
DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2012.2.a01Publicado:
01-07-2012Número:
Vol. 15 Núm. 2 (2012): Julio-DiciembreSección:
Artículos de investigación científica y tecnológicaESTIMACIÓN DE CARBONO ALMACENADO EN LA REGENERACIÓN NATURAL DE Pinus durangensis Martínez EN EL SALTO, DURANGO
Estimation of carbon stock in the natural regeneration of Pinus durangensis Martínez in El Salto, Durango
Palabras clave:
biomass, CO2, components, growth, models, volume. (en).Palabras clave:
biomasa, CO2, componentes, crecimiento, modelos, volumen. (es).Descargas
Referencias
Aguilar, R.M. (1991). Comparación de cuatro modelos matematicos aplicados al crecimiento forestal. Revista de Ciencia Forestal, 16(70), 87-107.
Clutter, J., Fortson, J., Pienaar, G., Brister, G., & Bailey, R. (1983). Timber Management: a Quantitative Approach. New York: John Wiley and Sons. 333 p.
Cubillos, D.V. (1987). Modelos de crecimiento diametral para algunos renovales de Raullí. Ciencia e Investigación Forestal, 1, 67-77.
García, C.X., Lezama, P.C., & Santiago, R.B. (1992). Modelos de crecimiento para una plantación de caoba (Swietenia macrophylla King). Revista de Ciencia Forestal, 17(71), 88-102.
Gayoso, J., Guerra, J., & Alarcon, D. (2002). Contenido de carbono y funciones de biomasa en especies nativas y exoticas: Medicion de la capacidad de captura de carbono en bosques de Chile y promoción en el mercado mundial. Valdivia: Universidad Austral de Chile. 157 p.
IPCC [Intergovernmental Panel on Climate Change]. (1994). The supplementary report to the IPPCC scientific assessment. Cambridge: Cambridge University Press. 47 p.
Morales, C.F., & Ruiz, G.V.J. (2004). Estimación de peso verde, peso seco y volumen de fuste mediante la densidad aparente de la madera en Pinus cooperi y Pinus leiophylla. En la región de El salto, Durango (Tesis de pregrado). El Salto, Durango: Instituto Tecnologico Forestal El Salto. 92 p.
Van Laar, A., & Akca, A. (1997). Forest Mensurasión. Göttingen: Cuvillier Verlag. 418 p.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
 |
Montes de Oca-Cano, E., Rojas-Ascensión, M., García-Ramírez, P., Nájera-Luna, J.A., Méndez-González, J., & Graciano-Luna, J.J. (2012). Estimación de carbono almacenado en la regeneración natural de Pinus durangensis Martínez en El Salto, Durango. Colombia Forestal, 15(2), 151-159. |
ESTIMACIÓN DE CARBONO ALMACENADO EN LA REGENERACIÓN NATURAL DE Pinus durangensis Martínez EN EL SALTO, DURANGO
Estimation of carbon stock in the natural regeneration of Pinus durangensis Martínez in El Salto, Durango
Eduardo Montes de Oca-Cano1, Mauricio Rojas-Ascensión2, Pedro García-Ramírez3, Juan Abel Nájera-Luna4, Jorge Méndez-González5 & José de Jesús Graciano-Luna6
1Instituto Tecnológico Superior de Zongolica. Carretera a la Compañía Col. Tepetitlanapa. C.P. 95005, Zongolica. Veracruz, México. montesdeoca_cano@hotmail.com. Autor para correspondencia.
