribuciones de diámetro y composición de especies (Ducey & Knapp, 2010).
El manejo de la densidad en los rodales y el espacio de crecimiento individual es la base de las intervenciones silvícolas (Santiago-García et al., 2013). En ese sentido, se considera un límite mínimo y uno máximo de árboles. En el primer caso, los recursos son utilizados por cada individuo para desarrollar su máxima capacidad de crecimiento en función de su condición genética (Álvarez-González, 1999), mientras que el segundo se puede utilizar como referencia para evaluar la densidad potencial del rodal de la población objetivo, a fin de aplicar los tratamientos silvícolas apropiados (Yang & Brandeis, 2022).
Los diagramas de manejo de densidad (DMD) son una herramienta de apoyo para el manejo forestal que reúne varios de los elementos necesarios para prescribir tratamientos silvícolas (Corral-Rivas et al., 2015), además de ser una fuente de información interesante para comprender las interacciones entre la densidad y el diámetro, la altura y el volumen promedio dentro de un rodal. Un DMD es una representación gráfica mediante la cual se estiman variables dasométricas importantes para los silvicultores de manera rápida y confiable, al mismo tiempo que se pueden diseñar y evaluar alternativas de corta para alcanzar un determinado objetivo de producción, así como cuantificar el volumen, la biomasa aérea y el carbono residuales y extraídos en cada alternativa silvícola (Magaña-Torres et al., 2008).
En bosques templados de México, se han construido DMD para rodales puros y coetáneos de Pinus patula Schltdl. & Cham. (Santiago-García et al., 2013) para bosques mixtos e irregulares (Corral-Rivas et al., 2015) y para rodales mezclados (Quiñonez-Barraza et al., 2018). De igual manera, en el hemisferio norte se han desarrollado para varias especies de coníferas y latifoliadas (Ginrich, 1967; Rogers, 1983; Wittwer et al., 1997; Day, 1998), y en España para Pinus radiata D. Don (Castedo-Dorado et al., 2009) y para Pinus pinaster Aiton (Barrio-Anta & Álvarez-González, 2005). Estos estudios explican la relación que existe entre el número de individuos por hectárea y su tamaño en diferentes situaciones de competencia. En este contexto, el objetivo de este estudio fue desarrollar DMD para masas puras de Pinus cooperi var. ornelasi (Martínez) Blanco en la región forestal de Pueblo Nuevo, Durango, México.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se llevó a cabo en el municipio de Pueblo Nuevo, Durango, México. Localizado entre los paralelos 22° 56’ y 23° 59’ de latitud norte y los meridianos 104° 45’ y 105° 48’ de longitud oeste (INEGI, 2010). Esta región es la más importante en el país en lo que respecta a de producción forestal (Figura 1).
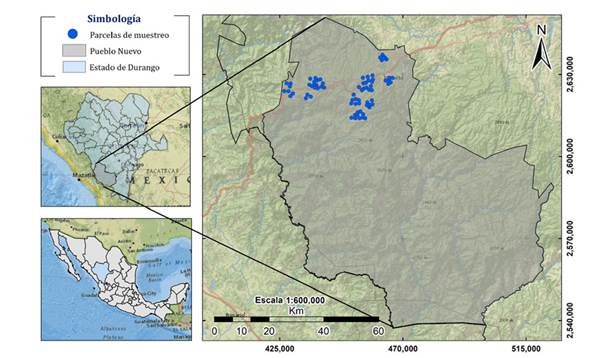
Figura 1: Localización de área de estudio
Su fisiografía se caracteriza por la presencia de terrenos accidentados, con elevaciones y depresiones irregulares. La altitud sobre el nivel del mar varía entre 1400 y 2600 m. El tipo de vegetación existente corresponde a bosques mixtos e irregulares de pino-encino. El clima predominante es templado, con lluvias en verano. La precipitación media anual oscila de 700 a 1440 mm, y el área presenta una temperatura promedio de 8.3 ºC en zonas altas y 16 °C en las partes bajas (González-Elizondo et al., 2007; Martínez-Antúnez et al., 2013).
