
DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2016.1.a04Publicado:
01-01-2016Número:
Vol. 19 Núm. 1 (2016): Enero-JunioSección:
Artículos de investigación científica y tecnológicaBiomasa de raíces finas y fertilidad del suelo en bosques pluviales tropicales del pacífico colombiano
Fine Root Biomass and Soil Fertility in Tropical Rain Forests of the Colombian Pacific
Palabras clave:
carbon balance, biogeographic Chocó, Opogodó, pacurita, phosphorus, plant nutrition, tropical soils (en).Palabras clave:
Balance del carbono, Chocó biogeográfico, Fósforo, Nutrición vegetal, Opogodó, Pacurita, Suelos tropicales. (es).Descargas
Referencias
Adams, T., McCormack M. L. & Eissenstat, D. M. (2013). Foraging strategies in trees of different root morphology: the role of root lifespan. Tree Physiology, 33, 940 – 948.
Aerts, R. & Chapin III, F. S. (1999). The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns.
Advances in Ecological Research, 30, 1 – 67.
Alvarez-Clare, S., Mack, M. C. & Brooks, M. (2013). A direct test of nitrogen and phosphorus limitation to net primary productivity in a lowland tropical wet forest. Ecology, 94(7), 1540 – 1551.
Barbhuiya, A. R., Arunachalam A., Pandey H. N., Khan M. L.& Arunachalam, K. (2012). Fine root dynamics in undisturbed and disturbed stands of a tropical wet evergreen forest in northeast India. Tropical Ecology, 53(1), 69 – 79.
Barreto, L. H. & León J. D. (2005). Masa total y contenido de nutrientes en raíces finas de ecosistemas forestales (Pinus patula Schltdl y Cham, Cupressus lusitanica Mill y Quercus humboldtii Bonpl.) de Piedras Blancas, Antioquia-Colombia. Revista Facultad Nacional de Agronomía Medellín, 58 (2), 2907 – 2929.
Burke, M.K. & Raynal, D.J. (1994). Fine root growth phenology, production, and turnover in a northern hardwood forest ecosystem. Plant and Soil, 162, 135 – 146.
Cavelier, J. (1992). Fine-root biomass and soil properties in a semideciduous and a lower montane rain forest in Panama. Plant and Soil, 142, 187 – 201.
Chapin III, F.S., Matson, P.A., & Mooney, H.A. (2002). Principles of Terrestrial Ecosystem Ecology. Springer-Verlag New York, Inc. United States of America. 436 p.
Espeleta, J.F. & Clark, D. A. (2007). Multi-Scale Variation in Fine-Root Biomass in a Tropical Rain Forest: A Seven-Year Study. Ecological Monographs, 77(3), 377 – 404.
Finér, L., Ohashi, M., Noguchi, K., & Hirano, Y. (2011). Factors causing variation in fine root biomass in forest ecosystems. Forest Ecology and Management, 261, 265 – 277.
Gardi, C., Angelini, M., Barceló, S., Comerma, J., Cruz Gaistardo, C., Encina Rojas, A., Jones, A., Krasilnikov, P., Mendonça Santos Brefin, M.L., Montanarella, L., Muñiz Ugarte, O., Schad, P., Vara Rodríguez, M.I., & Vargas, R. (eds), (2014). Atlas de suelos de América Latina y el Caribe, Comisión Europea - Oficina de Publicaciones de la Unión Europea, L-2995 Luxembourg, 176 p.
Gower, S.T. (1987). Relations between mineral nutrient availability and fine root biomass in two Costa Rican tropical wet forests: a hypothesis. Biotropica, 19(2), 171– 175.
Green, J.J., Dawson L.A., Proctor, J., Duff, E.I. & Elston, D.A. (2005). Fine root dynamics in a tropical rain forest is influenced by rainfall. Plant and Soil, 276, 23 – 32.