2Instituto Tecnológico Superior de Zongolica. Veracruz, México. mauricio_docente@itszongolica.edu.mx
3Instituto Tecnológico de El Salto, Maestría en Ciencias en Desarrollo Forestal Sustentable. Durango, México. garp820203@hotmai.com
4Instituto Tecnológico de El Salto. El Salto, Durango, México. jalnajera@yahoo.com.mx
5Universidad Autónoma Agraria Antonio Narro. Coahuila, México. jorgemendezgonzalez@gmail.com
6Instituto Tecnológico de El Salto. El Salto, Durango, México. forestal@itelsalto.edu.mx
Recepción. 6 de diciembre de 2011 / Aprobación. 28 de junio de 2012
RESUMEN
El presente estudio tuvo como objetivo ajustar ecuaciones lineales y no lineales para estimar carbono, por componente de acículas, ramas, fuste y total. Los resultados indican que en el ámbito individual, el componente total muestra los mejores estadísticos de ajuste, mientras que las mayores variaciones se detectan en las ramas y en el fuste. Adicionalmente se analizan porcentajes de biomasa por componente donde se observa que el fuste aporta el 46.90 % de la biomasa, las acículas el 35.60 % y las ramas solamente el 17.30 % de la biomasa total. Asimismo, dentro de este rango de edad, la biomasa en el fuste aumenta anualmente 2.96 %, la biomasa en las acículas disminuye 3.13 % y la biomasa en las ramas aumenta 0.17 %.
Palabras clave: biomasa, CO2, componentes, crecimiento, modelos, volumen.
ABSTRACT
The aim of the present study was to adjust linear and non-linear equations in order to estimate the carbon component of leaves, branches and entire trees. The results indicate that at the individual level, the total component shows the best adjustment statistics, while major variation is detected in the branches and in the trunk. In addition an analysis of the percentages of biomass in each component indicates that the trunk accounts for 46.9 % of the biomass, leaves 35.6 % and the branches only 17.3 %. Trunk biomass increases annually by 2.96 %, leaf biomass decreases by 3.13 % and branch biomass increases by 0.17 %.
Key words: biomass, CO2, components, growth, models, volume.
INTRODUCCIÓN
Las herramientas clásicas en el manejo forestal son los modelos de crecimiento, como tecnologías adecuadas en la estimación de variables de manejo, los modelos en el ámbito rodal predicen el rendimiento y la productividad por el pronóstico de los atributos del rodal como: área basal, densidad, volumen residual etc., el modelaje de crecimiento y la productividad puede ser constituido desde diferentes alternativas en el ámbito rodal del grupo de árboles o especies y en el ámbito del árbol individual (Clutter et al., 1983) aspectos que se pueden aplicar a la estimación de biomasa y captura de carbono. El volumen representa la parte más importante de una masa forestal debido a que una vez estimado se puede planificar el manejo actual y futuro de los bosques, así como la forma en que deberá desarrollarse dicho manejo. Actualmente es utilizado eficientemente dentro de proyectos de servicios ambientales, como captura de carbono y servicios hidrológicos (Morales & Ruiz, 2004). Para la estimación del crecimiento de los árboles se utilizan modelos matemáticos estadísticos que predicen el crecimiento de los individuos en un periodo determinado y sirve, en consecuencia, como elemento de apoyo para la planificación de intervenciones silvícolas (Cubillos, 1987). La presente investigación estudia los flujos y la asimilación (intercambio gaseoso) o fijación de CO2 atmosférico, del ambiente a la planta, en la regeneración natural de Pinus durangensis Martinez de la región de El Salto, Pueblo Nuevo, Durango.
Esta investigación tuvo como objetivo principal estimar biomasa para determinar carbono en los componentes de la regeneración natural de P. durangensis ajustando ecuaciones que permitan conocer estos importantes flujos como servicio ambiental. La hipótesis indica que la dinámica de captura de carbono es similar en los componentes foliar, ramal y fustal así como en las diferentes edades de la regeneración natural de P. durangensis de la región de El Salto, Durango.
MATERIALES Y MÉTODOS
ÁREA DE ESTUDIO
El estudio fue realizado en la región de El Salto Durango (México), inmerso en el Ejido denominado San Esteban y Anexos, se localiza en el macizo montañoso denominado Sierra Madre Occidental dentro del municipio de Pueblo Nuevo, al suroeste del estado, su lugar de acceso es por la carretera Durango-Mazatlán en el kilometro 101.