Datos de campo
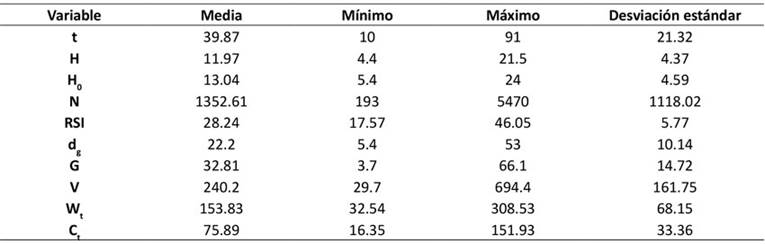
Los datos de campo fueron obtenidos de 218 sitios permanentes de muestreo (25 x 25 m) establecidos en rodales regulares puros para medir el efecto de los aclareos en el crecimiento de P. cooperi. Los sitios cubren el rango de edades, densidades y calidades de los rodales de esta especie. Se registró información de edad (t), altura dominante (H 0) y altura media (H) en m; del número de árboles por hectárea (N); del diámetro medio cuadrático (d g) en cm; del área basal por hectárea (G) en m2; del índice de espaciamiento relativo (RSI) en %; del volumen total del rodal (V) en m3.ha-1; y de biomasa aérea total (W t ) y carbono (C t ) en Mg.ha-1. En la Tabla 1 se presenta el resumen de las estadísticas descriptivos de las variables utilizadas para la construcción de los DMD.
Construcción de los diagramas de manejo de densidad
Tabla 1: Resumen de las estadísticas del conjunto de datos utilizados para la construcción de los diagramas de manejo de densidad
El DMD consta de un sistema de ecuaciones no lineales ajustadas a partir de datos de volumen, biomasa y carbono, así como del índice de espaciamiento relativo (RSI). La primera ecuación del sistema describe la relación alométrica del diámetro medio cuadrático con la densidad (árboles.ha-1) y la altura dominante del rodal (Ecuación (1)) (Long & Shaw, 2005). La segunda ecuación representa la relación entre la producción de rodal, expresada en volumen por hectárea, y el volumen de un árbol representativo, cuyo diámetro es el diámetro medio cuadrático y su altura es la altura dominante del rodal, multiplicado por el número de árboles por hectárea (Ecuación (2)). La tercera ecuación predice la biomasa aérea de manera similar a la del volumen total (Ecuación (3)). El índice de espaciamiento relativo (RSI) es útil en el manejo de la densidad de los rodales, pues es independiente de la calidad del sitio y de la edad del rodal, excepto para rodales muy jóvenes (Gadow & Hui, 1999). McCarter y Long (1986) lo definen como la proporción (%) entre la distancia media de los árboles y la altura dominante, suponiendo una distribución triangular (Ecuación (4)) (Tabla 2). Debido a que los DMD estiman la biomasa aérea total, y a partir de esta se puede obtener el carbono aéreo, se realizaron estimaciones de las reservas de carbono, para lo cual se utilizó la Ecuación (3) y el contenido de carbono reportado para P. cooperi (0.489) por Vargas-Larreta et al. (2017).
N = número de árboles.ha-1, H
0
= altura dominante (m), d
g
= diámetro medio cuadrático (cm), βi (i = 0-10) = coeficientes a estimar en el ajuste, V = volumen total (m3.ha-1), W
t
= biomasa aérea total (Mg.ha-1), RSI = índice de espaciamiento relativo (%)
Tabla 2. : Ecuaciones utilizadas en la construcción de los diagramas de manejo de la densidad

Las Ecuaciones (1), (2) y (3) definen un sistema de ecuaciones estructuralmente simultáneas, donde N y H 0 son variables exógenas (variables que aparecen solamente en la parte derecha de las ecuaciones, cuyos valores se determinan independientemente del sistema), V y W t son variables endógenas (las variables que el modelo intenta predecir y solo aparecen del lado izquierdo de las ecuaciones) y d g es una variable endógena que aparece tanto como variable dependiente como explicativa en el sistema (Borders, 1989). Las ecuaciones no lineales se ajustaron de manera simultánea para evitar la correlación de los errores empleando el método de máxima verosimilitud de información completa (full information maximum likelihood, FIML). El ajuste se realizó mediante el procedimiento MODEL del paquete estadístico SAS/ETS (SAS Institute, 2009).
