
DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2009.1.a13Publicado:
01-01-2009Número:
Vol. 12 Núm. 1 (2009): Enero-DiciembreSección:
Artículos de investigación científica y tecnológicaANÁLISIS ECOANATÓMICO, EVOLUTIVO Y COMPARATIVO DE LA MADERA DE 40 ESPECIES DE DOS ASOCIACIONES DEL BOSQUE ALTOANDINO COLOMBIANO
Ecoanatomical, developing and comparative analysis of 40 wood species belonging to two associations in colombian Andes highland forest
Palabras clave:
índice evolutivo, ecoanatomía, anatomía comparativa de la madera, asociación vegetal, xilología (es).Palabras clave:
evolutive index, ecoanatomy, comparative wood anatomy, vegetal association, xilology. (en).Descargas
Referencias
Aguilar-Rodríguez, S., T. Terrazas, & L. López-Mata. 2006. Anatomical wood variation of Buddeleja cordata (Buddlejaceae) along its natural range in Mexico. Trees 20: 253-261.
Araque, A. & W. León. 2006. Anatomía comparada del leño de Spondias mombin L. (Anacardiaceae) que crece en zonas del banco y bajío de la Reserva Forestal Caparo (Barinas, Venezuela). Revista Forestal Venezolana 50 (1): 9-17.
Arias S. & T. Terrazas. 2001. Variación en la anatomía de la madera de Pachycereus pectenaboriginum (Cactaceae). Serie Botánica 72 (2): 157-169.
Baas, P. & S. Carquilst. 1985. A comparison of the ecological wood and anatomy of the floras of Southern California and Israel. iawa Bulletin 6 (4): 349-353.
Baas, P. & R. Miller. 1985. Functional and ecological wood anatomy. Some introductory comments. iawa Bulletin 6 (4): 281-282.
Baas, P. & F. Schweingruber. 1987. Ecological trends in wood anatomy of trees, shrubs, and climbers from Europe. iawa Bulletin 8 (3): 245274.
Baas, P., E. Werker & A. Fahn. 1983. Some ecological trends in vessel characters. iawa Bulletin 4 (2-3): 141-159.
Barajas-Morales J. 1985. Wood structural differences between trees of two tropical forests in Mexico. iawa Bulletin 6 (1-4): 355-364.
Cantillo, E., A. Avella & K. Rodríguez. 2004. Diversidad y caracterización florística estructural de la vegetación arbórea de la Rserva Forestal Cárpatos, Guasca, Cundinamarca. Revista Colombia Forestal 8 (17): 5-21.
Carlquist, S. 2001. Comparative wood anatomy. Sistematic, ecological and evolutionary aspects of dicotyledon wood. Springer Verlag. Berlín y Heidelberg.
Carlquist, S. 1992. Wood anatomy of Hedyosmum (Chloranthaceae) and the tracheid-vessel element transition. aliso Rancho Santa Ana Botanic Garden 13 (3): 447-462.
Cronquist, A. 1981. An integrated system of classification of flowering plants. Columbia University Press. Nueva York.
Cuéllar-Moyano, F. 2008. Aves falconiformes de Colombia: Águila y afines. Colombia Curiosa. Bogotá.
Grande, C. & C. Polanco. 2007. Descripción anatómica de la madera de 40 especies del bosque altoandino en Guasca, Cundinamarca. Revista Colombia Forestal 10 (20): 180-206.
Gruber, A., D. Baumgartner, J. Zimmermann & W. Oberhuber. 2008. Temporal dynamic of wood formation in Pinus cembra along the alpine treeline ecotone and the effect of climate variables. Trees 23: 623-635.
Holdridge, L. 1979. Ecología basada en zonas de vida. Editorial iica. San José, Costa Rica.
Judd, W., C. Campbell, E. Kellogg & P. Stevens. 1999. Plant systematics. Sinauer Associates. Sunderland, Massachusetts.
Lens, F., J. Luteyn, E. Smets & S. Steven Jansen. 2009. Ecological trends in the wood anatomy of Vaccinioideae (Ericaceae). Flora 199: 309-319.
León, W. 2001. Anatomía del leño, aspectos ecológicos y filogenia en mangles de Venezuela. Revista Forestal Venezolana 45 (2): 191-203.
León, W. 2002. Anatomía xilemática del tronco de Pereskia guamacho (Cactaceae) procedente del Estado de Mérida, Venezuela. Pittieria 31: 4149.
León, W. & N. Espinoza de Pernía. 1999. Parénquima disyuntivo y filogenia. Revista Forestal Venezolana 43 (2): 137-146.
León, W. 2002. Anatomía xilemática del tronco de Pereskia guamacho (Cactaceae) procedente del Estado de Mérida, Venezuela. Pittieria 31: 4149.
León, W. & N. Espinoza de Pernía. 1999. Parénquima disyuntivo y filogenia. Revista Forestal Venezolana 43 (2): 137-146.
León, W. & N. Espinoza de Pernía. 2001. Anatomía de madera. Universidad de los Andes. Mérida, Venezuela.
Loepfe, L., J. Martínez-Vilalta, J. Piñol & M. Mencuccini. 2007. The relevance of xylem network structure for plant hydraulic efficiency and safety. Journal of Theoretical Biology 247: 788-803.
Marquínez, X. 2002. Historia evolutiva de las plantas. Universidad Nacional de Colombia. Bogotá.
Metcalfe, C. 1979. Anatomy of the dicotyledons. Vol. I: Systematic anatomy of the leaf and stem, with a brief history of the subject. Oxford University Press.
Milton, J. & J. Arnold. 2004. Probabilidad y estadística con aplicaciones para ingeniería y ciencias computacionales. McGraw-Hill. México.
Moglia, G. & A. Giménez. 1998. Rasgos anatómicos característicos del hidrosistema de las principales especies arbóreas de la región chaqueña argentina. Investigación Agraria 7 (1 y 2): 53-71.
Olano, C. 2007. Estudios filogenéticos moleculares del género Caryodaphnopsis (Lauraceae) y sinopsis de las especies colombianas. Trabajo de grado para optar al título de Biólogo. Universidad de los Andes. Bogotá.
Oskolski, A. & S. Jansen. 2009. Distribution of scalariform and simple perforation plates within the vessel network in secondary xylem of Araliaceae and its implications for wood evolution. Plant Systematics and Evolution 278: 43-51.
Pérez, A. 1989. Caracterización ecoanatómica del leño de 40 especies del bosque La Mucuy, Estado de Mérida, Venezuela. Revista Forestal Venezolana 33: 43-51.
Rana, R., R. Langenfeld-Heyser, R. Finkeldey & A. Polle. 2008. Functional anatomy of five endangered tropical timber wood species of the family Dipterocarpaceae. Tree 23: 521-529.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
 | Polanco-Tapia, C., & Grande-Pulido, C. (2009). Análisis ecoanatómico, evolutivo y comparativo de la madera de 40 especies de dos asociaciones del bosque altoandino colombiano. Colombia Forestal, 12, 183-203. |
ANÁLISIS ECOANATÓMICO, EVOLUTIVO Y COMPARATIVO DE LA MADERA DE 40 ESPECIES DE DOS ASOCIACIONES DEL BOSQUE ALTOANDINO COLOMBIANO
Ecoanatomical, developing and comparative analysis of 40 wood species belonging to two associations in colombian Andes highland forest
Análises eco-anatômicas, evolutiva e comparativa da madeira de 40 espécies das associações do bosque alto-andino colombiano
César Polanco Tapia1 & Diana Carolina Grande Pulido2
1Universidad Distrital Francisco José de Caldas. Correspondencia:
cpolanco@udistrital.edu.co
2Universidad Distrital Francisco José de Caldas.
cedrella@gmail.com
Recepción: Mayo 27 de 2009/Aprobación: Septiembre 5 de 2009
RESUMEN
En este estudio se hizo la relación de las características madereras encontradas con aspectos ecológicos derivados de un estudio anterior de caracterización, estructura y composición florística practicado en la Reserva Forestal Cárpatos, aledaña al sitio de obtención de las muestras xilológicas. Esta comparación muestra una marcada relación entre las características anatómicas microscópicas de las especies vegetales con el tipo de asociación al que pertenecen. Así, por ejemplo, caracteres como la forma de agrupación, el tipo de perforación del elemento del vaso y punteaduras intervasculares; la altura, número de células de ancho, estratificación y tipo de células que conforman el radio y tipo de fibras, muestran tendencias de mayor grado de evolución en las especies exclusivas de la Asociación Ocoteo callophylla-Weinmannietum pinnatae (OW), mientras que los mismos caracteres demuestran menor tendencia de evolución en las especies exclusivas de la Asociación Clusio multiflorae-Weinmannietum balbisianae (CW). El nivel de evolución de la madera es determinado por medio de un índice que proponen los autores, basado en la experiencia propia y en referencias bibliográficas como Carlquist (2001) y León (2001), y se argumenta su relación con las asociaciones vegetales encontradas en el área deestudio.