TOMA DE MUESTRAS PARA EL ANÁLISIS DESTRUCTIVO
Para la toma de muestras se seleccionaron 72 árboles de P. durangensis dentro del área de estudio, árboles libres de plagas y enfermedades, libres de defectos tanto físicos como mecánicos, árboles que representaran las categorías díamétricas de altura y edad correspondientes a la etapa de regeneración. Se extrajeron un promedio de siete arbolitos por categoría de edad, desde tres hasta doce años. Se etiquetaron todas las muestras con una clave de identificación con el número de árbol, la categoría de edad y fecha. Una vez obtenidos los arbolitos, se pusieron en bolsas de plástico para evitar, por un lado, la pérdida de humedad excesiva y, por otro lado, conservar un orden para el análisis de datos.
A cada individuo se le midieron las siguientes variables dasométricas: edad (E) fue estimada mediante el número de verticilos que presentó cada árbol muestra; diámetro a la base del suelo (DB) y diámetro a 10 cm de la base del suelo (D) medidos mediante vernier; altura del fuste limpio (HF) y altura total (HT) obtenidas mediante cinta métrica; numero de ramas (NR) se obtuvo mediante la cuenta del numero de ramas realizada en forma visual; cobertura (CB) medida mediante cinta métrica.
PROCEDIMIENTO DE LABORATORIO
Estimación de peso seco de los individuos
Cada árbol fue separado por sus componentes acículas, ramas y fuste, posteriormente cada componente fue pesado para obtener el peso fresco. Las muestras se introdujeron en una estufa de secado de tipo convencional perteneciente al Instituto de Producción y Enseñanza Forestal (INAFO), manteniendo la muestra a temperaturas superiores a los 75°C durante ocho días (Van Laar & Akca, 1997) y hasta tener el peso constante de cada componente (según muestras monitoreadas). El peso seco de cada uno de sus componentes de cada árbol, se obtuvo utilizando básculas con capacidades de 0.32, 10 y 25 kg con precisión de 0.001, 10 y 50 g respectivamente.
Estimación del carbono
Conocido el peso seco de cada componente de la regeneración natural de P. durangensis es posible determinar el contenido de carbono. Aunque el contenido de carbono en la especie puede variar desde 40.27 hasta 46.29 e incluso más de 50 % (Gayoso et al., 2002), una ponderación específica e interespecífica de factores de contenido de carbono de raíces, fuste, ramas y aciculas arroja una aproximación cercana al 50 %, sugiriendo que cerca de la mitad del peso anhidro de sus constituyentes sólidos es equivalente a contenido de carbono. Este factor de conversión fue sugerido por el Panel Intergubernamental del Cambio Climático (IPCC, 1994) para estudios de captura de carbono. Esta consideración fue importante y tomada en cuenta para este estudio.
CRITERIOS DE SELECCIÓN DE MODELOS
Los criterios utilizados para determinar el mejor modelo fueron la R2, el error estándar, coeficiente de variación, sesgo, F calculada, así como la probabilidad del modelo y de los parámetros de regresión. Otro criterio tomado en cuenta fue la parsimonia del modelo y numero de variables consideradas (Tabla 1).

RESULTADOS
En la tabla 2 se muestran los parámetros de los modelos probados para estimar el incremento en diámetro, altura, carbono en acículas, carbono en ramas, carbono en fuste y carbono total en la regeneración natural de P. durangensis.
