
DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2006.1.a01Publicado:
01-01-2006Número:
Vol. 9 Núm. 19 (2006)Sección:
Artículos de investigación científica y tecnológicaCaracterización florística y estructural de la vegetación en claros de la parcela permanente de 25ha en la reserva natural La Planada (Nariño)
Floristic and structural characterization of the vegetation in clearings of the permanent plot of 25 ha in the La Planada Natural Reserve (Nariño)
Palabras clave:
Claros, selvas subandinas, bosques de niebla, La Planada, Nariño, Colombia (es).Descargas
Referencias
Andrade, 1. G. 1993. Paisaje y biodiversidad en las selvas de los Andes. Pp. 31-47. En: Germán Andrade (ed.). En Carpanta, selva nublada y páramo. Ecología y conservación de un sistema alto andino.
Asquith, N. 2002. La dinámica del bosque y la diversidad arbórea. Pp. 377-406. En: Guariguata M. y Kattan G. (compiladores). Ecología y conservación de bosques neotropicales. Ediciones Lur. Costa Rica, 691 p.
Brokaw, N. 1985. Treefalls, regrowth, and comunita structure in tropical forest. En: The ecology of natural disturbance and patch dynamics. S.T. Pickett and P.S. White. Academic Press. Orlando, Florida.
Condit, R. 1998. Tropical Forest Census Plots: methods and results from BCI, Panamá and comparison with other plots. Springer-Verlang, 211 p.
Damascos A. y Rapaport, E. 2002. Diferencias de la flora herbácea y arbustiva entre claros y áreas bajo dosel en un bosque de Notofagus pumilio en Argentina. Departamento de Ecología. Universidad Nacional del Comahue, 8400 Bariloche, Argentina.
Denslow, J. 1980. Grap partitioning among tropical rainforest trees. Biotropica Supl. to 12(2): 47-55
Duellman, W.E. 1982. Comprensión climática cuaternaria de los Andes. Efecto sobre la especiación. Pp 77-201. En Salinas, P.J. (De) Zoología Neotropical. Acta VII Congreso Latinoamericano de Zoología. Mérida Venezuela.
FES. 2001. Plan de Manejo Reserva Natural La Planada 2001-2010. Fundación FES, Reserva Natural la Planada. Bogotá, Colombia,
Gentry, A.H. 1993. Vistazo general a los bosques nublados andinos y a la flora de Carpanta. Pp. 67-79. En: Germán Andrade (ed). En Carpanta, selva nublada y páramo. Ecología y conservación de un sistema alto andino.
Gentry, A.H. 1995. Pattems of diversity and floristic composition in neotropical montane forest. Biodiversity. Biodiversity and Conservation of Neotropical Montane Forest. Pp. 103-126.
Henderson, A., S.P. Churchill & J. Luyeyn. 1991. Neotropical plant diversity. Nature 351: 21-25.
Hogan K. y J. L. Machado, 2002. La luz solar: consecuencias biológicas y medición. En: Guariguata M. y Kattan G. (compiladores). Ecología y conservación de bosques neotropicales. Ediciones Lur. Costa Rica, 691 p.
Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. 2000. Colombia inegadiversa: cinco arios explorando la riqueza de un país biodiverso. Bogotá: Instituto Humboldt. Pp. 57-61.
Lamprecht, H. 1990. Silvicultura en los trópicos: los ecosistemas forestales en los bosques tropicales y sus especies arbóreas, posibilidades y métodos para un aprovechamiento sostenido. GTZ. República Federal Alemana.
Lieberman, M., D. Lieberman, R. Peralta & G. S. Hartshorn. 1995. Canopy closure and the distribution of tropical forest tree species at La Selva, Costa Rica. Joumal of Tropical Ecology 11: 161-178.
Louman B., Quirós D. y M. Nilsson. 2001_ (Editores). Silvicultura de bosques latifoliados húmedos con énfasis en América Central. Serie Técnica, Manual técnico 46. CATIE. Turrialba, Costa Rica, 265 p.
Mendoza, C. y B. Ramírez. 2000. Flora de La Planada: Guía ilustrada de Familias y Géneros. Instituto Alexander von Humboldt, Fundación FES, WWF, Bogotá.
Mendoza, H. y Ramírez, B. 2001. Dicotiledóneas de La Planada, Colombia: lista de especies. Biota Colombiana: V. 2, N° 1, septiembre de 2001.
Rangel O. y Velásquez, A. 1997. Métodos de estudio de la vegetación. En: O. Rangel, Lowy. P. y Aguilar. M. Colombia Diversidad Biótica, Tomo II: Tipos de vegetación en Colombia. Instituto de Ciencias Naturales Universidad Nacional de Colombia. Bogotá.
Vallejo, M. I. 2000. Estructura, demografía y dinámica del bosque Andino. Instituto de Investigaciones Biológicas Alexander von Humboldt. Documento interno, s.p.
Vallejo, et al. 2004. Tropical forest diversity and dynamism: Findings from a Large-Scale Plot Metwork. University of Chicago Press.
Whitmore T. 1989. Canopy gaps and the two major groups of forest trees. Ecology 70: 536-538.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Artículos de investigación científica y tecnológica
Colombia Forestal, 2006-11-00 vol:9 nro:19 pág:5-30
Caracterización florística y estructural de la vegetación en claros de la parcela permanente de 25 ha en la Reserva Natural la Planada (Nariño)
Adriana Rincon Velásquez
Max Alejandro Triana Gómez
Javier Dario Burgos
Universidad Distrital Francisco José de Caldas
Resumen
Las perturbaciones naturales del bosque promueven la diversidad biológica y estructural. Nosotros evaluamos los efectos de los claros sobre la estructura y composición de la vegetación, en la parcela perm' anente de 25 ha de La Planada. Con los datos del Instituto Alexander von Humboldt (IAvH), analizamos los cambios en dos tamaños de claro y dosel cerrado. Concluimos que no se evidencian cambios significativos en la estructura y composición de especies, género y familias.
Palabras claves:
Claros, selvas subandinas, bosques de niebla, La planada,Nariño, Colombia.
Abstract
The natural disturbances of the forest promote the biological and structure diversity. We evaluated the effects of canopy gaps at the structure and composition in the permanent plots of 25 ha in the La Planada. With the data of the Instituto Alexander von Humboldt (IAvii) we analyzed the changes in two size gaps and closed canopy. We concluded that natural disturbance processes showed slow significant alterations at the structure and composition of species, genus and families.
Key words:
Canopy gaps, neotropical montane forest, La planada, Nariño, Colombia
INTRODUCCIÓN
Los bosques nublados andinos y subandinos se encuentran entre los ecosistemas más complejos y dinámicos en el marco mundial (Duellman, 1982). Debido, entre otras razones, a su topografía, biogeografía e historias de vida representan un mosaico de diferentes comunidades biológicas típicamente caracterizadas por niveles de endemismo muy elevados (Gentry, 1993, 1995). Al mismo tiempo, son también poco conocidos y muy amenazados en el trópico (Stadmuller, 1987, citado por Andrade 1993). En Colombia se estima que cerca del 90% de la extensión original de bosques andinos ha desaparecido (Henderson et al., 1991), siendo una de las causas principales la concentración de la población en estas regiones, la cual ha dado paso a la deforestación y adecuación de tierras para la agricultura y la ganadería (Andrade, 1993).