Palabras clave: índice evolutivo, ecoanatomía, anatomía comparativa de la madera, asociación vegetal, xilología.
ABSTRACT
In this paper we conducted a relationship of the timber found with ecological aspects arising from an earlier study of the characterization, structure and floral composition practiced in the Cárpatos Forest Reserve, bordering the site from which wood samples were obtained. This comparison shows a strong link between the microscopic anatomical characteristics of vegetal species with the type of association to which they belong. For example, characters such as the grouping form, the type of perforated and intervessel pits, height, number of cells wide, stratification and type of cells that make up the radio and type fiber, show a greater degree of change in the exclusive species of Ocotea callophylla-Weinmannietum pinnatae (OW) association, while the same characters show minor changes in the exclusive species of Clusia multiflorae - Weinmannietum balbisianae(CW) association. The evolution of the timber was identified through an index that suggests the research based on references like Carlquist (2001) and León (2001), and its relationship with the plant associations found in the study area is argued.
Keywords: evolutive index, ecoanatomy, comparative wood anatomy, vegetal association, xilology.
RESUMO
Neste estudo pesquisou-se a relação das características das madeiras encontradas com aspectos ecológicos derivados de um estudo anterior de caracterização, estrutura e composição floral praticado na Resrva Florestal Cárpatos, vizinha ao lugar de obtenção das amostras xilológicas. Esta comparação mostra uma marcada relação entre as características anatômicas microscópicas das espécies vegetais com o tipo de associação à qual pertencem. Assim, por exemplo, caracteres como a forma da agrupação, o tipo de perfuração do elemento do vaso e punteaduras intervasculares; a altura, número de largura das células, extratificação e tipo de células que conformam o rádio e o tipo de fibras, mostram tendências de maior grau de evolução das espécies exclusivas da Associação Ocoteo callophylla-Weinmannietum pinnatae (OW), enquanto que os mesmos caracteres demonstram menor tendência de evolução nas espécies exclusivas da Associação Clusio multiflorae-Weinmannietum balbisianae (CW). O nível de evolução da madeira é determinado por meio de um índice que propõe os autores, baseado ne experiência propria e em referências bibliográficas como Carlquist (2001) e León (2001), e se argumenta sua relação com as associações vegetais encontradas na área de estudo.
Palavras chave: índice evolutivo, eco-anatomia, anatomia comparativa da madeira, associação vegetal, xilogia.
INTRODUCCIÓN
La amplia diversidad biológica presente en Colombia plantea un mosaico de ecosistemas que sustentan el 10% de las especies conocidas de la tierra (Cuéllar-Moyano 2008). Éstas adoptan una distribución y comportamientos específicos, relativos a los procesos evolutivos particulares y a fenómenos ambientales que suceden en cada región; es evidente entonces la necesidad de conocer si existe alguna relación entre los factores biofísicos del entorno y algunas características anatómicas de la madera para apreciar la influencia de dichos facto-res sobre la conformación actual de los elementos constitutivos de las mismas.
Si bien se han realizado amplios estudios acerca de la composición florística, estructura, diversidad y ordenación de la vegetación en algunos bosques altoandinos colombianos, aún no existen estudios en los que se correlacione esta información con la configuración anatómica celular; sólo se conoce que los elementos anatómicos de una especie dependen de ella misma y del medio donde se desarrolle (Rana et al. 2008). Partiendo de esta premisa se supone una relación directa entre las especies que crecen en las mismas asociaciones vegetales, como lo explican León & Espinoza de Pernía (2001), quienes enfatizan que algunos de los principales tipos de variación que ocurren en los elementos anatómicos de la madera se dan cuando hay cambios relacionados con la cantidad de agua disponible, latitud, altitud y características del suelo.
Para reforzar esta hipótesis fue necesario retomar estudios básicos, a partir de los cuales se puede profundizar en el conocimiento de las relaciones a nivel celular entre las especies vegetales que pertenecen a una misma asociación e inclusive entre las diferentes asociaciones, identificando índices que pudieren explicar la causa de agregación de dichas especies.
Considerando lo planteado, este trabajo propone el uso de un nuevo índice que puede ayudar a explicar la conformación y agregación de especies en los bosques de acuerdo a las características anatómicas de la madera y plantear así la ausencia o presencia de un desarrollo adaptativo diferente entre el conjunto de especies arbustivas y arbóreas presentes en una misma zona y pertenecientes a una misma asociación vegetal o entre las que pertenecen a diferentes asociaciones. Para ello se tomó como punto de partida la investigación “Diversidad y caracterización florística, estructural y ordenación de la vegetación arbórea en la Reserva Forestal Cárpatos (Guasca, Cundinamarca)”, realizada por Cantillo et al. (2004) y el “Estudio anatómico comparativo de la madera de 40 especies de bosque altoandino, Guasca (Cundinamarca) ” realizado por Grande & Polanco (2007).
Se recalca que la propuesta metodológica aquí presentada, especialmente en la parte evolutiva, tiene que ver exclusivamente con los elementos morfológicos anatómicos de la madera que han sido tradicionalmente usados para tal propósito, y en ningún momento pretenden reemplazar, revaluar o imponerse sobre metodologías últimamente desarrolladas para la clasificación y relacionamiento filogenético, como es el caso de los métodos cladísticos moleculares como los presentados por Olano (2007).
ANTECEDENTES
COMPOSICIÓN DE LA VEGETACIÓN OBJETO DE ESTUDIO
A partir de los resultados mostrados por Cantillo et al. (2004), donde se estudió ecológicamente el componente boscoso de la Reserva Forestal Cárpatos, mediante un enfoque florístico y fisionómico y algunas variables medioambientales representa tivas, se obtuvo la clasificación de la vegetación en dos asociaciones nuevas, a saber: Ocoteo callophyllae-Weinmannietum pinnatae (OW) y Clusio multiflorae-Weinmannietum balbisianae (CW), las cuales mostraron un gradiente diferencial en sus características florísticas y estructurales, debido sobre todo al gradiente altitudinal. Se concluye que la asociación Ocoteo callophyllae-Weinmannietum pinnatae presenta un mayor desarrollo fisionómico dado que las variables altura, cobertura, estructura diamétrica y área basal así lo confirman. Por su parte la asociación Clusio multiflorae-Weinmannietum balbisianae se caracterizó por presentar un mayor número de individuos y valores altos de cobertura relativa en los estratos menores donde se hace relevante la presencia de palmas de las especies Geonoma weberbaueri y Ceroxylum andicola, esta última más evidente en estratos más elevados.
De acuerdo a Marquínez (2002), las palmas tienen una aparición terrestre más reciente que la mayoría de las angiospermas. No obstante, este estudio considera que la ausencia de crecimiento secundario pone las características anatómicas evidentes en los estípites de las palmas, en un estadio evolutivo menor con relación a las angiospermas, dado el índice propuesto.
ECOANATOMÍA Y ANATOMÍA EVOLUTIVA
En cuanto a los temas concernientes a ecoanatomía y anatomía evolutiva, es muy escasa la bibliografía que se encuentra referente a los bosques altoandinos (Pérez 1989); los estudios llevados a cabo en este tema son referentes a otras zonas de vida y se han realizado en su totalidad en el exterior, Oskolski & Jansen (2009) y Gruber et al. (2008) dan cuenta de ello. Al respecto, León & Espinoza de Pernía (2001) explican el significado de la ecoanatomía y algunas tendencias de las relaciones ecología-anatomía, que han sido identificadas en diversos estudios.
Según estos autores, “Las condiciones ambientales tienen influencia directa sobre la estructura de la madera, pudiéndose encontrar especies que muestran una alta variabilidad, por ejemplo en cuanto a tipo de porosidad, debido a la variación ambiental”. La ecoanatomía de la madera se fundamenta en las correlaciones que existen entre factores ambientales y algunas características anatómicas, las cuales se pueden considerar como estrategias adaptativas. En este sentido, Barajas-Morales (1985) compara las características anatómicas del leño de las especies que crecen en dos tipos de bosques (húmedo tropical y deciduo tropical) que difieren principalmente en la cantidad de humedad disponible a través de todo el año. En las últimas décadas, los estudios de relación ecología-anatomía de maderas se han orientado hacia el conocimiento de la variación estructural de especies, subfamilias, géneros o familias, las cuales crecen en nichos ecológicos diferentes, como lo referencian Araque & León (2006), Lens et al. (2009), Arias & Terrazas (2001) y Aguilar-Rodríguez, Terrazas & López-Mata (2006). Estas investigaciones también han contemplado el estudio de la anatomía en la flora leñosa de áreas completas (Baas et al. 1983, Baas & Carlquist 1985, Moglia & Gimenez 1998) o de acuerdo al hábito de las especies (Baas & Schweingruber 1987). Mediante este tipo de trabajos se ha logrado establecer una serie de tendencias relacionadas con las variaciones de la estructura anatómica de la madera ante cambios en las condiciones ecológicas de las zonas en donde se desarrolla la planta, sólo a nivel general; es decir que estos resultados no constituyen reglas que se puedan generalizar para predecir rasgos anatómicos para otras especies o grupos de especies ante diferentes variaciones ambientales en otros nichos aún desconocidos.