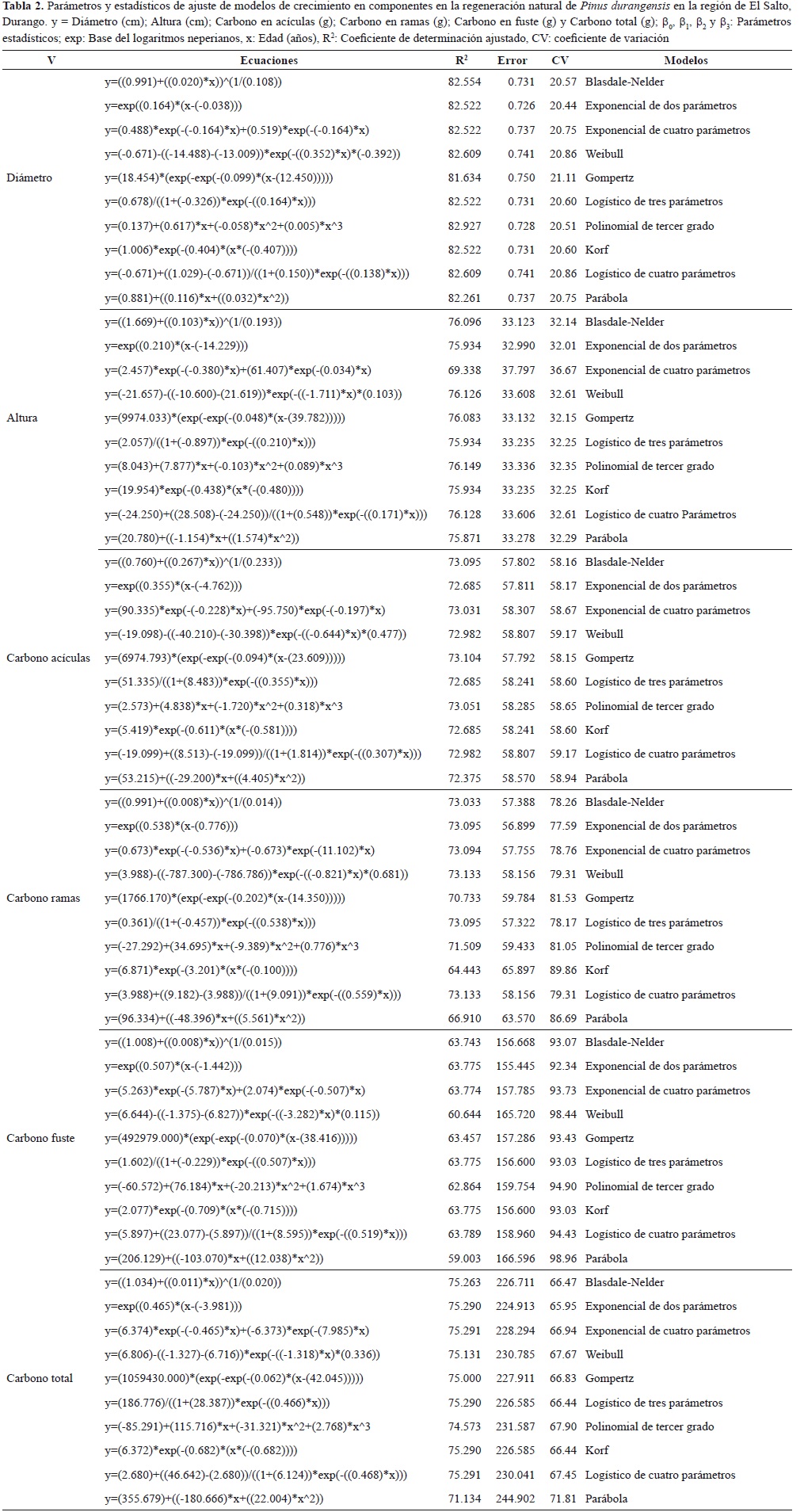
Basados en el promedio de la R2 de los diez modelos que se ajustaron en cada componente de la regeneración natural de P. durangensis se observa que los modelos se ajustan mejor al diámetro seguido por la altura, carbono total, carbono de las acículas, carbono de las ramas y finalmente el componente carbono en el fuste, esto es debido a la gran variación de tamaño y biomasa de fuste en las diferentes categorías de edad, al igual que la gran variación en la biomasa de ramas.
El mejor modelo que se ajustó para estimar el incremento en diámetro de P. durangensis fue el exponencial de dos parámetros presentando menor error (0.72) y mayor R2 (82.52). El modelo que menos ajuste presentó en la estimación de crecimiento en diámetro fue el de Gompertz presentando mayor error (0.74) y menor R2 (81.63).
El modelo que mejor ajuste presentó en el crecimiento en altura en la especie en regeneración natural de P. durangensis fue el polinomial de tercer grado, mostrando una R2 de 76.149 y error de 33.336. Por otra parte, el modelo que menos ajuste presentó para determinar el crecimiento en altura fue el exponencial de cuatro parámetros presentando menor R2 de 69.338 y mayor error de 37.797.