La línea de densidad máxima es importante en los DMD, pues incluirla permite simular condiciones de ocupación que podrían provocar mortalidad por competencia, al mismo tiempo que controla la prescripción de cortas e implica una relación de copa viva que promueve el buen vigor de los árboles (Barrio, 2003). La línea del límite superior se estimó mediante regresión cuantílica no lineal (Koenker & Bassett, 1978), ajustando una ecuación potencial que relaciona directamente N con los valores de H 0 (Ecuación (5)).
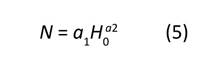
donde: N = número de árboles ha-1, a1 y a2 = parámetros que determinan la línea de autoaclareo. Se utilizó el cuantil 95 %, de tal manera que el límite superior de densidad solo fue superado por el 5 % de los rodales de máxima densidad, correspondientes a rodales con alturas dominantes menores de 8 m. Los parámetros de la Ecuación (5) se obtuvieron mediante la función nlrq del paquete quantreg (Koenker, 2015) del programa R Studio (R Core Team, 2016).
Evaluación del ajuste de los modelos
El ajuste de los modelos se midió a través del gráfico de los residuales y los siguientes criterios estadísticos: el coeficiente de determinación (R 2 ) (Ecuación (6)) y la raíz del cuadrado medio del error (REMC) (Ecuación (7)). Además, se evaluó la significancia de los parámetros a un nivel de probabilidad de 0.05.
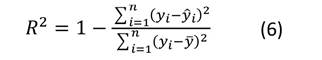
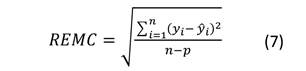
donde: yi, y i, y ̅= valores observado, predicho y medio de la variable dependiente respectivamente; n = número de datos empleados en el ajuste; p = número de parámetros de la ecuación ajustada.
Representación gráfica de las isolíneas
Después de obtener las estimaciones de los parámetros para el sistema de ecuaciones, se representaron las isolíneas de las variables involucradas en el diagrama. Primeramente, se expresó el nivel de espesura de la masa mediante el RSI. Las líneas para este índice se obtuvieron despejando N de la Ecuación (4) y asignando valores a RSI (Ecuación (8)). La representación de las isolíneas para el diámetro medio cuadrático se realizó mediante la Ecuación (1), estableciendo d g como un valor constante (Ecuación (9)) y resolviendo N a través de la gama de alturas (H 0 ) observadas en las parcelas de muestreo. Las isolíneas para el volumen total y la biomasa aérea total se derivaron del despeje de N en las Ecuaciones (2) y (3) respectivamente. Posteriormente, se resolvió N a través del rango de la altura dominante observada (H 0 ), fijando como constante V y W t (Ecuaciones (10) y (11)). No se representaron las isolíneas para los almacenes de carbono total, ya que los valores de esta variable se obtienen del gráfico que presenta la biomasa aérea total, multiplicando el valor de W t por el contenido de C de P. cooperi (0.489).
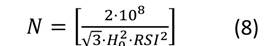
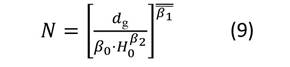
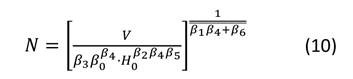
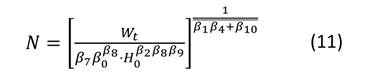
donde: N = número de árboles.ha-1, H 0 = altura dominante (m), d g = diámetro medio cuadrático (cm), βi (i = 0-10) = coeficientes a estimar en el ajuste, V = volumen total (m3.ha-1), W t = biomasa aérea total (Mg.ha-1), RSI = índice de espaciamiento relativo (%).
RESULTADOS
Ajuste de los modelos
La dispersión de los residuales contra los valores predichos no presentó tendencias anómalas que hicieran suponer una violación de los supuestos de regresión como normalidad, homogeneidad de varianzas e independencia de los errores. Por lo tanto, los parámetros se consideraron adecuados para la elaboración del DMD (Figura 2).