En estos bosques la formación de claros en el dosel puede determinar profundos cambios en, las condiciones de luz y modificar la temperatura, humedad y riqueza de nutrientes del. suelo(Denslow, 1980; Brokaw, 1985); estos cambios dependen del tamaño del claro, la edad y su posición en el bosque. Tales diferencias afectan la dinámica de las poblaciones de especies vegetales y conforman un "mosaico sucesional" que contiene simultáneamente diferentes fases de desarrollo del bosque; el conocimiento de su composición y estruCtura posterior a la apertura del dosel constituye una pieza clave para futuros programas de rehabilitación de bosques subandinos\ afectados por la fragmentación, el cambio de uso del suelo y la invasión de especies foráneas.
OBJETIVO GENERAL
Realizar la caracterización fisonómico-estructural del componente vegetal, relacionado con los claros de la parcela permanente de 25 ha en la Reserva Natural la Planada, Nariño, a partir del análisis de la estructura vertical y las variables asociadas al Índice de Valor de Importancia (IVI).
ÁREA DE ESTUDIO
La Reserva Natural La Planada está ubicada en la vertiente pacífica de la Cordillera Occidental, municipio de Ricaurte, departamento de Naririo (1°09'28."N, 77°58'34"W), y cuenta con una extensión aproximada de 3.200 hectáreas de bosque que van desde los 1.300 hasta los 2.100 msmn. No en toda su extensión se pueden encontrar bosques primarios o en buen estado de conservación; algunas zonas han sido transformadas en el pasado y corresponden a lo que se conoce como bosques secundarios o zonas de regeneración natural (Vallejo et al., 2004).
La parcela permanente de 25 ha (Figura 1) está ubicada dentro de la Reserva Natural La Planada en un terreno con buen estado de conservación del bosque, se estableció en un área ligeramente ondulada, con rango de elevación entre 1.796 y 1.891 msnm (Vallejo et al., 2004). Legal y oficialmente, los predios de la planada están cobijados bajo la figura de Reserva Forestal Protectora, reconocida por la junta directiva del desaparecido Instituto Nacional de Recursos Naturales Renovables y del Ambiente (INDERENA), mediante el Acuerdo 19 del 25 de abril de 1984 (FES, 2001).
ASPECTOS GENERALES
La Planada forma parte de la región ampliamente conocida como Chocó Biogeográfico, de acuerdo con la clasificación de ecosistemas de Gentry (1995), también hace parte de los bosques húmedos tropicales (bh-T) que comúnmente se denominan bosques de niebla. Su ubicación intermedia entre las selvas de montaña y los húmedos bosques del Pacífico han propiciado el encuentro de plantas y especies animales de ambos ecosistemas (Vallejo et al., 2004).
ASPECTOS CLIMÁTICOS
La Planada hace parte de un ecosistema conocido como bosque de niebla altoandino, con altos niveles de precipitación a lo largo del año. El clima es bimodal, existiendo dos periodos de alta pluviosidad y dos relativamente secos; los meses más lluviosos van de octubre a enero (media: 515 mm). La pluviosidad disminuye entre febrero y mayo (media: 432,5 mm); la época de verano por lo general se ubica entre julio y agosto (media: 162,5 mm), mientras que los meses de tendencia seca son junio y septiembre (media: 280 mm) FES (2001).
Los datos de temperatura diaria multianual registrados para La Planada indican una temperatura media de 19.2 °C, con una temperatura máxima de 24.5° C y una mínima de 13.1 La humedad relativa o porcentaje de humedad está entre 85-95%. Se reportan elevados niveles de humedad que crean un ambiente propicio para el establecimiento de plantas epifitas (FES 2001). La reserva está ubicada dentro de la zona de vida bosque pluvial premontano (bpPM), según la clasificación por zonas de vida de Holdridge), o bosque subandino siguiendo a Van der Hammen y Rangel (1997), citados por Mendoza y Ramírez (2001).
ASPECTOS GEOLÓGICOS Y EDÁFICOS
La Planada hace parte de una región ubicada en el antiguo geosinclinal Bolívar, el cual se extiende a manera de depresión alargada en la mima dirección de la cordillera occidental, el origen de la región es marino; en la época en que estuvo sumergida, la lava liberada por volcanes dio origen a formaciones de rocas diabásicas. En la región abunda un tipo de roca llamado fluvio-volcánica del periodo terciario-cuaternario (FES, 2001). En la Reserva los suelos predominantes son andisoles, cuyo material parental es piroclástico, derivado de materia orgánica y cenizas volcánicas. La textura es principalmente arenosa, lo cual afecta la aireación del suelo, el drenaje, la retención de la humedad, fertilidad, susceptibles la erosión y la permeabilidad (Vallejo et al., 2004).
MATERIALES Y MÉTODOS
ANTECEDENTES
En 1990, el Centro para las Ciencias Forestales del Trópico (CTFS) del Instituto Smithsonian inició la promoción del establecimiento de parcelas permanentes de gran tamaño para evaluar la composición, estructura y dinámica de los bos4ues tropicales a largo plazo. Actualmente se han establecido 18 parcelas permanentes en el mundo, distribuidas en América Latina (5), África (2) y Asia (10) (Vallejo et al., 2000). El Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH) comenzó en 1996 el montaje de una parcela permanente de 25 ha en un bosque nublado de la Reserva Natural La Planada, ubicada el suroccidente colombiano en límites cercanos al Ecuador (Figura 1).
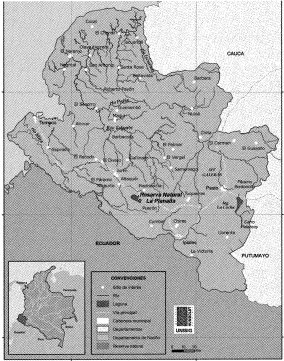
En el primer inventario de 1997 se registraron 115.130 individuos dentro de la parcela, pertenecientes a 63 familias, 141 géneros y 208 morfoespecies. Entre las más abundantes se destacan la familia Rubiaceae con 392.778 in-. dividuos, Arecaceae con 9.619 y Melastomateceae con 9.557 individuos. La especie más abundante corresponde a Faramea calyptrata, y los géneros más abundantes son Miconia con 11 especies, Cyathea con 8 y Psychotria con 8. El 88% de los individuos censados corresponden a tallos entre 1 y 10 cm de DAP y,solo un 2% agrupa especies con individuos de Más de 30 cm de DAP, entre ellos se puede mencionar: el Corozo (Couepia platycalyx), el Corocillo (Solanea sp) y el Hojiancho (Akhornea triplinervia), con registros de más de 100 cm de DAP (IAvH, 2000).
ESTABLECIMIENTO DE LA PARCELA
De acuerdo con Vallejo (2000), para el establecimiento de la parcela (500 x 500 m) y el inicio del primer censo de árboles se contó con el apoyo y asesoría del CTFS, y se siguió la misma metodología empleada para el montaje de la parcela de 50 ha en la Isla de Barro Colorado (Panamá), la cual fue detallada por Condit (1998). Se escogió un bosque maduro, ubicado en un terreno ligeramente ondulado, cuyo dosel alcanza alturas entre los 15 y 25 m. En su interior se diferencian varios tipos de hábitat generados en parte por los cambios microclimáticos ocasionados por la presencia de claros en el bosque.