Hendricks, J.J., Nadelhoffer, K.J. & Aber, J.D. (1993). Assessing the role of fine roots in carbon and nutrient cycling. Trends in Ecology and Evolution, 8,174 – 178.
Holdridge, L. P. (1996). Ecología Basada en las Zonas de Vida. San José, Costa Rica: Instituto Interamericano para la Agricultura. 216 p.
Hopkins, M.S., Reddell, P., Hewett, R.K. & Graham, A.W. (1996). Comparison of Root and Mycorrhizal Characteristics in Primary and Secondary Rainforest on a Metamorphic Soil in North Queensland, Australia. Journal of Tropical Ecology, 12 (6), 871 – 885.
Hoshmand, A. R. (1998). Statistical Methods for Environmental & Agricultural Sciences. Second edition. New York: CRC Press LLC. 439 p.
IGAC. Instituto Geográfico Agustín Codazzi. (2002). Mapa de Suelos de Colombia. Recuperado de: http://mapascolombia.igac.gov.co/wps/portal/mapasdecolombia/.
Jackson, R. B., Mooney, H. A. & Schulze. E.D. (1997). A global budget for fine root biomass, surface area, and nutrient contents. Proceedings of the National Academy of Sciences of the United States of America, 94, 7362 – 7366.
James, J.J., Ziegenhagen, L. & Aanderud, Z. T. (2010). Exploitation of Nutrient-Rich Soil Patches by Invasive Annual and Native Perennial Grasses. Invasive Plant Science and Management, 3, 169 – 177.
Jaramillo, V.J., Ahedo-Hernández, R. & Kauffman, J. B. (2003). Root biomass and carbon in a tropical evergreen forest of Mexico: changes with secondary succession and forest conversion to pasture. Journal of Tropical Ecology, 19, 457 – 464.
Jiménez, E.L., Moreno-Hurtado, F., Peñuela, M.C., Lloyd, J., Patiño, S., & Álvarez. E. (2008). Estimación de la masa de raíces finas en dos bosques de tierra firme sobre suelos contrastantes en la Amazonia colombiana. Pp. 165 – 196. En: Buitrago, A.I., &
Jiménez, E.M. (Eds.). Gente, Tierra y Agua: Imani Mundo III. Universidad Nacional de Colombia Sede Amazonia. Instituto Amazónico de Investigaciones IMANI. Bogotá: Editorial Guadalupe Ltda. 240 p.
Kochsiek, A., Tan, S. & Russo, S.E. (2013). Fine root dynamics in relation to nutrients in oligotrophic Bornean rain forest soils. Plant Ecology, 214(5), 1385 – 0237.
Kummerow, J., Castillanos, J. Maas, M. & Larigauderie, A. (1990). Production of fine roots and the seasonality of their growth in a Mexican deciduous dry forest. Vegetatio, 90, 73 – 80.
Malagon D., Pulido, C., Llinas, R. D., Chamorro, C. & Fernández, J. (1995). Suelos de Colombia. Origen, Evolución, Clasificación, Distribución y uso. Bogotá: Instituto Geográfico Agustín Codazzi. Subdirección de Agrología. 632 p.
Martínez, J. O. (1993). Geomorfología. En: Leyva, P. (Ed.). Colombia Pacífico, Tomo I. Fondo para la Protección del Medio Ambiente “José Celestino Mutis” FEN Colombia.
Maycock, C.R. & Congdon, R.A. (2000). Fine Root Biomass and Soil N and P in North Queensland Rain Forests. Biotropica, 32(1), 185 – 190.
Metcalfe, D. B., Meir, P., Aragao, L. E., Da Costa, A. C. L., Braga, A. P., Gonḉalves, P. H. L., De Athaydes Silva Jr., J., De Almeida, S. S., Dawson, L. A., Malhi, Y., & Williams, M. (2008). The effects of water availability on root growth and morphology in an Amazon rainforest. Plant Soil, 311, 189 – 199.