Para la determinación del incremento de carbono en las acículas el modelo que mayor ajuste presentó fue el de Gompertz presentando mayor R2 de 73.104 y menor error estándar de 57.792. El modelo que menor ajuste tuvo en modelar el incremento de carbono en las acículas fue el de la parábola, presentando un menor R2 de 72.375 y uno de los mayores errores (58.570).
El modelo el exponencial de dos parámetros fue el modelo que presentó mejor ajuste para la determinación del incremento de carbono en las ramas en la especie de P. durangensis de regeneración natural, registrando una de las mejores R2 (73.096), menor error (56.899) así como menor coeficiente de variación. El modelo que menos ajuste presentó para estimar el incremento en carbono en ramas fue el de Korf presentando una menor R2 (64.444), mayor error y mayor coeficiente de variación.
El modelo que mostró mejor ajuste en la estimación del incremento de carbono en el fuste en la regeneración natural de P. durangensis fue el exponencial de dos parámetros presentando una de las mejores R2, siendo de 63.775 y de menor error (155.445), así como menor coeficiente de variación, mientras que el modelo que menos ajuste presento para modelar el incremento de carbono en el fuste de la especie estudiada fue el modelo de la Parábola, ya que los estadísticos de ajuste no fueron muy aceptables.
El incremento de carbono total en P. durangensis fue mejor explicado por el modelo Exponencial de dos parámetros por presentar la segunda mayor R2 de 75.290 y un error de 224.913. Contrariamente el modelo que menos ajuste presentó para estimar esta variable fue el de la Parábola presentando el menor R2 (71.134) y un mayor error (244.902).
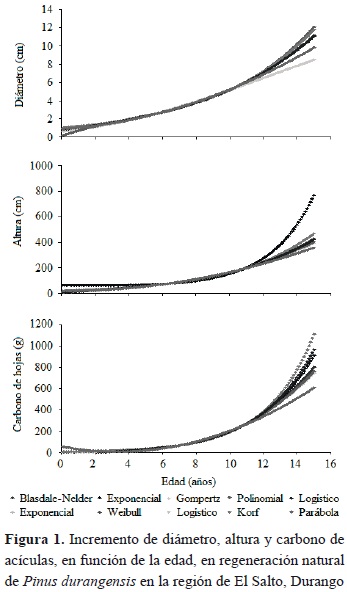
En las figuras 1 y 2 se muestra el comportamiento de los modelos de crecimiento utilizando la edad como variable independiente para la estimación del incremento en diámetro, altura, carbono en acuculas, en ramas, en fuste y carbono total en la regeneración natural de P. durangensis.
Los puntos que se muestran en la figura 3 corresponden a la biomasa promedio (por componente) de siete árboles por categoría de edad, mientras que las líneas indican las tendencias a lo largo de la edad del árbol.
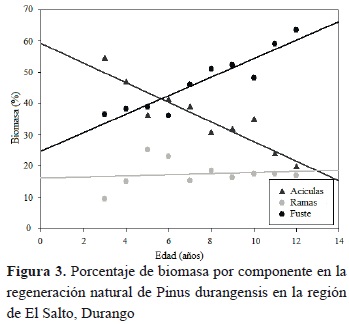
El porcentaje de biomasa promedio (35.64) que presentan las acículas en el intervalo de edad de tres a doce años va de un máximo de 54.26 % a un mínimo de 19.52 %; presentando un coeficiente de variación de 28.89 %. La grafica muestra que a medida que el árbol crece presenta una disminución de biomasa en este componente de 3.14 % por año, además, dicha tendencia puede ser explicada mediante regresión lineal simple (Tabla 3). En las ramas se presenta un porcentaje promedio de 17.38 % de biomasa, variando desde 9.26 % a un máximo de 25.25 %, presentando un coeficiente de variación de 24.96 en el intervalo de tres a doce años. Contrariamente al componente acículas, se observa que a medida que el árbol crece, el porcentaje de biomasa en ramas va aumentando (0.17 % por año; Tabla 4).