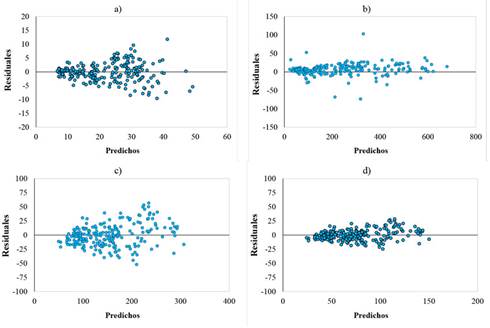
Figura 2.: Valores predichos vs. residuales para dg (a), V (b), Wt (c) y Ct (d), respectivamente
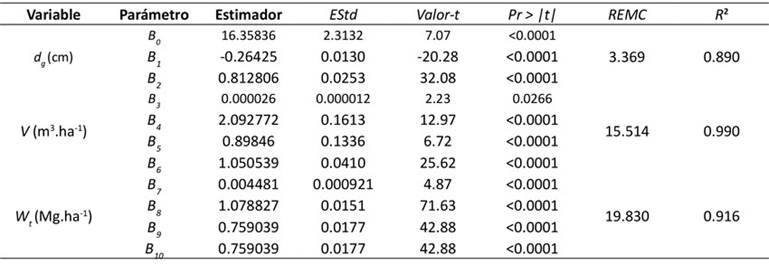
Los estimadores de los parámetros de las ecuaciones resultaron ser significativos a un nivel de 5 % (p<0.05). En la Tabla 3 se presentan los estimadores, su error estándar y su significancia, así como los estadísticos de bondad de ajuste del sistema de ecuaciones (d g, V y W t). El diámetro medio cuadrático se modeló con menor precisión. No obstante, el ajuste se consideró aceptable, ya que la ecuación explicó más del 88 % de la variabilidad de los datos de diámetro cuadrático de las parcelas. Las ecuaciones que obtuvieron los valores más altos de R 2 fueron las de volumen total y biomasa aérea total, explicando alrededor del 99 y 92 % de la variabilidad observada respectivamente. El valor de la raíz del error medio cuadrático (REMC) de la ecuación de d g fue 3.36 cm, mientras que, para las ecuaciones de V y W t , este estadístico presentó valores de 15.51 y 19.83 Mg.ha-1 respectivamente.
EStd = error estándar, Pr > |t| = probabilidad calculada, REMC = raíz del cuadrado medio del error, R
2
= coeficiente de determinación
Tabla 3: Parámetros estimados y estadísticos de bondad del ajuste simultáneo de las Ecuaciones (1), (2) y (3)
En la Tabla 4 se muestran los estimadores de los parámetros de la ecuación que define el límite de densidad máxima, obtenidos mediante el ajuste de la regresión cuantílica no lineal. La pendiente de la línea de autoaclareo fue -1.5764, un valor muy cercano al teórico (-3/2), ampliamente reportado en estudios sobre densidad. Los estimadores resultaron ser significativos al 99 %.
EStd = error estándar, Pr > |t| = probabilidad calculada
Tabla 4: Parámetros estimados para la regresión cuantílica no lineal de la Ecuación (5)

Diagrama de manejo de la densidad
Se elaboraron DMD para volumen total y biomasa aérea total. Para ello, en un primer paso, se trazó la línea de autoaclareo o límite de densidad máxima (Figura 3).
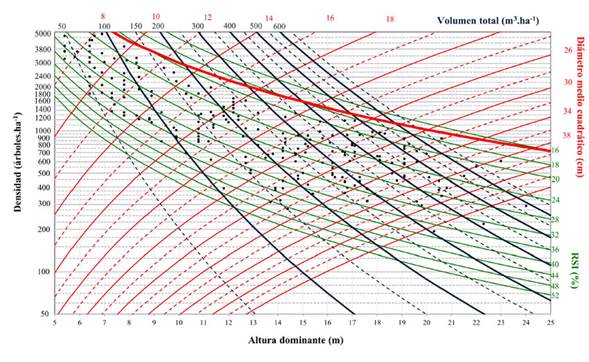
Figura 3: Diagrama de manejo de densidad para Pinus cooperi con límite de densidad máxima (línea roja gruesa)
Posteriormente, se añadieron las isolíneas del RSI y del diámetro medio cuadrático (d g) en un gráfico bivariado, con la altura dominante (H 0 ) en el eje de las abscisas y el número de árboles por hectárea (N) en el eje de las ordenadas (Figuras 4 y 5). Los valores de H 0 oscilaron entre 5 y 25 m, y la densidad (N) fluctuó entre 50 y 5000 árboles.ha-1, utilizando una escala logarítmica.