La parcela fue dividida en 625 cuadrantes de 20x20 m demarcados con tubos plásticos de PVC de 1 m de altura. Cada cuadrante fue a su vez subdividido en 16 subcuadrantes de 5x5 m también marcados con tubos plásticos pero de 50 cm de altura; para la ubicación de los tubos que delimitan cuadrantes y subcuadrantes se hizo ún levantamiento topográfico del terreno con curvas de nivel cada metro de altura.
CENSO DE ÁRBOLES
Durante el ario siguiente (1997) todas las plantas leñosas (menos las lianas) (DAP mayor o igual a 1 cm fueron marcadas con placas de aluminio numeradas con 6 dígitos; cada cuadrante de 20x20 m se utilizó como mapa de referencia para localizar los árboles de la parcela.
A su vez, los subcuadrantes de 5x5 m sirvieron para hacer una aproximación más exacta de los individuos censados y como área de referencia para medir los claros.
Una vez terminado el censo se inició la fase de identificación taxonómica de especies. Los registros obtenidos del censo fueron procesados posteriormente en una base de datos diseñada para el proyecto y los mapas se digitalizaron mediante un Sistema de Información Geográfica (SIG) (Vallejo, 2000).
FUENTE DE LOS DATOS
Los datos que se utilizaron para conducir los análisis del presente estudio fueron facilitados por el Proyecto Dinámica del Bosque Andino del IAvH a través de su base de datos ANDIGENA, que contiene la información alfanumérica de todos los árboles leñosos marcados durante los censos. Los metadatos son los siguientes.
- Número de placa.
- DAP (Diámetro a la altura del pecho)
- Altura de medición del DAP
- Fecha de medición
- Identificación taxonómica de los individuos (familia, género y especie)
- Códigos de medición (múltiple o no)
- Códigos de forma (quebrado, inclinado, rebrote, sin hojas, raíces altas)
- Localización: Coordenadas cartesianos a nivel de cuadrantes de 20m x 20m y de subcuadrantes de 5m x 5m)
- Topografía: pendiente, elevación, convexidad
DEFINICIÓN DEL TAMAÑO DE CLAROS
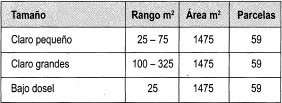
Para efectos del presente estudio, se interpretó claro como una superficie de 5m x 5m o conjunto de éstas, existentes dentro de la parcela permanente, en las cuales se presenta una abertura del dosel que permite la entrada de luz. Los claros se dividieron en dos tamaños en un área total en m2 normalizada: los claros pequeños tienen una extensión de 25 a 75 y los claros grandes de 75 a 325 m2 . Para esta clasificación se tomó como referencia la compilación realizada por Louman et al. (2001), además de la resolución misma de la grilla, la cual no permitía el análisis de áreas menores de 5x5 m.
POBLACIÓN Y MUESTRA
La población de estudio se definió por plantas leñosas (menos lianas) (DAP mayor o igual a 1 cm, presentes en las muestras de vegetación correspondientes a claros y áreas bajo dosel en la parcela permanente de 25 ha localizada en la Reserva. La muestra consistió en individuos presentes en los dos tamaños de claros formados en el ario 2003 y una muestra de vegetación presente en dosel cerrado, que se obtuvo de forma aleatoria en subparcelas bajo dosel, en un área total igual a la de claros (Tabla 1).
SISTEMATIZACIÓN Y ANÁLISIS DE LOS DATOS
Se filtraron de la base de datos ANDIGENA los datos correspondientes a los individuos inventariados en las parcelas que presentaron claros en el 2003; la caracterización fisonómico- estructural y composición florística se hizo con base en los datos del segundo censo, excluyendo los individuos muertos y discriminando los datos por familias, géneros y especies. El análisis de la vegetación se define por medio de atributos cuantitativos como abundancia, frecuencia y dominancia a través del Índice de Valor de Importancia.
ESTRUCTURA VERTICAL
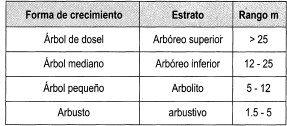
El estudio de la estructura vertical se determinó por el método de clasificación de estratos idealizados, es decir, las especies fueron clasificadas según la posición que ocuparía en el dosel en la madurez, con el propósito de determinar los cambios que ocurren en grupos de plantas dentro de la parcela.
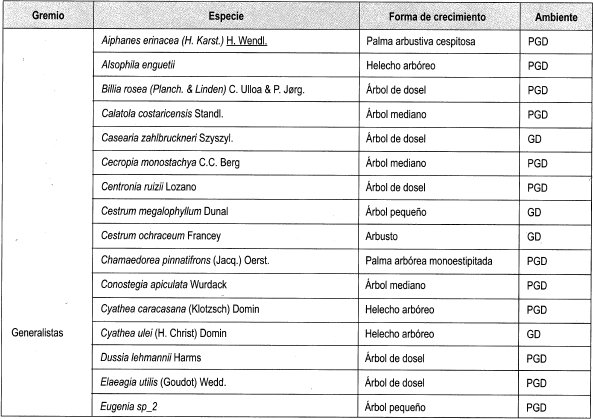
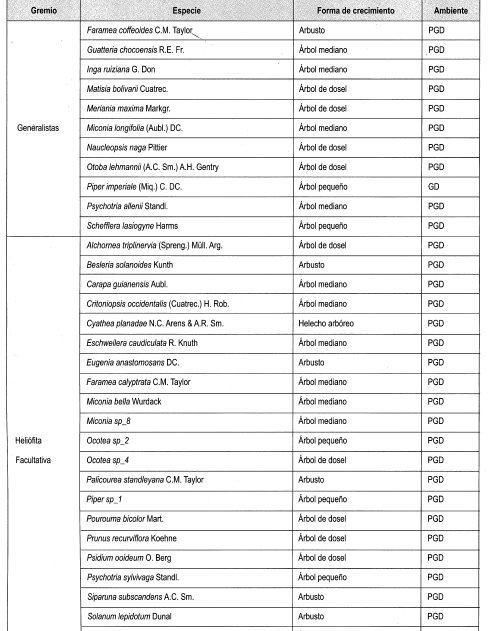
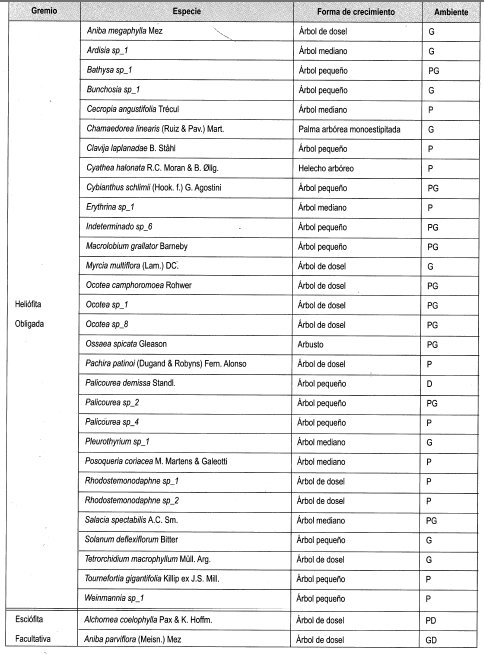
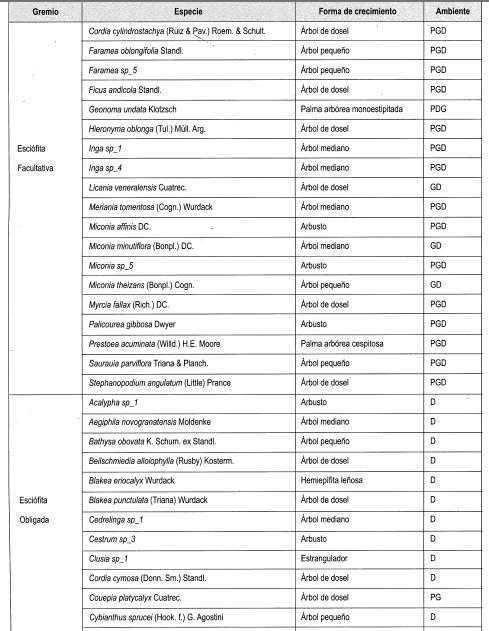
Se interpretó la clasificación de las especies presentes en claros y en dosel cerrado establecida por Mendoza y Ramírez (2000), según su forma de crecimiento (grupos), y .en estratos, de acuerdo con lo propuesto por Rangel y Velásquez (1997), como se muestra en la Tabla 2.