En los estudios de ecología y anatomía de maderas se hace gran énfasis en las características cuantitativas de los elementos vasculares como el número de barras por platina de perforación escaleriforme, diámetro de los poros, número de poros por mm2, longitud de los elementos vasculares y el número de poros que se presentan en los grupos múltiples (León 2001 y 2002).
Este último autor menciona también algunos caracteres que se han tenido en cuenta para el diagnóstico filogenético asociado a la anatomía de la madera. En tal sentido, la ausencia de vasos y fibras, la presencia de células de gran longitud, la ausencia de estratificación, la presencia de barras abundantes en perforaciones, punteaduras escaleriformes en los vasos, fibrotraqueidas, radios homocelulares de células rectas en conjunto con radios multiseriados con extremos uniseriados, parénquima axial apotraqueal difuso y parénquima axial en serie, son indicativos de bajos niveles de especialización. Algunos referentes que lo sustentan pueden consultarse en León & Espinoza de Pernía (1999) y Oskolski & Jansen (2009). A diferencia de las tendencias ecoanatómicas, los diagnósticos filogenéticos asociados a la anatomía de la madera guardan ciertas relaciones como las ya mencionadas, que se mantienen constantes de forma independiente a los gradientes geográficos. No obstante, es de vital importancia no aislar estos dos elementos, ecoanatomía y filogenia, para un mejor entendimiento e interpretación de las manifestaciones xiloanatómicas de forma particular y agregada. Al respecto Baas & Miller (1985) acuñan el término “anatomía ecofilética” para definir el estudio simultáneo de factores ambientales, atributos anatómicos de la madera, estrategias adaptativas y otros mecanismos acerca del origen de la diversidad anatómica en el curso de la evolución, a partir de hipótesis inspiradas por diferentes autores.
OBJETIVO
Este estudio tiene por objeto determinar, a partir de un análisis comparativo de las propiedades anatómicas cualitativas del leño de 40 especies, la existencia de elementos ecoanatómicos y filogenéticos tendenciales para dos asociaciones vegetales pertenecientes a un mismo ecosistema altoandino colombiano.
METODOLOGÍA
DESCRIPCIÓN DEL ÁREA DE ESTUDIO
El área de estudio se encuentra aledaña a la Reserva Forestal Cárpatos, al extremo oriental de municipio de Guasca, Cundinamarca, en el sitio llama-do Potreritos, ubicado al límite sur de Guatavita y al occidente de Junín. Se ubica entre los 2600 y 3000 msnm. La temperatura media anual, reportada por Corpoguavio (2000), citado por Cantillo et al. (2004), es aproximadamente de 12° C. La precipitación media anual es de 1772 mm. Se presentan lluvias en todos los meses del año, con mayor intensidad entre los meses de abril a septiembre y una época más seca en diciembre y enero. Los valores máximos se presentan entre junio y julio y los mínimos en enero. Según la clasificación de Zonas de Vida de Holdridge (1979), este territorio está localizado en la zona de vida bosque húmedo montano bajo (bh-MB).
RECOLECCIÓN DE MUESTRAS
Se realizó mediante la visita a diversas veredas aledañas a la Reserva Forestal Cárpatos en el municipio de Guasca, Cundinamarca, donde se contó con la ayuda de un reconocedor experto en identificación dendrológica, quien a su vez actuó como motosierrista. Se contó con la colaboración de los propietarios de las fincas para la obtención e identificación de las muestras requeridas para este estudio.
De esta manera, se obtuvieron 40 muestras sobre individuos maduros, con dos repeticiones para cada especie, cada una con una longitud de 15 cm y entre 10 y 15 cm de diámetro. Una vez reunido todo el material, se procedió a marcarlo y fotografiarlo para documentar el estado fresco de las muestras. Posteriormente, fueron envueltas en papel periódico y almacenadas en costales de fique para ser transportadas a Bogotá.
La totalidad del material recolectado obedece a tejido maduro de las especies según criterio de los autores, dada la evidencia de hábito de las plantas.
PREPARACIÓN DE PROBETAS
Se utilizaron seis probetas orientadas de acuerdo a los tres planos de la madera para cada especie, cada una de las cuales tenía una sección transversal cuadrada de 2 cm de lado y 2 cm de longitud. Para lograr su hidratación, las probetas fueron colocadas por separado con su repetición en frascos con agua destilada e hipoclorito de sodio al 1% durante un periodo aproximado de dos semanas. El siguiente paso fue el ablandamiento, para lo cual se pusieron a hervir las probetas en agua destilada. Para las maderas más densas, se procedió a hervir las muestras en diferentes lapsos de tiempo hasta que se ablandaron. Para su conservación hasta el momento de sacar los cortes, se mantuvieron en remojo con alcohol al 50% y glicerina en proporción 1:1.
OBTENCIÓN DE CORTES
Se realizaron cortes de cada una de las tres secciones: transversal, tangencial y radial; se obtuvieron con el micrótomo Richter disponible en el Laboratorio de Maderas de la Universidad Distrital. El ángulo de corte de la cuchilla se calibró de acuerdo con la especie, dependiendo de la densidad y del espesor deseado.
COLORACIÓN, DESHIDRATACIÓN Y MONTAJE
Los cortes se mantuvieron en agua destilada hasta el momento de la coloración y el montaje. El procedimiento fue igual para todas las especies y siguió los pasos que se enumeran a continuación:
1. Inmersión de los cortes por un periodo de dos a 10 minutos en fucsina básica
al 1%.
2. Inmersión durante uno a cinco minutos en solución acuosa de ácido
pícrico saturada.
3. Lavado y deshidratado de los cortes, pasándolos sucesivamente en lapsos de 10 minutos
por soluciones de etanol al 25, 50, 75, 100%.
4. Inmersión en solución de etanol-xilol (50 -50%) por diez minutos.
5. Inmersión en xilol puro (100%) hasta el montaje definitivo.
6. Montaje del corte en lámina de vidrio, cubierta con bálsamo de Canadá y
cubreobjeto.
7. Secado de la pieza a 60° C por 24 horas en estufa.
8. Limpieza de la superficie con alcohol.
CARACTERÍSTICAS EMPLEADAS PARA ECOANATOMÍA
Para el presente trabajo se tomaron como elementos básicos para dilucidar ecoanatomía el diámetro de poros y longitud de elementos vasculares, además del número de poros por mm2, en los rangos descritos a continuación:
Cantidad de poros: Pocos (menos de 50), Numerosos (51-200), Muy numerosos
(más de 200).
Tamaño de poros: Muy pequeños (menos de 50µm), Moderadamente
pequeños (50-100 µm), Medianos (100-200µ), Grandes (200 -300 µm),
Muy grandes (más de 300 µm).
Longitud de segmentos vasculares: Cortos (menos de 110 µm), Medianos
(111–800 µm), Largos (más de 800 µm).
CARACTERÍSTICAS ANATÓMICAS EMPLEADAS PARA FILOGENIA
A partir de la descripción microscópica de la anatomía de las maderas estudiadas (Grande & Polanco 2007) y basándose en las características biofísicas del entorno donde se desarrollan, descritas en el estudio de caracterización florística, estructural, diversidad y ordenación de la vegetación arbórea en la Reserva Forestal Cárpatos realizado por Cantillo et al. (2004), se analizó la influencia de dichas condiciones en la configuración anatómica de la madera, apoyados en anteriores estudios relacionados con filogenia. Para esto se analizaron 11 características microanatómicas consideradas de alta heredabilidad y baja afectación medio ambiental según Carlquist (2001), León (2001, 2001 y 2002). De esta manera, se determinaron índices para definir el estado evolutivo de cada especie según las características anatómicas consideradas indicadoras de grado de evolución. Los índices fueron definidos para cada característica de acuerdo a los valores de la tabla 1; el mayor número indica un mayor grado de evolución o especialización. Para cada especie se hace referencia a la característica más evolucionada encontrada a menos que se especifique lo contrario. Así por ejemplo si la especie “X” posee vasos y traqueidas, se tomó la característica vaso para la calificación.