Moreno-Hurtado, F. (2004). Soil Carbon Dynamics in Primary and Secondary Tropical Forests in Colombia. Thesis of Doctor of Philosophy in Biology. Miami: Florida International University. 208 p.
Osorio, N. W. (2014). Manejo de nutrientes en suelos del Trópico. Segunda edición. Medellín: Editorial L. Vieco S.A.S. 416 p.
Ostertag, R. (2001). Effects of nitrogen and phosphorus availability on fine-root dynamics in Hawaiian montane forests. Ecology, 82(2), 485–499.
Poveda, I.C., Rojas, C., Rudas, A., & Rangel, O. (2004). El Chocó biogeográfico: Ambiente Físico. En Rangel, O. Colombia Diversidad Biótica IV. El Chocó biogeográfico/ Costa Pacífica. Instituto de Ciencias Naturales. Bogotá: Universidad Nacional de Colombia. 1024 p.
Powers, J. S., Treseder, K. K. & Lerdau, M. T. (2005). Fine roots, arbuscular mycorrhizal hyphae and soil nutrients in four neotropical rain forests: patterns across large geographic distances. New Phytologist, 165, 913 – 921.
Quinto, H., & Moreno, F. H. (2014). Diversidad florística arbórea y su relación con el suelo en un bosque pluvial tropical del Chocó Biogeográfico. Revista Árvore, 38 (6), 1123 -1132.
R Development Core Team. (2012). R: A language and environment for statistical computing. Vienna, Austria. R Foundation for Statistical Computing. ISBN: 3-900051-07-0. Recuperado de http://www.r-project.org/.
Ruiz – Murcia, J. F. (2010). Cambio Climático en Temperatura, Precipitación y Humedad Relativa para Colombia Usando Modelos Meteorológicos de Alta Resolución (Panorama 2011-2100). Instituto de Hidrología, Meteorología y Estudios Ambientales – IDEAM. Subdirección de Meteorología. Nota Técnica IDEAM–METEO/005-2010. Bogotá, D. C., Mayo (2010). 91 p.
Sanford, R. L. & Cuevas, E. (1996). Root growth and rhizosphere interactions in tropical forests. En: Mulkey, S. S.; Chazdon, R. L and Smith, A. P. (Eds). Tropical forest plant ecophysiology. New York: Chapman and Hall. p. 268 – 300.
Saugier, B., Roy, J. & Mooney, H.A. (2001). Estimations of global terrestrial productivity: Converging toward a single number? in Roy, J., Saugier, B. & Mooney, H.A. (Eds). Terrestrial Global Productivity. San Diego, CA: Academic Press. 543–557.
Sayer, E. J., Wright, S. J., Tanner, E. V. J., Yavitt, J. B., Harms, K. E., Powers, J. S., Kaspari, M., Garcia, M. N., & Turner, B. L. (2012). Variable Responses of Lowland Tropical Forest Nutrient Status to Fertilization and Litter Manipulation. Ecosystems, 15, 387 – 400.
Silver, W. L., Thompson, A. W., Mcgroddy, M. E., Varner, R. K., Dias, J. D., Silva, H., Crill, P. M., & Keller, M. (2005). Fine root dynamics and trace gas fluxes in two lowland tropical forest soils. Global Change Biology, 11, 290 – 306.
Silver, W.L., Neff, J., McGroddy, M., Veldkamp, E., Keller, M. & Cosme, R. (2000). Effects of Soil Texture on Belowground Carbon and Nutrient Storage in a Lowland Amazonian Forest Ecosystem. Ecosystems, 3, 193 – 209.
Stark, N.M. & Jordan, C.F. (1978). Nutrient retention by the root mat of an Amazonian Rain Forest. Ecology, 59(3), 434 – 437.
Statistical Graphics Corp. (2002).StatgraphicsPlusCenturium Version 5.1. Recuperado de http://www.Statgraphics.com.