ESTRUCTURA HORIZONTAL
Se determinó en cada muestra la estructura horizontal a partir de las variables de abundancia, frecuencia, área basal, e IVI, para familias, géneros y especies de claros pequeños (25-75 m2), claros grandes (75-325 m 2) y bajo dosel.
CARACTERIZACIÓN FLORÍSTICA
En 118 parcelas que presentaron claros (59 de claros grandes más 59 de claros pequeños) y en 118 parcelas ubicadas bajo dosel se calculó la frecuencia de cada especie. De acuerdo con la metodología propuesta por Damascos y Rapaport (2002), se identificaron en primer lugar las especies exclusivas de los claros (heliófitas obligadas) y las de áreas bajo el dosel (esciófitas obligadas). Para las especies presentes en claros y bajo dosel, se determinó su preferencia por una de estas condiciones comparando su frecuencia mediante la prueba t Student para muestras emparejadas. Fueron clasificadas como heliófitas facultativas, aquellas especies con mayor frecuencia en los claros (P<0.05); esciófitas facultativas, aquellas que presentaron mayor frecuencia bajo el dosel (P<0.05), y generalistas, aquellas especies cuya frecuencia no difirió estadísticamente entre áreas de claros y áreas bajo el dosel.
De acuerdo con la metodología adoptada, las especies presentes en menos de tres parcelas (raras) fueron excluidas de este análisis; adicionalmente, las especies presentes en claros se clasificaron según su frecuencia en claros grandes o pequeños. La identificación de especies y su posterior clasificación taxonómica estuvo a cago de profesionales especializados del IAvH, donde reposan además las respectivas colecciones botánicas de soporte del presente trabajo.
RESULTADOS
CARACTERIZACIÓN FISONÓMICO ESTRUCTURAL
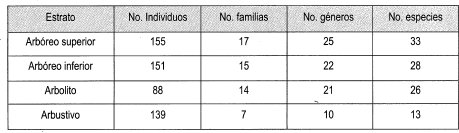
El estudio de la composición fisonómico estructural para la vegetación presente en los claros y bajo dosel, para las especies leñosas con DAP > 1 cm, encontró 1617 individuos, representados en 39 familias, 85 géneros y 161 especies, en un área muestreada total de 4425 m2, correspondiente a la muestra (Tabla 1). Los estratos se idealizaron para las diferentes especies por la falta de datos de altura total y altura a la primera rama, con base en la clasificación de las especies de la Reserva, realizada por el Biólogo Humberto Mendoza del IAvH.
· Distribución de estratos en claros pequeños
Se analizaron los datos de 40 claros pequeños entre 25 y 75 m2, donde hay 525 individuos pertenecientes a 37 familias, 63 géneros y 93 especies.
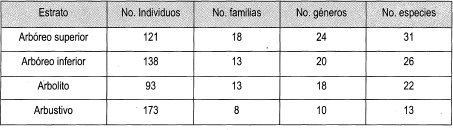
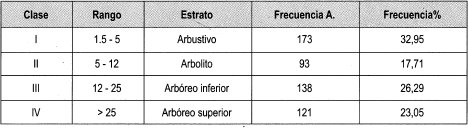
Los estratos con mayor representatividad de individuos son el arbustivo y el arbóreo inferior; el estrato con mayor heterogeneidad de familias, géneros y especies es el arbóreo superior, como se puede apreciar en la Tabla 3. La Tabla 4 muestra la frecuencia relativa de los estratos presentes en las parcelas de claro pequeño, donde el estrato arbustivo tiene la frecuencia relativa más alta por su alta cantidad de individuos, mientras que el estrato arbolito tiene la frecuencia relativa más baja por la poca representatividad de individuos.
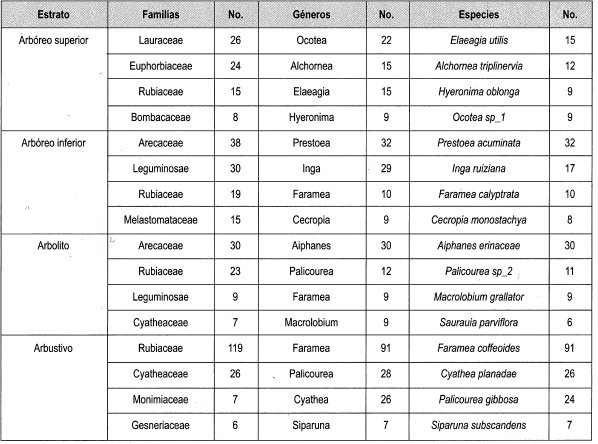
En la Tabla 5 se observan las familias, géneros y especies más importantes por estrato en las parcelas de claro pequeño, donde la familia Rubiaceae está presente en todos los estratos y es dominante en el estrato arbustivo con la especie con mayor cantidad de individuos Faramea coffeoides (616 ind/ha). La familiá Arecaceae domina en los estratos de arbolito y arbóreo inferior con los géneros Prestoea y Aiphanes. Las palmas que dominan estos estratos son Prestoea acuminata (81 ind/ha) y Aiphanes erinaceae (203 ind/ha); en el estrato superior, la familia Lauraceae está representada con el género Ocotea y la especie más representativa es Elaeagia utilis con 101 individuos por ha.
· Distribución de estratos en claros grandes
Se examinaron 9 claros grandes entre 100 m2 y 325 m2, que ocupan un área total de 1475 m2,los cuales incluyen 533 individuos, distribuidos en 34 familias, 63 géneros y 100 especies.
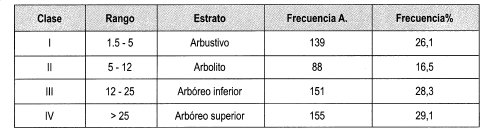
En la Tabla 6 se aprecia la distribución de los estratos en las parcelas de claro grande, donde el estrato con mayor número de individuos, familias, géneros y especies es el arbóreo superior, teniendo alta heterogeneidad y abundancia por sus 155 individuos distribuidos en 33 especies. El estrato arbustivo es más homogéneo, ya que sus 139 individuos están representados en 13 especies. En la Tabla 7 se muestra la frecuencia relativa de los estratos de las parcelas de claro grande, donde los estratos arbóreo superior y arbóreo inferior son los más representativos (57.4% de frecuencia relativa.