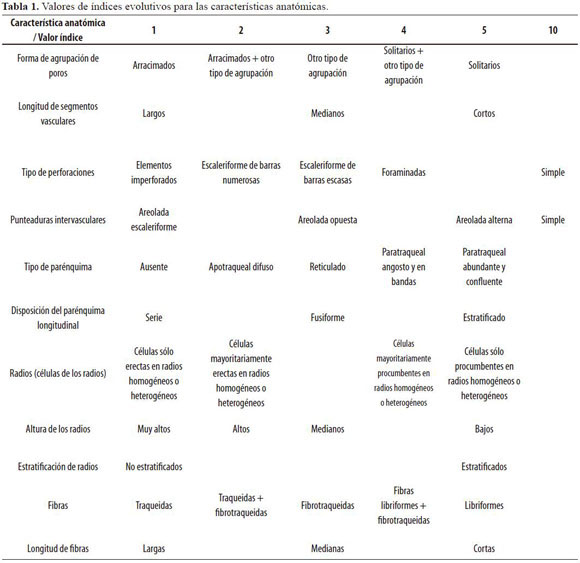
Las clasificaciones al interior de cada característica están direccionadas por el sistema de clasificación de IAWA Committe, obtenido de León & Espinoza de Pernía (2001) de manera modificada para el presente artículo en algunas propiedades. Para los casos “Longitud de segmentos vasculares” y “Altura de los radios” se utilizaron los siguientes rangos:
Segmentos vasculares: Corto (menos de 110 µm), Mediano (111-800 µm),
Largo (más de 800 µm).
Altura de radios: Muy bajos (menos de 200 µm), Bajos (201-500 µm),
Medianos (501-800 µm), Altos (801-2000 µm), Muy altos (más de 2000 µm).
En el caso del parénquima longitudinal, los términos angosto y ancho hacen referencia al número de células en el ancho. Así, por ejemplo, parénquima con tres o menos células en el ancho rodeando el vaso se consideró paratraqueal vascicéntrico angosto; para el caso complementario, se consideró abundante.
ANÁLISIS DE LA INFORMACIÓN
Con base en esta información, se confrontaron las estructuras celulares de las maderas de las especies presentes, utilizando un programa de procesamiento de datos (Excel), que brindó las herramientas necesarias para el análisis, identificando características comunes que las relacionan. De igual manera, se compararon las estructuras de las maderas de las especies de distintas asociaciones, buscando la existencia de diferencias contundentes entre las medianas de los rangos que permitieran asegurar que las estructuras celulares de las especies exclusivas responden a las condiciones particulares de cada asociación, y para comprobar dichas comparaciones se procedió a realizar la prueba no paramétrica de Kruskal-Wallis (Milton & Arnold 2004).
Los datos analizados obedecieron a valores de tipo cualitativo, por lo que para el análisis ecoanatómico no se tienen en cuenta criterios cuantitativos como vulnerabilidad y mesomorfía, los cuales se abordarán de manera detallada y precisa en otra publicación más adelante.
RESULTADOS
ELEMENTOS ECOANATÓMICOS
Con relación a la cantidad de poros se encontró que para la asociación OW el 63.6% de las especies presentan pocos poros, el 36.4% restante corresponde a poros numerosos. En contraste, el 14.3% de las especies exclusivas de la asociación WC presentan pocos poros y el 57.1% poros numerosos.
Con relación al tamaño de poros, la asociación OW presenta un 54.5% de poros moderadamente pequeños, un 36.3% de poros muy pequeños y un 9.1% de poros medianos. En este orden, la asociación WC presentó un 28.65% de poros moderadamente pequeños, un 42.9% de poros muy pequeños y ausencia total de poros medianos. Se recalca que esta última asociación incluye una especie exclusiva que no tiene elementos vasculares sino imperforados, tipo traqueida (Drimys granadensis).
Concerniente a la longitud de los elementos vasculares para OW, en el 100% de las especies se da longitud mediana. Para la asociación WC el 42.9% de los elementos son medianos y el 28.6% se clasifican como largos.
DETERMINACIÓN DE ELEMENTOS INDICADORES DE EVOLUCIÓN
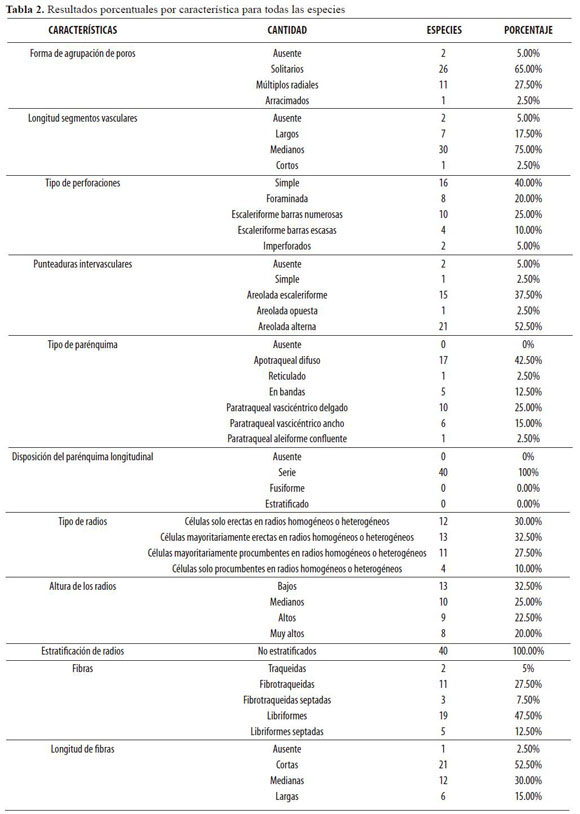
Los resultados presentados en esta sección abarcan todas las especies objeto de estudio; en ella se muestra, primero, la cantidad de especies que presentan la característica y el porcentaje correspondiente (Tabla 2); luego se expone el cuadro de índices evolutivos (Tabla 3) creado para determinar la tendencia evolutiva de las especies estudiadas.
Vasos
La forma de agrupación de los poros predominante para el total de las especies estudiadas fue: solitarios con un 65% de las especies, seguida por múltiplos radiales que corresponde al 27.50% y poros arracimados con una sola especie que corresponde al 2.5%. La longitud de los segmentos vasculares está entre 51 y 800 µm en un 75% de las especies estudiadas; éstos se clasificaron como medianos.
Perforaciones
El 40% de las especies presenta perforación simple, luego se encuentra la perforación escaleriforme de barras numerosas con un 25% y no muy lejos están la perforación foraminada y la perforación escaleriforme de barras escasas con porcentajes de 20 y 10% respectivamente. Las punteaduras intervasculares son en su mayor parte areoladas alternas con un 55%; el 37.5% presenta punteaduras areoladas escaleriformes y las punteaduras areoladas opuestas y simples tan sólo representan el 2.5% cada una.
Parénquima
El mayor porcentaje pertenece al parénquima apotraqueal difuso que corresponde al 42.5% de las especies, después se encuentra el parénquima paratraqueal vascicéntrico angosto con un 25%, y los menores porcentajes son presentados por los parénquimas paratraqueal aleiforme confluente y reticulado, cada uno con un 2.56%.
Radios
De los radios según Kribs predominaron los que presentan células mayoritariamente erectas en radios homogéneos o heterogéneos con un 32.5%, seguidos de cerca por los que presentan células sólo erectas en radios homogéneos o heterogéneos con un 30% y después por los de células mayoritariamente procumbentes en radios homogéneos o heterogéneos con un 27.5%.
De la misma forma, se encontró un 32.5% de las especies con radios bajos, que se encuentran en el rango de 201 a 500 µm. Seguido de ellos se encontraron los radios medianos con un 25%, los altos con un 22.5% y los muy altos representando el 20% de las especies.
Fibras
La mayoría de las especies estudiadas, el 47.5%, mostraron fibras libriformes, y las Fibrotraqueidas ocuparon el segundo lugar en cantidad con 27.5%. En cuanto a longitud, se observó que las fibras cortas predominaron con un porcentaje de 52.5%, seguidas por las fibras de longitud mediana con un 30%.
GRADO DE EVOLUCIÓN POR ESPECIE A PARTIR DE CARACTERES XILOANATÓMICOS
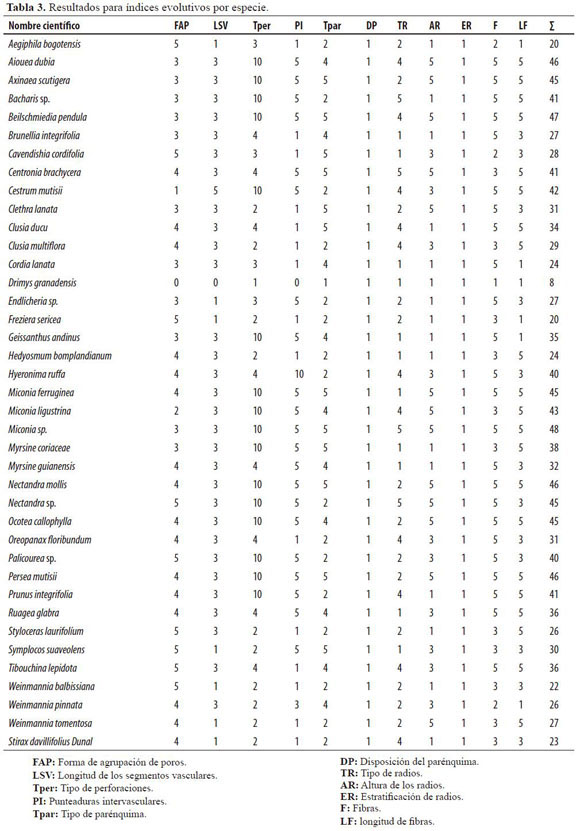
La tabla 3 expone los resultados agregados por especie para las características que son relevantes como indicadores de evolución, según la calificación definida en la metodología. La sumatoria (columna final), expresa el grado de especialización de cada especie; el menor valor representa menor especialización y el mayor valor, mayor grado de especialización.