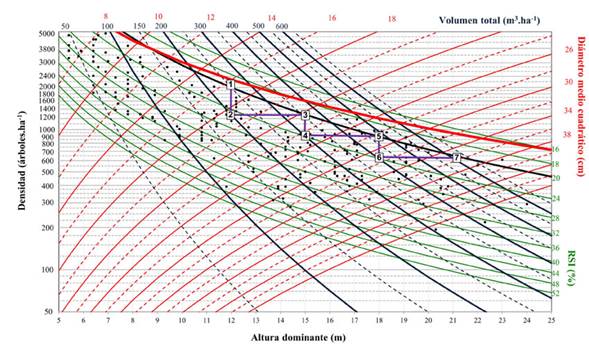
Figura 4: Diagrama de manejo de la densidad para Pinus cooperi con esquema de aclareos que maximiza el volumen total del rodal
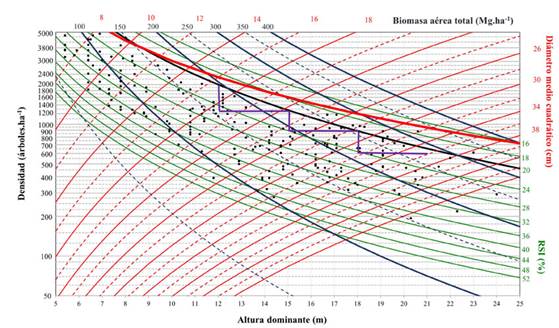
Figura 5: Diagrama de manejo de densidad para Pinus cooperi con esquema de aclareos que maximiza la biomasa aérea del rodal
El diagrama proporciona isolíneas para el RSI. La línea superior o límite de densidad máxima correspondió a un valor de 16 %, aproximándose a la relación tamaño-densidad máxima, que es donde inicia la mortalidad natural y, por otro lado, en el límite inferior de la zona óptima de densidad, las isolíneas restantes presentaron valores cercanos a 52 %. Se fijó un rango de valores de 8 a 44 cm para el d g. Los valores para el volumen total variaron de 50 a 650 m3. ha-1, y las isolíneas presentaron una tendencia ascendente. Los valores de biomasa aérea total oscilaron entre 50 y 400 Mg.ha-1. La tendencia de las isolíneas relacionadas con la biomasa aérea total resultó ser menos vertical que la de aquellas relacionadas con el volumen. Esto se debe a que las isolíneas relacionadas con W t son menos sensibles a los cambios de densidad que las que presenta V.
Sobre el mismo gráfico, se definió el esquema de aclareos de acuerdo al objetivo fijado en el estudio de caso (Figuras 4 y 5, línea negra escalonada). Existen dos factores que determinan el esquema de aclareos: el estado del rodal a la edad de corta y los límites superior e inferior de densidad. El primero está representado por la combinación lógica de las variables que se muestran en el DMD, mientras que los límites de densidad son fijados por el RSI y están representados por el máximo crecimiento por unidad de superficie y el crecimiento y vigor individual del árbol (Diéguez-Aranda et al., 2009).
Cada punto marcado en el diagrama permite estimar los valores de densidad, altura dominante, diámetro medio cuadrático, índice de espaciamiento relativo, volumen total, biomasa aérea total y contenido de carbono aéreo total. Por ejemplo, en el punto final de cosecha en el gráfico (punto 7), los valores fueron 600.31 m3.ha-1, 281 Mg.ha-1 y 137.5 Mg.ha-1 para V y W t y C t respectivamente (Figuras 4 y 5).
DISCUSIÓN
Los valores del coeficiente de determinación de las ecuaciones ajustadas para la construcción de los DMD fueron aceptables (Tabla 3) y en algunos casos superiores a los reportados en otros estudios realizados en bosques regulares. Por ejemplo, Castedo-Dorado et al. (2007) obtuvieron valores R 2 de 0.91 y 0.99 para d g y W t respectivamente, mientras que la R 2 para V fue 0.99 (similar al valor obtenido en esta investigación) en rodales de P. radiata en el noroeste de España. Por su parte, Pérez-Cruzado et al. (2009) obtuvieron valores de R 2 de 0.94, 0.996 y 0.998 para las ecuaciones de dg, V y W t respectivamente en plantaciones de Eucalyptus nitens Maid. en el norte de España. Asimismo, en bosques de coníferas de Durango, México, Corral-Rivas et al. (2015) reportaron coeficientes de determinación de 0.52 a 0.62 para el d g , y 0.83 a 0.84 para V en niveles de productividad alto y medio respectivamente. En bosques de Pinus ubicados en la misma área de estudio de esta investigación, se han registrado valores cercanos a 0.70 para d g y 0.99 para V (Acevedo-Benítez et al., 2018).