Al analizar los estratos por número de familias, géneros y especies (Tabla 8) de las parcelas de claros grandes, se halló que la familia Rubiaceae es la única que se encuentra en los 4 estratos, siendo la más abundante en los estratos arbolito y arbustivo con los géneros Aiphanes y Farame' a. Las especies más importantes son Aiphanes erinaceae y Faramea coffeoides. La familia Arecaceae está presente en los estratos arbóreo inferior y en arbolito, pero solo domina en el primero con el género Prestoea y la especie Prestoea acuminata, la familia Cyatheaceae se presenta en los estratos arbolito y arbustivo, pero no es la dominante en ninguno de los dos. La familia Euphorbiaceae es la que tiene la frecuencia más abundante en el estrato arbóreo superior con el género Ocotea y la especie Elaeagia utilis.
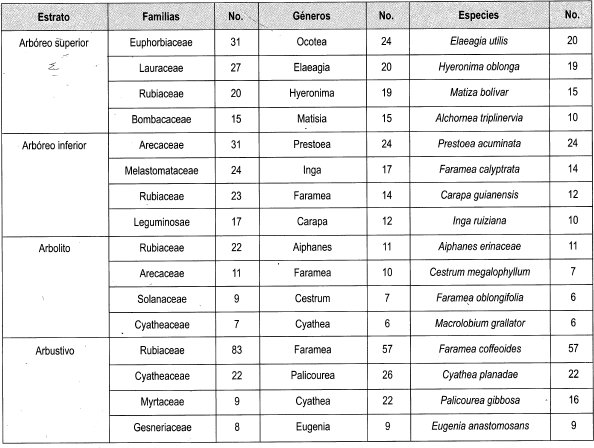
· Distribución de estratos de bajo dosel En la muestra bajo dosel se registraron 617 individuos agrupados en 34 familias, 67 géneros y 110 especies. El estrato más homogéneo es el arbustivo, que solo registra 18 especies y 222 individuos; el estrato más heterogéneo es el arbóreo inferior con 26 especies y 128 individuos (Tabla 9).
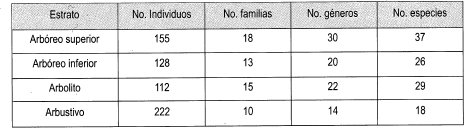
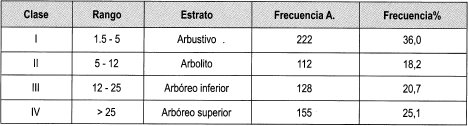
Al observar la Tabla 10 se halló que, en las parcelas bajo dosel, el estrato con mayor frecuencia relativa es el arbustivo (36%), que se concentra más en una parcela. El estrato arbolito es el menos representativo en esta parcela, (18%) pero este registro es el más alto entre las parcelas estudiadas del estrato arbolito.
En la Tabla 11 se observan las más importantes familias, géneros y especies de las parcelas bajo dosel. La familia que se encuentra en todos los estratos es Rubiaceae y es la más representativa en tres estratos con los géneros Elaeagia y Faramea, con las especies Elaeagia utilis, Aiphanes erinaceae y Faramea coffeoides.
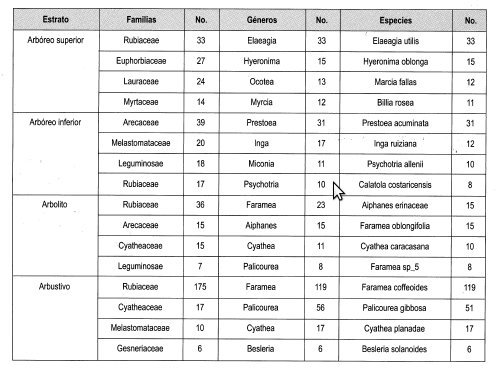
La familia Arecaceae está presente en dos estratos, siendo la más representativa en el estrato arbóreo inferior con el género Prestoea y la especie Prestoea acuminata. Las familias Melastomataceae y Cyatheaceae están representadas en dos estratos, pero no son las representativas en los diferentes estratos.
· Índices de Valor de Importancia (IVI)
En los Anexos 1, 2, 3, 4, 5, 6, 7 y 8 se encuentran los primeros 10 datos de frecuencia relativa, abundancia relativa y dominancia relativa con los cuales se calculó el IVI absoluto y relativo para las familias, género y especies de los claros pequeños, claros grandes y bajo dosel.
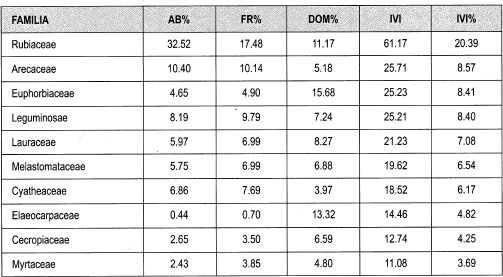
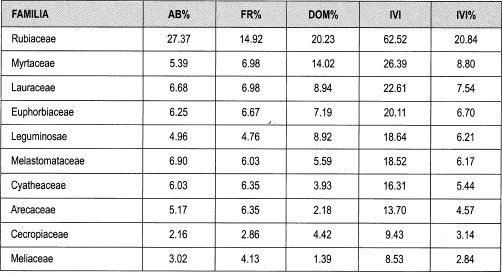
Claros pequeños. Las familias con mayor abundancia relativa son Rubiaceae (33%), Arecaceae (10%), y Leguminosae (8%). La frecuencia relativa está representada por las familias Rubiaceae (17%), Arecaceae y Leguminosae (cada una 10%). Las familias con mayor dominancia por área basal son Euphorbiaceae (16%), tlaeocarpaceae (13%) y Rubiaceae (11%).
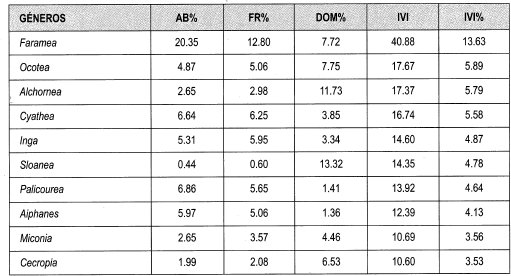
El IVI relativo para las familias del claro pequeño fluctúa entre 20.4% y 0.2%. Las familias con mayor IVI son Rubiaceae (20%), Arecaceae (9%), Euphorbiaceae y Leguminosae (cada una 8%). Los géneros con mayor abundancia relativa son Faramea (20%), Palicourea y Cyathea (7% cada una) y Aiphanes (6%). Los géneros que presentan mayor Frecuencia relativa son Faramea (13%), Cyathea, Inga y Palicourea (6% cada una). Los géneros que muestran mayor dominancia relativa por área basal son Solanea (13%), Akhornea (12%), Ocotea y Faramea (8%).