COMPARACIÓN ENTRE LA EVOLUCIÓN DEL LEÑO Y LA ESPECIALIZACIÓN FLORAL
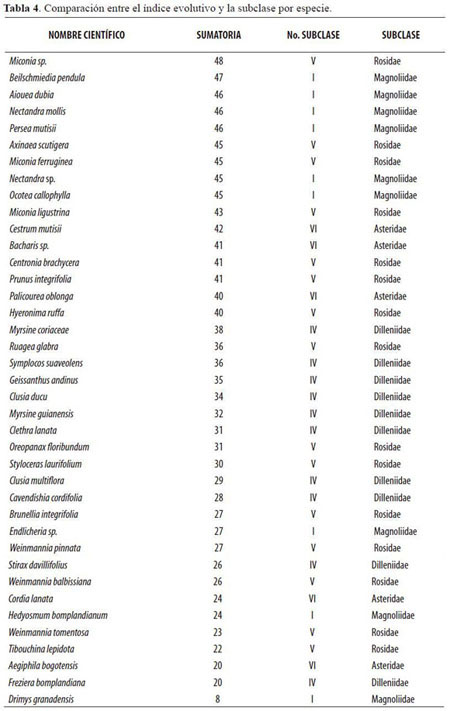
Tomando como punto de partida la información contenida en la tabla 3, las especies se ordenaron desde la mayor calificación (Miconia sp., Beilschmiedia pendula), hasta la menor (Drimys granadensis), basándose en las clasificaciones por subclase reportadas por Judd et al. (1999) y Cronquist (1981). En la tabla 4 se consignan los resultados por especie.
COMPARACIÓN DEL GRADO DE EVOLUCIÓN DE LA ESTRUCTURA XYLOANATÓMICA ENTRE ASOCIACIONES
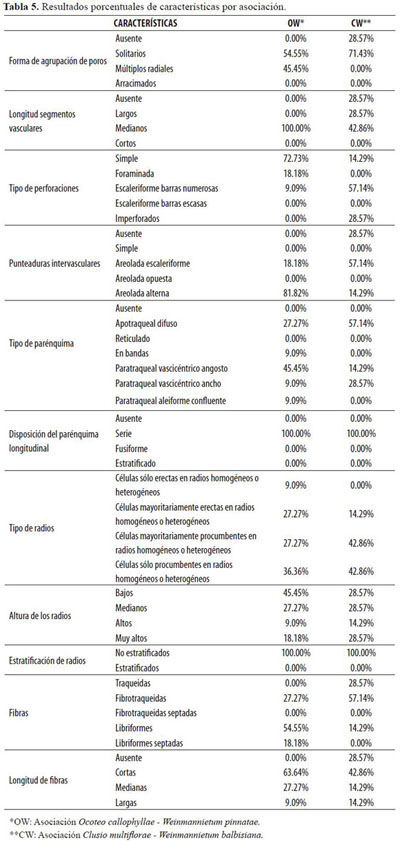
Los resultados presentados a continuación corresponden a las 11 características heredables que son representativas de la madera de las especies del bosque altoandino de Guasca, Cundinamarca (Tabla 5 y 6), para cada asociación encontrada. De la tabla 5 se puede destacar lo siguiente:
Vasos
El tipo de perforación predominante para OW es simple, siendo Styloceras laurifolíum la única especie de la asociación que presenta perforación escaleriforme; en contraste, en WC el tipo de perforación escaleriforme de barras numerosas es el más abundante con un 57.14% de aparición, seguido por los imperforados con un 28.57%. Las punteaduras intervasculares para OW son en su mayor parte aeroladas alternas con un 81.82%, y en las especies restantes 18.18% son areoladas escaleriformes; en cambio para WC las punteaduras intervasculares son en su mayor parte aeroladas escaleriformes con un 57.14% de aparición.
Radios
Los radios de las especies de la Asociación OW son en su mayoría de células sólo procumbentes en radios homogéneos o heterogéneos con un 36.36%, seguidos por los radios con células mayoritariamente erectas en radios homogéneos o heterogéneos y radios con células mayoritariamente procumbentes en radios homogéneos o heterogéneos con un 27.27% cada una. La Asociación WC supera en porcentaje los radios con células sólo procumbentes en radios homogéneos o heterogéneos y radios con células mayoritariamente procumbentes en radios homogéneos o heterogéneos, ya que se presentan cada uno en un 42.86% de los casos.
Fibras
Las fibras de la asociación OW son mayormente libriformes, en un 54.55% de las especies; las fibrotraqueidas aparecen en un 27.27%, y las demás especies, que corresponden al 18.18%, presentan fibras libriformes septadas, en la asociación no aparecen las fibrotraqueidas septadas ni traqueidas. En contraste, para WC la mayoría son fibrotraqueidas, en un 57.14% de las especies, seguidas por las especies con traqueidas con un 28.57%. La longitud de las fibras en OW representan un 63.64% para fibras cortas; se encuentran también un 27.27% con fibras medianas y las especies restantes tienen fibras clasificadas como largas; de igual manera, para WC la longitud de las fibras en el 42.86% de los casos son cortas.
Parénquima longitudinal
En la asociación OW predomina el parénquima vascicéntrico angosto en un 45.45% de las especies, seguido del parénquima apotraqueal difuso en un 27.27%; sucede lo contrario en la asociación CW, donde predomina el parénquima apotraqueal difuso en un 57.14% de las especies en tanto el parénquima vascicéntrico angosto se encuentra apenas en el 14.29% de los casos.
ANÁLISIS NO PARAMÉTRICO PARA EL GRADO DE EVOLUCIÓN DE LOS CARACTERES XYLOANATÓMICOS
Las dos pruebas de Kruskal-Wallis realizadas permitieron comparar simultáneamente las medianas de los rangos para la totalidad de los datos: 40 cuando se tomaron todas las especies y 19 cuando se compararon las especies exclusivas de las dos asociaciones presentes. Uno de los temas que interesan en este trabajo es la existencia de diferencias estadísticas significativas entre los índices de evolución para las especies exclusivas estudiadas en cada asociación.
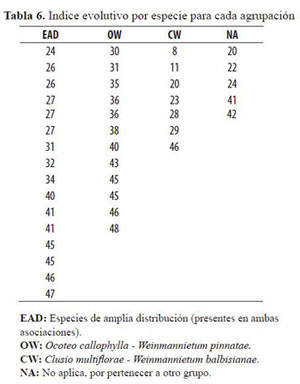
Las variables utilizadas en este diseño fueron los índices evolutivos definidos en la tabla 3. Las tablas 6, 7 y 8 muestran en detalle los datos de origen y el análisis no paramétrico.
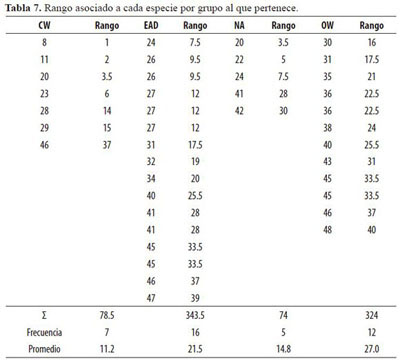

De acuerdo a los resultados desprendidos de la tabla 8, se rechaza la hipótesis nula, es decir que hay diferencias en al menos un par de medianas de las cuatro agrupaciones de especies con una confiabilidad mayor al 97.5% para el caso de las 40 especies. Como las Asociaciones OW y WC presentan especies exclusivas con caracteres anatómicos diferenciados, se practicó una segunda evaluación no paramétrica con n = 19 (12 especies para OW y 7 para CW); los resultados indican diferencias en los caracteres evaluados de las dos asociaciones con un 99% de confiabilidad. La mayor medida de posible evolución la presentan los caracteres anatómicos del leño de las especies exclusivas de la Asociación Ocoteo callophyllae - Weinmannietum pinnatae. La menor medida de evolución la presentan los caracteres anatómicos del leño de las especies exclusivas de la Asociación Clusio multiflorae-Weinmannietum balbisiana.