Los valores del error estándar (REMC) de las ecuaciones fueron 3.37 cm, 15.51 m3.ha-1 y 19.83 Mg.ha-1 para d g , V y W t respectivamente. Estos valores son similares a los reportados por Corral-Rivas et al. (2015) para el tipo de bosque ‘coníferas’ en Durango, México, donde el REMC para d g fluctuó entre 2.46 y 3.08 cm y osciló entre 16.25 y 29.45 m3.ha-1 para V en niveles de productividad alto y medio. López-Sánchez & Rodríguez- Soalleiro (2009) reportaron precisiones (REMC) de 3.03 cm para dg y 14.5 m3.ha-1 para V en Pseudotsuga menziesii (Mirb.) Franco en España. Por su parte, Castedo-Dorado et al. (2009) obtuvieron valores de error medio por debajo de los obtenidos en este estudio (2.75 cm y 12.03 m3.ha-1 para las ecuaciones de d g y V respectivamente).
Como se mencionó anteriormente, los DMD permiten planificar los esquemas de corta bajo un régimen silvícola particular, el cual generalmente busca maximizar el volumen o la biomasa del rodal. En este estudio de caso, para V, W t y C t , se asumió una masa inicial de 2000 árboles por hectárea y una altura dominante de 12 m, asumiendo además una calidad de estación promedio y un intervalo de corta basado en un incremento en altura dominante de 3 m (aproximadamente cada diez años). Para el límite superior de densidad, se suele fijar un RSI constante, siempre y cuando se encuentre por debajo de la línea de autoaclareo, el cual se fijó en 20 % en este caso (Figuras 4 y 5). La definición del límite superior de los DMD evita que los bosques naturales lleguen a un punto de autoaclareo, i.e., evita que existan pérdidas por mortalidad natural, mientras que el límite inferior representa la relación existente entre los objetivos del manejo y las especies presentes en el rodal (Corral-Rivas et al., 2015). Esto permite interpretar el comportamiento de una masa forestal y, al mismo tiempo, facilitar la toma de decisiones en el desarrollo silvícola.
Para rodales naturales del género Pinus, el límite de densidad máxima puede alcanzarse cuando el índice de espaciamiento relativo se encuentra entre el 25 y el 30 % (Escobedo-Soto, 2014; Acevedo-Benítez et al., 2018; Cabrera-Pérez et al., 2019). Los valores para los limites superior e inferior obtenidos en este estudio concuerdan con lo reportado por Barrio-Anta y Álvarez-González (2005), quienes definieron un valor de RSI = 20 % para el límite superior de las existencias volumétricas en rodales de Quercus robur L. en Galicia, España. Por otro lado, Castedo-Dorado et al. (2009) reportaron un RSI cercano al 16 %, un valor similar al límite de densidad máxima obtenido en este estudio. Por su parte, Castaño-Santamaría et al. (2013) y Tang et al. (2016) obtuvieron valores de RSI cercanos a 24 % para el límite superior de densidad, mientras que, para el límite inferior, obtuvieron 32 y 36 % en rodales de abeto chino y roble melojo respectivamente. Estos valores son mayores que los obtenidos en este estudio.
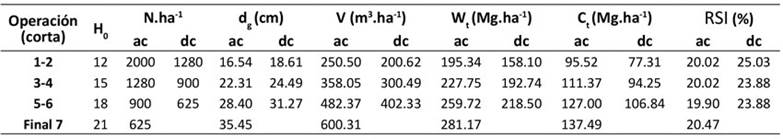
El esquema de manejo utilizado como ejemplo de aplicación del DMD busca maximizar el volumen y la biomasa aérea total, y consta de tres intervenciones silvícolas o aclareos (recorrido de los aclareos 1-2, 3-4 y 5-6, Figura 4) y la edad de cosecha, la cual fue definida para H 0 = 21 m y d g = 35.5 cm (Tabla 5). Utilizando estos valores, la secuencia de intervenciones necesaria para alcanzar el objetivo final (punto 7) se encuentra por escalonamiento inverso (Figura 4, línea negra escalonada).