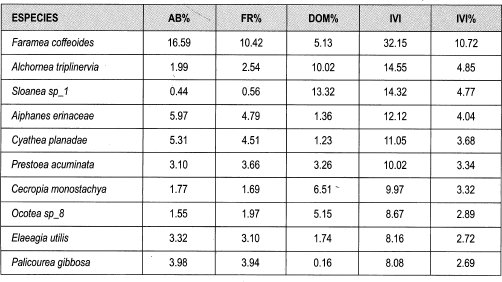
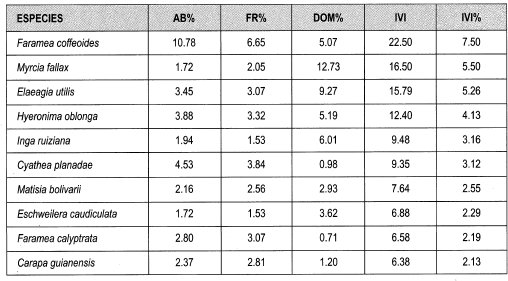
Los datos obtenidos de IVI para género fluctúan entre 14 y 0.2%. Los géneros con mayores porcentajes de IVI son Faramea (14%), Ocotea, Alchornea y Cyathea (6% cada una); las especies que presentan mayor abundancia relativa son Faramea coffeoides (17%), Aiphanes erinaceae (6%), Cyathea planadae (5%) y Palicourea gibbosa (4%). Las especies que muestran más frecuencia relativa son Faramea coffeoides (10%), Aiphanes erinaceae y Cyathea planadae (cada una 5%), Palicourea gibosa y Prestoea acuminata (4%). Las especies que tienen mayor dominancia por área basal son: Sloanea sp_l (13%), Alchornea triplinervia (10%), Cecropia monostachya (7%), Ocotea sp_8 y Faramea coffeoides (5%). Las especies que presentan mayor IVI son Faramea coffeoides (11%), Alchornea triplinervia y Sloanea sp_l (cada una 5%), Aiphanes erinaceae y Cyathea planadae (4% cada una).
Claros grandes. Las familias que presentan mayor abundancia son Rubiaceae (27%), Melastomataceae y Lauraceae (cada un 7%), Euphorbiaceae y Cyatheaceae (6%). Las familias que muestran mayor frecuencia son Rubiaceae (15%), Lauraceae, Myrtaceae y Euphorbiaceae (7% cada una), Cyatheaceae, Arecaceae y Melastomataceae (cada una 6%). Las familias de mayor dominancia por área basal son Rubiaceae (20%), Myrtaceae (14%), Lauraceae y Leguminosae (9%).
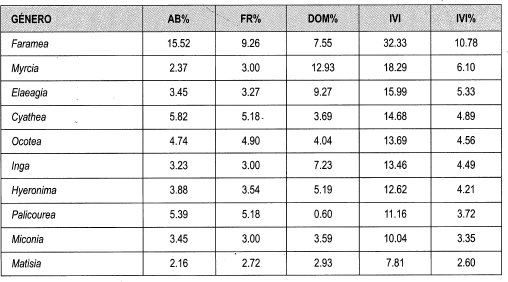
Los datos obtenidos del IVI de las familias de claros grandes fluctúan entre 20.8% y 0.2%. Las familias más representativas por su mayor porcentaje de IVI son Rubiaceae (21%), Myrtaceae (9%), Lauraceae (8%) y Euphorbiaceae (7%). Los géneros que presentan mayor abundancia son Faramea con el 16%, Cyathea (6%), Palicourea (5%), Ocotea (5%) y Hyeronima (4%). Los géneros más frecuentes son F aramea (9%), Cyathea, Palicourea y Ocotea (5%) y Hyeronima (4%). Los géneros que presentan mayor dominancia por área basal son Myrcia (13%), Faramea (8%) e Inga (7%).
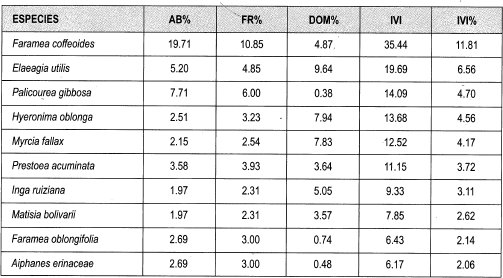
Entre los datos obtenidos de IVI relativo para los géneros bajo dosel sobresalen Faramea (11%), Myrcia (6%), Elaeagia,',Cyathea y Ocotea (cada una 5%), Inga, Hyeroniina y Palicourea (4%). Las especies que presentan mayor Abundancia son Faramea coffeoides (11%), Cyathea planadae (5%), Hyeronima oblonga (4%), Elaeagia utilis, Faramea calyptrata y Palicourea gibbosa(3%). Las especies con mayor frecuencia son Faramea coffeoides (7%), Cyathea planadae (4%), Hyeronima oblonga, Elaeagia utilis, Faramea calyptrata, Carapa guianensis, Palicourea gibosa, y Matisia bolivarii (cada una 3%). Las especies que presentan mayor dominancia por área basal son Myrcia fallax (13%), Elaeagia utilis (9%), Inga ruiziana (6%), Hyeronima oblonga y F aramea coffeoides (5%).
Con respecto al IVI relativo para las especies presentes en claros grandes se destacan Faramea coffeoides (7%), Myrcia fallax, Elaeagia utilis (5%), Hyemnima oblonga (4%), Inga ruiziana, Cyathea planadae y Matisia bolivarii (3%).
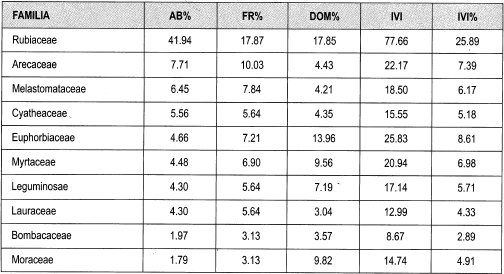
Bajo dosel. Las familias que presentan mayor abundancia son Rubiaceae (42%), Arecaceae (8%), Melastomataceae, Cyatheaceae (6%) y Euphorbiaceae (5%), mientras que las más frecuentes son Rubiaceae (18%), Arecaceae (10%), Melastomataceae (8%), Euphorbiaceae y Myrtaceae, (7%). La mayor dominancia está representada por Rubiaceae (18%), Euphorbiaceae (14%), Moraceae, Myrtaceae (10%) y Leguminosae.
Las familias que presentan mayor porcentaje de IVI son Rubiaceae (26%), Euphorbiaceae (9%), Arecaceae, Myrtaceae (7%), Melastomataceae y Leguminoseae (6%). En cuanto a los géneros, los presentan mayor IVI son Faramea (15%), Elaeagia (7%), Palicourea (6%) y Miconia (5%). Las especies presentes en parcelas de bajo dosel con mayor peso ecológico son Faramea coffeoides (11.8%), Myrcia fallax, Elaeagia utilis (5%), Hyeronima oblonga (4%), Inga ruiziana, Cyathea planadae y Matisia bolivarii (3%).
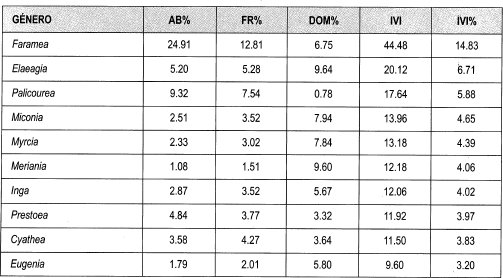
Los géneros más abundantes son Faramea (25%), Palicourea (9%), Elaeagia, Cyathea (5%), Prestoea (4%); en cuanto a la frecuencia se destacan Faramea (13%), Palicourea (8%), Elaeagia (5%), Cyathea, Prestoea, Inga y Miconia (4%); mientras que por su dominancia sobresalen Elaeagia, Meriania (10%), Miconia, Myrcia (8%), Faramea (7%), Eugenia e Inga (6%).