DISCUSIÓN DE RESULTADOS
ELEMENTOS ECOANATÓMICOS
Los datos arrojados para la longitud del elemento del vaso demuestran que el 75% de las especies estudiadas presentan longitudes de los elementos del vaso medianas y un 17.50% de las especies presentan longitudes largas, en contraste con tan sólo un 2.50% de elementos cortos. Se puede presumir que esta característica se relaciona con la mejora gradual de la eficacia conductora, reflejada en índices de caudal más altos en vasos e índices de corriente más altos de fotosintatos en parénquima axial y del radio, lo cual representa una ventaja selectiva que han desarrollado las especies presentes en ambientes húmedos. Asimismo, Zimmermann (1983), citado por Carlquist (2001), concluyó que los vasos más largos conceden mayor eficacia conductora y los vasos más cortos confieren mayor seguridad en relación con los embolismos (la extensión de los embolismos de aire es frenada por el límite de un vaso). Aunque el fenómeno de embolismo debe tratarse en una dimensión mucho más amplia, que involucre la red interna conductora para todo el sistema denominado árbol, pues según Loepfe et al. (2007) la interacción de punteaduras y elementos conductores longitudinales, denominada por ellos conectividad, afecta a tal punto que una mayor conectividad genera un aumento en la conductividad hidráulica y una mayor vulnerabilidad al embolismo.
Así, la combinación de las características anatómicas ya mencionadas en la asociación OW, junto con la presencia de punteaduras areoladas alternas en la mayoría de sus especies exclusivas (82%), determina una mayor seguridad al disminuir la conectividad de la red, la cual se distingue por pocos poros de tamaño moderadamente pequeños. No sucede lo mismo en la asociación CW, donde la mayoría de las especies exclusivas presentan punteaduras aeroladas escaleriformes (57%), lo cual induciría, ceteris paribus, una menor seguridad. Para fortuna de este último grupo de especies se da una tendencia hacia poros numerosos y muy pequeños, lo cual constituye un elemento paliativo que jalona la seguridad a niveles más altos.
Según Carlquist (2001), el cambio de los poros hacia diámetros más anchos de formas redondeadas es un cambio de la capacidad para manejar un mayor volumen de agua por unidad de tiempo, por vaso; de acuerdo a ello, se puede decir que las 40 especies objeto de estudio se han especializado para manejar mayores volúmenes de agua, ya que el 100% de las especies presentan poros de forma redondeada; sin embargo, el diámetro de los poros no ha cambiado al mismo ritmo, puesto que el 55.26% de las especies presentan un tamaño de poros moderadamente pequeños y un 26.32% presentan poros muy pequeños.
Lo anterior no quiere decir que las especies se especializaron de manera truncada, lo que puede haber ocurrido es que los cambios bruscos de temperatura (12° C) que se dan en la zona, que según Corpoguavio (2000), citado por Cantillo et al. (2004), pueden tener variaciones diarias de +/- 10° C, que condujeron a otra adaptación: ya que los vasos estrechos tienen menor posibilidad de sufrir embolismo por estrés hídrico, se cree (Ellmore & Ewers 1985, Hargrave et al. 1994 citado por Carlquist 2001) que los vasos se han estrechado, sin importar la usual abundancia hídrica de la cual gozan en este ambiente, para evitar este problema. Ni siquiera en situaciones de tensión hídrica se eliminan los vasos estrechos, pues brindan mayor seguridad a los vasos aunque se sacrifique un poco la eficiencia en la conducción.
Además, Erak (1971), citado por Metcalfe (1979), encontró que a mayores altitudes, los vasos son más pequeños y desarrollan paredes más gruesas que los observados en especies que crecen en tierras bajas. Esto quiere decir que aunque el ensanchamiento del vaso es indicativo de especialización según algunos estudios, no se debe considerar como característica universal de la evolución de la madera, puesto que, según Carlquist (2001), una vez que se hayan alcanzado las placas simples de la perforación, el ensanchamiento adicional de vasos es controlado probablemente por factores ecológicos independientes de las tendencias principales de la evolución del xilema.
Esta investigación considera que los individuos estudiados poseen bajas tasas de transpiración porque no existe un alto gradiente de humedad entre la planta y el ambiente que facilite el movimiento de vapor de agua de la planta hacia afuera, esto puede deberse a la elevada humedad relativa promedio de la zona (86%). Asimismo, debido a la baja temperatura de la región (12º C), los individuos usualmente no requieren transpirar como método de enfriamiento. Según León & Espinoza de Pernía (2001), la mayoría de las especies que poseen bajas tasas de transpiración se caracterizan porque su xilema posee numerosos vasos y la mayoría de ellos son de diámetro reducido, como sucede en la mayoría de las especies objeto de este estudio; mientras que las especies con altas tasas de transpiración tienen una baja frecuencia de vasos pero con mayores diámetros.
La cantidad de poros resultó ser mayor para la categoría pocos, con un 57.89%, aunque no dista mucho de la categoría numerosos, con un 42.11%. La frecuencia de poros aumenta, según León & Espinoza de Pernía (2001), cuando se produce un incremento de la sequía o disminuciones de temperatura.
De igual manera, las punteaduras alternas al estar presentes a lo largo y ancho de toda la pared del vaso permiten una comunicación más eficiente y mejor distribución con los elementos vecinos; cuando los vasos son solitarios, esta comunicación puede presentarse entre vasos y fibras, ya que éstas también tienen punteaduras, lo cual supone que las fibras tienen función de almacenaje para situaciones de insuficiencia hídrica, que, para el caso de los vasos con perforación simple, sería una adaptación para evitar el embolismo.
ÍNDICES EVOLUTIVOS PARA TODAS LAS ESPECIES
A continuación se analizan las estructuras anatómicas de las especies estudiadas con base en los resultados obtenidos referentes a su grado evolutivo.
En cuanto al tipo de perforación de los vasos, un 40% de las especies posee platina de perforación simple, lo cual confirma el supuesto alto grado de evolución de estas especies. Frost (1930-1931), citado por León & Espinoza de Pernía (2001), concluye que las platinas de perforación escaleriformes con muchas barras e inclinadas son características primitivas. La especialización conlleva a una reducción progresiva del número de barras hasta llegar a las platinas simples y horizontales.
No muy lejos están las platinas de perforación escaleriformes de barras numerosas con un 25%. Esta característica hace evidente un bajo grado de evolución en estas especies, ya que, según Carlquist (1999), citado por Carlquist (2001), las placas de perforación de especies antiguas tienen a menudo barras numerosas, mientras que los elementos del vaso formados más recientemente tienen platinas simples de perforación; esto demuestra el menor grado de evolución para las especies exclusivas de la asociación Clusio multiflorae-Weinmannietum balbisianae, las cuales presentaron un 57.14% de las platinas de perforación escaleriformes de barras numerosas, y el mayor grado evolutivo de las especies exclusivas de la asociación Ocoteo callophyllae-Weinmannietum pinnatae, las cuales poseen un 72.73% de las perforaciones de tipo simple.
La mayoría de las especies de la asociación CW, el 57.14%, presentaron parénquima apotraqueal difuso, el cual, según Kribs (1937), citado por León & Espinoza de Pernía (2001), es claramente uno de los más primitivos. Igualmente, estos autores afirman para el parénquima paratraqueal, que en la asociación OW suma un 63.64% en sus diferentes tipos, que se encuentra más asociado a caracteres evolucionados que a caracteres primitivos y que, aparentemente, el parénquima aliforme y el confluente se presentan como los más evolucionados. Respecto a esta característica, Carlquist (1992) afirma que el parénquima axial vascicéntrico es un indicativo de especialización, por lo menos en el género Hedyosmum.
De acuerdo con el sistema de clasificación de radios establecido por la Asociación Internacional de Anatomistas de la Madera (IAWA committe), citado por León & Espinoza de Pernía (2001), y comparándola con el sistema de clasificación de Kribs, la condición más primitiva corresponde a los radios heterocelulares y multiseriados con más de cuatro rutas de células marginales en combinación con radios homocelulares y uniseriados constituidos por células cuadradas o erectas. El proceso evolutivo se corresponde con una disminución en la anchura y la altura de radios, así como un aumento de su homogeneidad. La condición más evolucionada corresponde a radios uniseriados y homocelulares, constituidos por células procumbentes.
El estudio registró un 32.5% de individuos con presencia de radios heterogéneos con células en su mayoría procumbentes, es decir que según los radios el grado de evolución de dicho grupo es medianamente alto; sin embargo, se encontró otro grupo, que corresponde al 30% de las especies estudiadas, con radios heterogéneos de células en su mayoría erectas, es decir que tienen un grado de evolución medianamente bajo. En cuanto a las especies con células homocelulares y erectas en su totalidad, sumaron un 25%; esta característica expone una condición de baja evolución y está asociada a radios multiseriados y tipo roble. En contraste se encontró un 12.5% de las especies con células homocelulares de células procumbentes, lo que evidencia un proceso evolutivo mucho más avanzado.