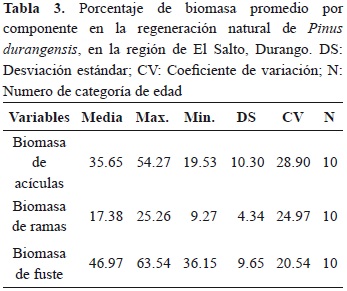
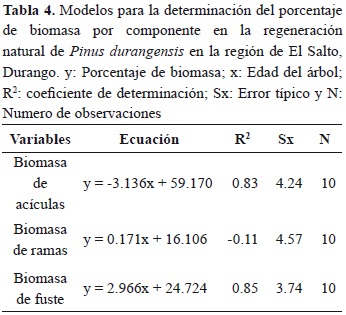
En el fuste el porcentaje de biomasa promedio (46.95 %) que se presenta en el intervalo de edad de tres a doce años cambia de un mínimo de 36.14 % a un máximo de 63.53 %, mostrando menor variación en este componente. La tendencia general indica que a medida que el árbol crece, el porcentaje de biomasa se incrementa, la regresión lineal ajustada a estas observaciones indican que este incremento es del orden de 2.96 % por año.
DISCUSIÓN
Aguilar (1991) realizó una comparación de cuatro modelos matemáticos (Weibull, Schumacher, Gompertz y logístico) aplicados al crecimiento edad-altura en tres calidades de estación en la especie de P. douglasiana. Los modelos Weibull, Schumacher y Gompertz presentan mejor ajuste que otros en diferentes etapas y el modelo de Gompertz presenta una mínima de falta de ajuste. A diferencia con García et al. (1992), donde obtuvieron un coeficiente de determinación de 0.940 y un error cuadrático medio de 0.359, con el modelo lineal para predecir el crecimiento en altura en especies de S. macrophylla en la plantación de �San Felipe-Bacalar� Quintana Roo.
García et al. (1992) determinaron el crecimiento en diámetro de una plantación de la especies de Swietenia macrophylla King (caoba) con el modelo lineal obteniendo un coeficiente de determinación de 0.98 y un error estándar de 0.43.
CONCLUSIONES
Los modelos de crecimiento ajustados a los componentes de P. durangensis indican que los mejores ajustes se registran al diámetro, seguido por altura, carbono total, carbono en acículas, carbono en ramas y finalmente al carbono en el fuste. Los resultados indican que los mejores ajustes se muestran al incluir todo el árbol completo (suma de todos los componentes) seguido por acículas, ramas y finalmente el fuste.
REFERENCIAS BIBLIOGRÁFICAS
Aguilar, R.M. (1991). Comparación de cuatro modelos matematicos aplicados al crecimiento forestal. Revista de Ciencia Forestal, 16(70), 87-107.
Clutter, J., Fortson, J., Pienaar, G., Brister, G., & Bailey, R. (1983). Timber Management: a Quantitative Approach. New York: John Wiley and Sons. 333 p.
Cubillos, D.V. (1987). Modelos de crecimiento diametral para algunos renovales de Raullí. Ciencia e Investigación Forestal, 1, 67-77.
García, C.X., Lezama, P.C., & Santiago, R.B. (1992). Modelos de crecimiento para una plantación de caoba (Swietenia macrophylla King). Revista de Ciencia Forestal, 17(71), 88-102.
Gayoso, J., Guerra, J., & Alarcon, D. (2002). Contenido de carbono y funciones de biomasa en especies nativas y exoticas: Medicion de la capacidad de captura de carbono en bosques de Chile y promoción en el mercado mundial. Valdivia: Universidad Austral de Chile. 157 p.
IPCC [Intergovernmental Panel on Climate Change]. (1994). The supplementary report to the IPPCC scientific assessment. Cambridge: Cambridge University Press. 47 p.
Morales, C.F., & Ruiz, G.V.J. (2004). Estimación de peso verde, peso seco y volumen de fuste mediante la densidad aparente de la madera en Pinus cooperi y Pinus leiophylla. En la región de El salto, Durango (Tesis de pregrado). El Salto, Durango: Instituto Tecnologico Forestal El Salto. 92 p.
Van Laar, A., & Akca, A. (1997). Forest Mensurasión. G�ttingen: Cuvillier Verlag. 418 p.




Licencia
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