Las especies que presentan mayor abundancia son Faramea coffeoides (11%), Cyathea planadae (5%), Hyeronima oblonga (4%), Elaeagia utilis, Faramea calyptrata y Palicourea gibosa (3%); dentro de las más frecuentes se pueden mencionar Faramea coffeoides (7%), Cyathea planadae (4%), Hyeronima oblonga, Elaeagia utilis, Faramea calyptrata, Carapa guianensis, Palicourea gibosa, Matisia bolivarii (3%), Akhornea triplinervia y Eugenia anastomosans (2%); finalmente por su área basal Myrcia fallax (13%), Elaeagia utilis (9%), Inga ruiziana (6%), Hyeronima oblonga y F aramea coffeoides (5%) constituyen el grupo más representativo.
· Caracterización florística
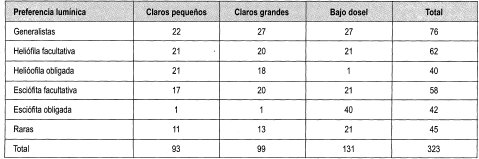
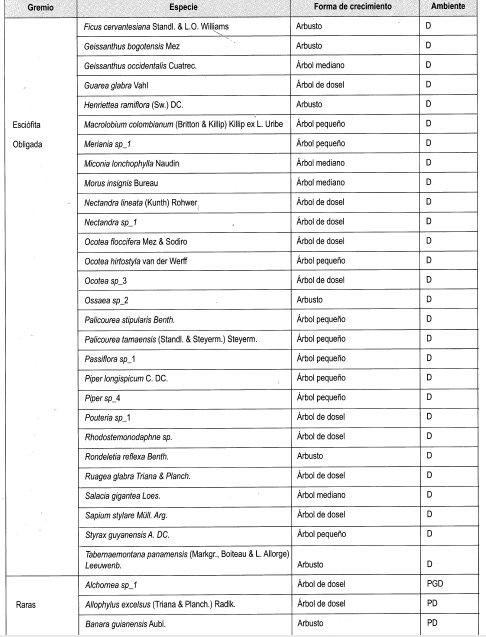
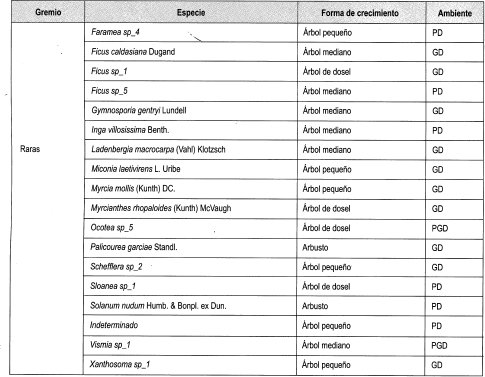
En el Anexo 10 se relacionan las especies por su forma de crecimiento y su preferencia lumínica. Se identificaron en total 161 especies, de las cuales se clasificaron las especies presentes en más de dos parcelas, siendo las especies esciófitas obligadas y facultativas las más numerosas (39%), seguidas de las especies heliófitas obligadas y facultativas (31%). Las especies generalistas representan el 17% del total; 21 especies se consideraron raras por estar presentes en menos de tres parcelas (Damascos y Rapaport, 2002).
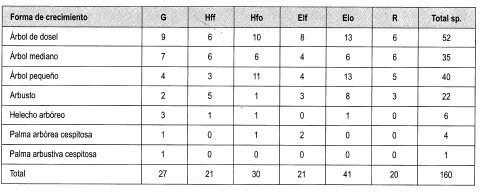
Con base en la Tabla 12 y la Figura 2, es posible inferir que las formas de crecimiento predominantes corresponden a árboles, entre los cuales sobresalen los esciófitos obligados, seguidos de los heliófitos obligados y los genera listas. Sin embargo se débe resaltar que, en las formas de crecimiento superiores, la evidencia muestra que las preferencias lumínicas guardan una proporción relativa, aun en las consideradas raras.
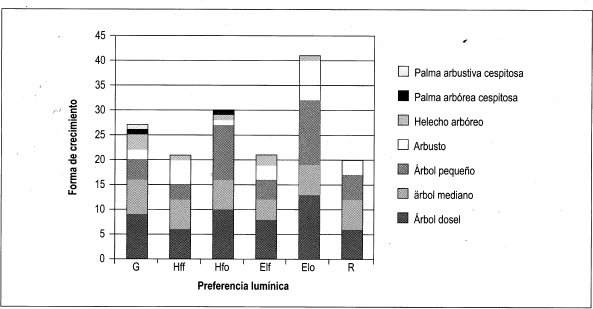
En general se puede afirmar, con base en la vegetación asociada a claros, que no existe una marcada diferenciación con respecto a preferencias sus lumínicas, es decir que independiente de las formas de crecimiento dentro del bosque, la dinámica asociada a la formación y recuperación de claros lleva a una redistribución constante de los temperamentóscle las especies, evitando el predominio competitivo y ampliando el espectro de posibilidades de la diversidad.
En la Tabla 13, como balance, se hace de nuevo énfasis en el carácter generalista de gran can- tidad de especies, así como la equiparidad relativa asociada a la preferencia lumínica en los tres diferentes ámbitos evaluados. Se reafirma lo anterior al evidenciarse un comportamiento similar a las generalistas de las facultativas, tanto heliófitas como esciófitas. Por otro lado, la cantidad de especies no experimenta grandes variaciones relacionadas con el tamaño de claro, lo cual podría estar asociado no solo al comportamiento antes reseñado, sino al tamaño de la perturbación.
CONCLUSIONES
Como se ha afirmado (Asquith, 2002), la investigación en claros ha contemplado muchos aspectos relacionados con la definición de sus límites espaciales, la frecuencia, el origen, el tamaño de la apertura del dosel, la dependencia de distintas especies de plantas a la presencia de un claro, su papel en la depredación de semillas, su efecto sobre los nutrientes del suelo y el crecimiento de las raíces y su injerencia en la tasa de mortalidad de los árboles remanentes. Aun así, salvo los casos ampliamente conocidos de La Selva y Barro Colorado, los trabajos basados en parcelas permanentes de monitoreo aún son pocos para nuestros bosques neotropicales; de ahí el significado de unos resultados que representan el esfuerzo institucional por contar con información validad con el tiempo.
Clark y Clark (1992) sugieren que los requerimientos de luz de las distintas especies de árboles varían durante su ciclo de vida de acuerdo con su estado de desarrollo, y que estos requerimientos pueden diferir entre una especie y otra. Los autores proponen incorporar las etapas del ciclo de vida de los árboles para lograr una clasificación más realista y objetiva de la estrategia ecológica de estos, y reemplazar así otras clasificaciones más rígidas, como la que distingue entre especies "pioneras" y especies "tolerantes a la sombra", las cuales por lo general remiten a un solo estadio de tamaño.