León & Espinoza de Pernía (2001) señalan que la tendencia evolutiva correspondiente al tejido de soporte en el xilema de maderas dicotiledóneas se inicia con traqueidas, para luego pasar a fibras tipo fibrotraqueidas y, finalmente, a fibras libriformes. De acuerdo a esta afirmación, el presente estudio posee pocas especies con este rasgo primitivo puesto que sólo se encontró un 5% de las especies con traqueidas en CW, y en cambio un porcentaje considerable (35%) de fibrotraqueidas, el cual se ubica mayoritariamente en la asociación CW. Esto evidencia un grado medio de evolución, sin embargo, en OW es mucho más alta la cifra de especies con fibras libriformes (54.55%), lo que demuestra un alto grado de evolución en la mayoría de sus especies.
La evidencia circunstancial: en Carlquist (1984), citado por Carlquist (2001), sugiere que tanto las fibrotraqueidas como las fibras libriformes son células no conductivas y demuestra que las familias con traqueidas demuestran siempre vasos solitarios, y aunque existen excepciones en todas las especies que presentan traqueidas en este estudio, en Drimys granadensis, por ejemplo, se cumple dicha afirmación. De otro lado, las familias con fibrotraqueidas y fibras libriformes pueden tener los vasos solitarios o vasos agrupados, dependiendo del grado de aridez que caracteriza el hábitat particular de una especie.
Lo anterior explica que los individuos estudiados presenten vasos solitarios asociados a fibras libriformes y a fibrotraqueidas sin presencia de traqueidas vasculares, debido a la disponibilidad constante de agua en la zona de estudio que hace innecesarios elementos conductores adicionales. Asimismo, Carlquist (2001) afirma que aunque la especialización de los elementos imperforados en una madera dada no están siempre en sincronía con el estado evolutivo de los elementos del vaso, se puede decir que éstos cambian hacia conductos más eficientes, mientras que los elementos imperforados siempre se llegan a adaptar más como elementos mecánicos, conformando así una efectiva división del trabajo para el primer caso.
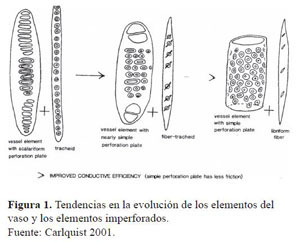
Como se puede observar en las descripciones, haciendo una relación entre los individuos que presentan perforaciones escaleriformes se encuentra que un 78.57% de ellos están asociados a fibrotraqueidas y a traqueidas. De igual manera, un 87.5% de las especies que presentan perforaciones simples y foraminadas se asocian a fibras libriformes. Lo anterior demuestra lo descrito por Carlquist (2001) en un esquema donde se muestra la relación evolutiva entre perforaciones y tipos de fibra (Figura 1). Este esquema se basa en la afirmación de que en una madera primitiva un elemento del vaso con una placa de perforación escaleriforme tenderá a ser asociada a un traqueida, mientras que en una madera evolucionada un elemento del vaso con placa de perforación simple tenderá a ser asociado a fibras libriformes.
En cuanto a la longitud de las fibras, según anteriores estudios (Carlquist 1984 citado por Carlquist 2001) se observa una reducción en la longitud de las fibras. El presente estudio encontró que el 52.5% de las especies estudiadas presentan fibras de longitud corta y el 71.43% tienen presencia de fibras libriformes.
La relación de índices (Tabla 3) muestra claramente, mediante la sumatoria de los resultados para cada característica, una indicación del estado evolutivo de cada especie. Así, se observa que la especie con menor calificación es Drimys granadensis y obedece principalmente a la ausencia de poros que son reemplazados por traqueidas que realizan la función de conducción debido a la densidad de punteaduras areoladas que posee.
Los datos disponibles en Carlquist (1984), citado por Carlquist (2001), señalan que la presencia de traqueidas son un mecanismo más eficaz para mantener caminos conductores con relación a la agrupación de los vasos, puesto que lo último ocurre solamente en maderas cuyos elementos imperforados son no conductivos.
Siguiendo con el análisis, se encontró un grupo de especies que presentan un alto índice evolutivo: Nectandra mollis, Beilschmiedia pendula, Ocotea callophylla, Nectandra sp., Persea mutisii y Aiouea dubia; estas especies presentan una particularidad, y es que todas, aunque obtuvieron valores altos de índice evolutivo, se encuentran clasificadas en la subclase Magnoliidae, que es la subclase más antigua de todas las familias de angiospermas presentes en este estudio. Para explicar este fenómeno es necesario recordar que la clasificación en subclases fue determinada de acuerdo a la especialización de las flores; en contraste, las especies mencionadas no tienen flores especializadas puesto que se encuentran en la parte alta del dosel del bosque y poseen estrategias de dispersión de polen por viento (anemofilia), lo cual puede significar que una mayor especialización del leño no va a la par con una alta especialización floral. Los estudios cladísticos podrían explicar mejor este fenómeno.
Por el contrario, el grupo de las especies pertenecientes a la subclase Asteridae (que es la más evolucionada de las angiospermas) obtuvo valores bajos de evolución del leño, aunque no en extremo, en la calificación de índices evolutivos. Esta situación, hipotéticamente, se debe al fenómeno explicado con anterioridad. De esta manera, se puede decir que las especies Stirax davillifolius, Palicourea oblonga, Cestrum mutisii, Cordia lanata, Bacharis sp. y Aegiphila bogotensis, tienen una notable especialización de las flores, muy probablemente debido a la posición media en el estrato arbóreo que hace necesario implementar estrategias de dispersión de polen que utilicen flores vistosas para atraer insectos y aves; por tanto, la evolución fisiológica de la madera se ha quedado rezagada para estas especies que no alcanzan grandes alturas y no necesitan características que aseguren la conducción efectiva de líquidos en su interior a grandes alturas.
COMPARACIÓN DEL GRADO DE EVOLUCIÓN DE LA ESTRUCTURA XYLOANATÓMICA ENTRE ESPECIES DE LAS DOS ASOCIACIONES
Como se observa en la figura 2, la asociación Ocoteo callophyllae-Weinmannietum pinnatae (OW) posee una marcada tendencia a presentar especies exclusivas con perforaciones simples, lo que indica una adaptación de las estructuras para lograr una mayor conducción por vaso, por área, por unidad de tiempo. En contraste, en las especies exclusivas de la asociación Clusio multiflorae-Weinmannietum balbisianae (CW) las perforaciones son en su gran mayoría de tipo escaleriforme, este rasgo hace que la conducción del agua sea menos eficiente y está relacionado con el grado de evolución de la madera de las especies.
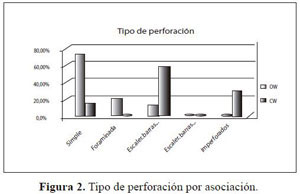
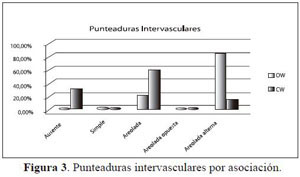
La asociación Ocoteo callophyllae-Weinmannietum pinnatae (OW) presenta punteaduras intervasculares areoladas alternas en un 80% (Figura 3), lo que indica, al igual que las perforaciones del vaso, una adaptación de las especies de esta asociación a una administración hídrica más eficiente, mientras que las especies exclusivas de la asociación Clusio multiflorae - Weinmannietum balbisianae (CW) tienen un comportamiento no proporcional, siendo más frecuente para los rangos medios y bajos de especialización.
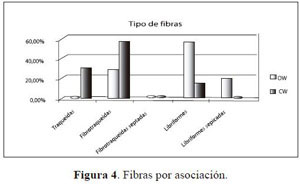
El análisis de las fibras (Figura 4) demuestra que, de igual manera, los caracteres más especializados de la madera se encuentran en las especies exclusivas de la asociación Ocoteo callophyllae - Weinmannietum pinnatae (OW), que para el caso corresponde a las fibras libriformes, y que según Carlquist (2001) suelen asociarse a vasos con perforación simple y punteaduras del vaso alternas. Por el contrario, las especies exclusivas de la asociación Clusio multiflorae -Weinmannietum balbisianae (CW) posee una evidente mayoría de traqueidas y fibras tipo fibrotraqueida.
ANÁLISIS ESTADÍSTICO COMPARATIVO ENTRE ASOCIACIONES
Como se observa en los resultados de la prueba no paramétrica, existe una marcada diferencia entre los índices evolutivos según las características anatómicas para las especies exclusivas de las asociaciones presentes en el área de estudio. Esta diferencia está representada con un 99% de confiabilidad. El grado de evolución de las especies medido por las variables elegidas desciende en el sentido Asociación Ocoteo callophyllae-Weinmannietum pinnatae a Asociación Clusio multiflorae-Weinmannietum balbisianae.