Los segmentos verticales y horizontales representan, respectivamente, las etapas de aclareos y los intervalos entre estos. Por lo tanto, la reducción de la densidad por dichos aclareos no causa un efecto directo en la altura dominante (López-Sánchez & Rodríguez-Soalleiro, 2009). De acuerdo con el DMD, la primera intervención (punto 2) se deberá realizar cuando el diámetro medio cuadrático alcance un valor de 16.5 cm y la densidad sea cercana a los 2000 árboles.ha-1. En este punto, el volumen correspondería aproximadamente a 250 m3.ha-1. De igual manera, se muestran las estimaciones de las variables para las dos intervenciones restantes (puntos 4 y 6). Las condiciones anteriores definen el punto final de cosecha en el gráfico (punto 7). En este punto resulta una densidad de 625 árboles.ha-1, un RSI de 20.5 % (Figura 4, línea negra gruesa) y existencias maderables cercanas a los 600 m3.ha-1.
La producción total del rodal se puede obtener directamente de cualquier punto del diagrama a partir de las isolíneas que modelan el volumen total. Los volúmenes extraídos durante el primer, segundo y tercer aclareo (la diferencia entre el volumen antes y después de la corta) fueron 50, 57.6 y 80 m3.ha-1 aproximadamente, y el volumen al final de la rotación alcanzó los 600.31 m3.ha-1. La suma de estos volúmenes representa una estimación del volumen del rodal bajo este régimen específico de manejo de la densidad.
Con el esquema de cortas propuesto, y de acuerdo con modelo desarrollado, la biomasa aérea total extraída durante el primer, segundo y tercer aclareos ascendería a 37.2, 35 y 41.2 Mg.ha-1 respectivamente, alcanzando un valor aproximado de 281 Mg ha-1 al final de la rotación, mientras que la cantidad de carbono aéreo total extraído (calculado con una proporción de carbono de 0.489) alcanzaría los 18.2, 17.1 y 20.2 Mg.ha-1. Al final de la rotación, se obtendría un valor de contenido de carbono de 137.5 Mg.ha-1 (Tabla 5). Los valores finales de V y W t y C t son superiores a los reportados en diferentes estudios realizados en la misma región, cuyos objetivos fueron estimar el volumen y el contendido de biomasa y carbono en bosques templados (Návar et al., 2005; Nájera & Hernández, 2009; Graciano-Ávila et al., 2019).
ac = antes de la corta; dc = después de la corta
Tabla 5: Esquema de cortas para volumen, biomasa y carbono total basado en el DMD
CONCLUSIONES
Se desarrollaron diagramas de manejo de la densidad que proyectan la evolución del volumen y la biomasa de masas puras de Pinus cooperi var. ornelasi a partir de variables de fácil medición, tales como el diámetro medio cuadrático y el número de árboles del rodal. El índice de espaciamiento relativo y la altura dominante son las variables más importantes para definir la frecuencia y el momento de las intervenciones silvícolas. Las ecuaciones que integran el DMD estiman de manera precisa e insesgada la ocupación del sitio, el volumen y la biomasa, mientras que la regresión cuantílica ajustada a la relación densidad-altura dominante es una alternativa para calcular de manera confiable la densidad máxima del rodal. Los DMD desarrollados son herramientas disponibles para los manejadores forestales, las cuales les permitirán planificar y evaluar el efecto de una amplia gama de escenarios silvícolas, tanto en la producción maderable como en la estructura de la masa forestal residual. Asimismo, a través de los DMD, es posible evaluar la dinámica del carbono forestal. Esto, mediante de la incorporación de un modelo de biomasa en el sistema de ecuaciones que define las isolíneas del DMD.
Acknowledgements
AGRADECIMIENTOS
A los ejidos forestales de la región forestal de Pueblo Nuevo, Durango, por permitirnos establecer los sitios de muestreo utilizados para la colecta de datos.
REFERENCIA




Licencia
Derechos de autor 2024 Colombia forestal

Esta obra está bajo una licencia internacional Creative Commons Atribución-CompartirIgual 4.0.
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