Se ha sostenido también, que salvo algunas especies arbóreas consideradas estrictamente "pioneras" o "demandantes de luz", la gran mayoría son en realidad "generalistas" (Lieberman et al. 1995); es decir, no presentan un alto grado de especialización en cuanto a sus necesidades de luz. La hipótesis de la ausencia de tal dicotomía se soporta en que en el bosque húmedo neotropical, la luminosidad varía drásticamente entre el nivel más alto del dosel y el piso del bosque. A esta variabilidad se suma la determinada por la dinámica de perturbaciones naturales del bosque, que tipifican como factor ecológico la distribución de las especies que viven en distintos ámbitos del bosque y como factor evolutivo las adaptaciones que ellas desarrollen (Terborgh, 1985, citado por Hogan y Machado, 2002).
Los resultados que arroja la investigación apuntan en esa misma dirección al no encontrar grandes diferencias entre variables medidas en los tres ámbitos seleccionados, y resaltar el carácter generalista de una gran proporción de las especies, géneros y familias. De ningún modo esto sugiere la homogeneidad de este tipo de bosques, sino la importancia de la dinámica de perturbaciones en la estabilidad del sistema y en su funcionamiento. Se debe tener en cuenta que estos bosques están sujetos en su mayoría a perturbaciones naturales de pequeña escala, lo que haría en parte justificable la ausencia de valores extremos en las variables evaluadas; sin embargo, hay que destacar la gran amplitud en la capacidad de asimilar las perturbaciones que poseen estos bosques, pues como se mostró, la magnitud de la perturbación puede ampliarse hasta 13 veces sin castigar la estructura significativamente.
Consideraciones importantes derivadas del estudio sugieren que las preferencias lumínicas deben ser contrastadas de acuerdo con el estado de desarrollo, con la forma de crecimiento o incluso con los niveles de intervención del bosque para el diseño de propuestas de restauración o manejo. De igual forma se reiteran ideas de trabajos anteriores, los cuales sugieren considerar la dinámica de perturbaciones, ya sea natural o antrópica, como herramienta para el diseño de propuestas de manejo más acordes con las condiciones de extensas selvas que nunca van a estar bajo un régimen de protección estricta, y deberán ser usadas para el cumplimiento de funciones sociales de comunidades que viven de ellas.
AGRADECIMIENTOS
Los autores agradecen al Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH) y a la doctora Martha Vallejo por permitir el uso de la base de datos ANDIGENA del Proyecto Estructura, Dinámica y Regeneración del Bosque Andino, así como al Centro de Información y Documentación y al Banco de Imágenes Ambientales del IAv1-1, igualmente al profesor Orlando Riario por su interés y asesoría, y fundamentalmente a la Universidad Distrital Francisco José de Caldas por su apoyo logístico y de capital humano, invaluables para el desarrollo del presente trabajo.
REFERENCIAS BIBLIOGRÁFICAS
- Andrade, 1.G. 1993. Paisaje y biodiversidad en las selvas de los Andes. Pp. 31-47. En: Germán Andrade (ed.). En Carpanta, selva nublada y páramo. Ecología y conservación de un sistema alto andino.
- Asquith, N. 2002. La dinámica del bosque y la diversidad arbórea. Pp. 377-406. En:
- Guariguata M. y Kattan G. (compiladores).Ecología y conservación de bosques neotropicales. Ediciones Lur. Costa Rica, 691 p.
- Brokaw, N. 1985. Treefalls, regrowth, and comunita structure in tropical forest. En: The ecology of natural disturbance and patch dynamics. S.T. Pickett and P.S. White. Academic Press. Orlando, Florida.Condit, R. 1998. Tropical Forest Census Plots: methods and results from BCI, Panamá and comparison with other plots. Springer-Verlang, 211 p.
- Damascos A. y Rapaport, E. 2002. Diferencias de la flora herbácea y arbustiva entre claros y áreas bajo dosel en un bosque de Notofagus pumilio en Argentina. Departamento de Ecología. Universidad Nacional del Comahue, 8400 Bariloche, Argentina.
- Denslow, J. 1980. Grap partitioning among tropical rainforest trees. Biotropica Supl.to 12(2): 47-55
- Duellman, W.E. 1982. Comprensión climática cuaternaria de los Andes. Efecto sobre la especiación. Pp 77-201. En Salinas, P.J.(De) Zoología Neotropical. Acta VII Congreso Latinoamericano de Zoología. Mérida Venezuela.
- FES. 2001. Plan de Manejo Reserva Natural La Planada 2001-2010. Fundación FES, Reserva Natural la Planada. Bogotá, Colombia,
- Gentry, A.H. 1993. Vistazo general a los bosques nublados andinos y a la flora de Carpanta. Pp. 67-79. En: Germán Andrade (ed). En Carpanta, selva nublada y páramo. Ecología y conservación de un sistema alto andino.
- Gentry, A.H. 1995. Pattems of diversity and floristic composition in neotropical montane forest. Biodiversity. Biodiversity and Conservation of Neotropical Montane Forest. Pp. 103-126.
- Henderson, A., S.P. Churchill & J. Luyeyn. 1991. Neotropical plant diversity. Nature 351: 21-25.
- Hogan K. y J. L. Machado, 2002. La luz solar: consecuencias biológicas y medición.En: Guariguata M. y Kattan G. (compiladores). Ecología y conservación de bosques neotropicales. Ediciones Lur. Costa Rica, 691 p.
- Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. 2000. Colombia inegadiversa: cinco arios explorando la riqueza de un país biodiverso. Bogotá: Instituto Humboldt. Pp. 57-61.
- Lamprecht, H. 1990. Silvicultura en los trópicos: los ecosistemas forestales en los bosques tropicales y sus especies arbóreas, posibilidades y métodos para un aprovechamiento sostenido. GTZ. República Federal Alemana.
- Lieberman, M., D. Lieberman, R. Peralta & G. S. Hartshorn. 1995. Canopy closure and the distribution of tropical forest tree species at La Selva, Costa Rica. Joumal of Tropical Ecology 11: 161-178.
- Louman B., Quirós D. y M. Nilsson. 2001 (Editores). Silvicultura de bosques latifoliados húmedos con énfasis en América Central. Serie Técnica, Manual técnico 46.CATIE. Turrialba, Costa Rica, 265 p.
- Mendoza, C. y B. Ramírez. 2000. Flora de La Planada: Guía ilustrada de Familias y Géneros. Instituto Alexander von Humboldt, Fundación FES, WWF, Bogotá.
- Mendoza, H. y Ramírez, B. 2001. Dicotiledóneas de La Planada, Colombia: lista de especies. Biota Colombiana: V. 2, N° 1,septiembre de 2001.
- Rangel O. y Velásquez, A. 1997. Métodos de estudio de la vegetación. En: O. Rangel, Lowy. P. y Aguilar. M. Colombia Diversidad Biótica, Tomo II: Tipos de vegetación en Colombia. Instituto de Ciencias Naturales Universidad Nacional de Colombia.Bogotá.
- Vallejo, M. I. 2000. Estructura, demografía y dinámica del bosque Andino. Instituto de Investigaciones Biológicas Alexander von Humboldt. Documento interno, s.p.
- Vallejo, et al. 2004. Tropical forest diversity and dynamism: Findings from a Large-Scale Plot Metwork. University of Chicago Press.
- Whitmore T. 1989. Canopy gaps and the two major groups of forest trees. Ecology 70: 536-538.
ANEXOS
Creation date:




Licencia
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