El grupo de especies exclusivas de la asociación Clusio multiflorae -Weinmannietum balbisianae evidencia caracteres anatómicos más primitivos; estas diferencias muy probablemente se deben, se gún el estudio realizado para la zona por Cantillo et al. (2004), a la adaptación diferencial entre las asociaciones, dados los rangos de altitud y posición fisiográfica. Así, Ocoteo callophyllae-Weinmannietum pinnatae se encuentra entre los 2620 y los 2800 msnm, generalmente ubicada en zonas de faldas y/o laderas bajas con pendiente promedio de 55%, mientras que Clusio multiflorae-Weinmannietum balbisianae se encuentra entre los 2755 y los 2920 msnm, por lo general en las cimas de las montañas y/o laderas altas con una pendiente promedio de 75%.
A pesar de estos resultados, se debe probar la metodología en zonas con mayor número de especies como los bosques basales heterogéneos a fin de aumentar el tamaño de la muestra (n) por especie y para especies con el ánimo de obtener una evidencia circunstancial más contundente que la mostrada en el presente trabajo, apoyado además por un fisiólogo vegetal especialista.
CONCLUSIONES
Los resultados porcentuales para las características anatómicas microscópicas por asociación muestran una diferencia evidente entre las especies exclusivas de cada asociación vegetal presente en el área de estudio, debido principalmente a 11 caracteres anatómicos descritos que pueden ser indicadores de evolución o especialización.
A partir del análisis no paramétrico, basado en rangos para variables anatómicas evolutivas propuestas, se encontró una diferencia significativa con un 99% de confiabilidad entre las dos asociaciones, revelando así que el grado de evolución del leño desciende en el sentido Asociación Ocoteo callophylla -Weinmannietum pinnatae a Asociación Clusio multiflorae - Weinmannietum balbisianae.
Las características anatómicas observadas para las especies de este estudio fueron coherentes con las teorías propuestas por diversos autores que han realizado investigaciones sobre evolución en anatomía del leño. De esta forma, se señala que las condiciones ambientales presentes en la zona, así como el comportamiento propio de las especies, expresados en la manera como pertenecen a una u otra asociación, tienen influencia directa sobre la estructura y caracteres considerados como evolucionados en la madera, siendo difícil delimitar la responsabilidad entre filogenia y ecología.
Los índices evolutivos propuestos en este estudio muestran que la especialización del leño guarda una relación irregular con la especialización floral, ya que las especies que obtuvieron mayores valores para evolución del leño se encuentran clasificadas en la subclase más antigua (Magnoliidae) y las especies que obtuvieron menores valores evolutivos en el leño están clasificadas en una subclase aparentemente más evolucionada de flores (Asteridae).
En la actualidad se están aplicando nuevas metodologías de tipo molecular para asignar filogenéticamente las especies vegetales, superando la mera caracterización floral, por lo que aún no está escrita la última palabra al respecto. Los caracteres anatómicos del leño son una propuesta con más de 30 años de evidencia que deben complementarse o sobreponerse con los últimos resultados encontrados; aún falta allanar un largo camino en este sentido.
AGRADECIMIENTOS
A los evaluadores anónimos por sus valiosos aportes para el redireccionamiento de este trabajo y a Heliodoro Sánchez por su paciencia e intermediación para la obtención de información secundaria. A los reconocedores de campo y finqueros que prestaron su ayuda. Agradecemos especialmente a la UDFJC por permitir el desarrollo de las investigaciones precedentes. Al Comité Editorial por su objetividad en las apreciaciones.
REFERENCIAS BIBLIOGRÁFICAS
Aguilar-Rodríguez, S., T. Terrazas, & L. López-Mata. 2006. Anatomical wood variation of Buddeleja cordata (Buddlejaceae) along its natural range in Mexico. Trees 20: 253-261.
Araque, A. & W. León. 2006. Anatomía comparada del leño de Spondias mombin L. (Anacardiaceae) que crece en zonas del banco y bajío de la Reserva Forestal Caparo (Barinas, Venezuela). Revista Forestal Venezolana 50 (1): 9-17.
Arias S. & T. Terrazas. 2001. Variación en la anatomía de la madera de Pachycereus pectenaboriginum (Cactaceae). Serie Botánica 72 (2): 157-169.
Baas, P. & S. Carquilst. 1985. A comparison of the ecological wood and anatomy of the floras of Southern California and Israel. IAWA Bulletin 6 (4): 349-353.
Baas, P. & R. Miller. 1985. Functional and ecological wood anatomy. Some introductory comments. IAWA Bulletin 6 (4): 281-282.
Baas, P. & F. Schweingruber. 1987. Ecological trends in wood anatomy of trees, shrubs, and climbers from Europe. IAWA Bulletin 8 (3): 245-274.
Baas, P., E. Werker & A. Fahn. 1983. Some ecological trends in vessel characters. IAWA Bulletin 4 (2-3): 141-159.
Barajas-Morales J. 1985. Wood structural differences between trees of two tropical forests in Mexico. IAWA Bulletin 6 (1-4): 355-364.
Cantillo, E., A. Avella & K. Rodríguez. 2004. Diversidad y caracterización florística estructural de la vegetación arbórea de la Rserva Forestal Cárpatos, Guasca, Cundinamarca. Revista Colombia Forestal 8 (17): 5-21.
Carlquist, S. 2001. Comparative wood anatomy. Sistematic, ecological and evolutionary aspects of dicotyledon wood. Springer Verlag. Berlín y Heidelberg.
Carlquist, S. 1992. Wood anatomy of Hedyosmum (Chloranthaceae) and the tracheid-vessel element transition. ALISO Rancho Santa Ana Botanic Garden 13 (3): 447-462.
Cronquist, A. 1981. An integrated system of classification of flowering plants. Columbia University Press. Nueva York.
Cuéllar-Moyano, F. 2008. Aves falconiformes de Colombia: Águila y afines. Colombia Curiosa. Bogotá.
Grande, C. & C. Polanco. 2007. Descripción anatómica de la madera de 40 especies del bosque altoandino en Guasca, Cundinamarca. Revista Colombia Forestal 10 (20): 180-206.
Gruber, A., D. Baumgartner, J. Zimmermann & W. Oberhuber. 2008. Temporal dynamic of wood formation in Pinus cembra along the alpine treeline ecotone and the effect of climate variables. Trees 23: 623-635.
Holdridge, L. 1979. Ecología basada en zonas de vida. Editorial IICA. San José, Costa Rica.
Judd, W., C. Campbell, E. Kellogg & P. Stevens. 1999. Plant systematics. Sinauer Associates. Sunderland, Massachusetts.
Lens, F. J. Luteyn E. Smets & S. StevenJansen. 2009. Ecological trends in the wood anatomy of Vaccinioideae (Ericaceae). Flora 199: 309-319.
León, W. 2001. Anatomía del leño, aspectos ecológicos y filogenia en mangles de Venezuela. Revista Forestal Venezolana 45 (2): 191-203.
León, W. 2002. Anatomía xilemática del tronco de Pereskia guamacho (Cactaceae) procedente del Estado de Mérida, Venezuela. Pittieria 31: 41-49.
León, W. & N. Espinoza de Pernía. 1999. Parénquima disyuntivo y filogenia. Revista Forestal Venezolana 43 (2): 137-146.
León, W. & N. Espinoza de Pernía. 2001. Anatomía de madera. Universidad de los Andes. Mérida, Venezuela.
Loepfe, L. J. Martínez-Vilalta, J. Piñol & M. Mencuccin. 2007. The relevance of xylem network structure for plant hydraulic efficiency and safety. Journal of Theoretical Biology 247: 788-803.
Marquínez, X. 2002. Historia evolutiva de las plantas. Universidad Nacional de Colombia. Bogotá.
Metcalfe, C. 1979. Anatomy of the dicotyledons: Systematic anatomy of the leaf and stem, with a brief history of the subject. Vol. I. Oxford University Press.
Milton, J. & J. Arnold. 2004. Probabilidad y estadística con aplicaciones para ingeniería y ciencias computacionales. McGraw-Hill. México.
Moglia, G. & A. Giménez. 1998. Rasgos anatómicos característicos del hidrosistema de las principales especies arbóreas de la región chaqueña argentina. Investigación Agraria 7 (1-2): 53-71.
Olano, C. 2007. Estudios filogenéticos moleculares del género Caryodaphnopsis (Lauraceae) y sinopsis de las especies colombianas. Trabajo de grado para optar al título de Biólogo. Universidad de los Andes. Bogotá.
Oskolski, A. & S. Jansen. 2009. Distribution of scalariform and simple perforation plates within the vessel network in secondary xylem of Araliaceae and its implications for wood evolution. Plant Systematics and Evolution 278: 43-51.
Pérez, A. 1989. Caracterización ecoanatómica del leño de 40 especies del bosque La Mucuy, Estado de Mérida, Venezuela. Revista Forestal Venezolana 33: 43-51.
Rana, R., R. Langenfeld-Heyser, R. Finkeldey & A. Polle. 2008. Functional anatomy of five endangered tropical timber wood species of the family Dipterocarpaceae. Tree 23: 521-529.




Licencia
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