Trumbore, S., Da Costa, E. S., Nepstad, D. C., De Camargo, P. B., Martinelli, L., Ray, D., Restom, T., & Silver, W. (2006). Dynamics of fine root carbon in Amazonian tropical ecosystems and the contribution of roots to soil respiration, Global Change Biology, 12, 217 – 229.
Vitousek, P.M. (1984). Litterfall, Nutrient Cycling, and Nutrient Limitation in Tropical Forests. Ecology, 65 (1), 285 – 298.
Vitousek, P.M. & Sanford. R. L. (1986). Nutrient cycling in moist tropical forest. Annual Review of Ecology and Systematics, 17, 137 – 167.
Vogt, K.A., Grier, C.C. & Vogt, D.J. (1985). Production, turnover, and nutrient dynamics of above- and below-ground detritus of world forests. Advances in Ecological Research, 15, 303– 377.
West, R. (1957). Las tierras bajas del Pacífico colombiano. Instituto Colombiano de Antropología. Bogotá: Imprenta Nacional de Colombia. .300 pp.
Wright, S.J., Yavitt, J.B., Wurzburger, N., Turner, B. L., Tanner, E.V.J., Sayer, E.J., Santiago, L.S., Kaspari, M., Hedin, L.O., Harms, K.E., Garcia, M.N. & Corre, M.D. (2011). Potassium, phosphorus, or nitrogen limit root allocation, tree growth, or litter production in a lowland tropical forest. Ecology, 92(8), 1616 – 1625.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
 |
Quinto M, H., Moreno H, H., Caicedo M, H. & Pérez L, M. (2016). Biomasa de Raíces Finas y Fertilidad del Suelo en Bosques Pluviales Tropicales del Pacífico Colombiano. Colombia Forestal, 19(1), 53-67. |
Artículo de investigación
BIOMASA DE RAÍCES FINAS Y FERTILIDAD DEL SUELO EN BOSQUES PLUVIALES TROPICALES DEL PACíFICO COLOMBIANO
Fine Root Biomass and Soil Fertility in Tropical Rain Forests of the Colombian Pacific
Harley Quinto Mosquera1, Flavio Moreno Hurtado2, Haylin Yineth Caicedo Moreno3 & May Thelis Yineth Perez Luis4
1Facultad de Ciencias Básicas. Universidad Tecnológica del Chocó “Diego Luis Córdoba”. Quibdó, Chocó, Colombia. hquintom@gmail.com. Autor de correspondencia.
2Departamento de Ciencias Forestales. Facultad de Ciencias Agrarias. Universidad Nacional de Colombia Sede Medellín. fhmoreno@unal.edu.co.
3Facultad de Ciencias Básicas. Universidad Tecnológica del Chocó “Diego Luis Córdoba”. Quibdó, Chocó, Colombia
4Facultad de Ciencias Básicas. Universidad Tecnológica del Chocó “Diego Luis Córdoba”. Quibdó, Chocó, Colombia
RESUMEN
Las raíces finas juegan un papel importante en la ecología y dinámica de los bosques tropicales. La biomasa de raíces finas (BRF) está determinada principalmente por las características del suelo (disponibilidad de agua y nutrientes) y es mayor en suelos con menor fertilidad. Para evaluar esta hipótesis se establecieron cinco parcelas permanentes de una hectárea en las localidades de Opogodó (Condoto) y Pacurita (Quibdó), Chocó, Colombia, donde se midió la BRF a 0-10 y 10-20 cm de profundidad, y la fertilidad edáfica (pH, nutrientes y textura), los cuales se relacionaron con la BRF. Ambas localidades presentaron suelos pobres en nutrientes, con mayor contenido de arena, N total y materia orgánica (MO) en Opogodó. La BRF presentó poca relación con la textura y el contenido de nutrientes del suelo, pues solo las correlaciones de la BRF con el pH y el contenido de arcilla fueron significativas. En ambas zonas la BRF disminuyó con la profundidad; los valores encontrados entre 0 - 20 cm de profundidad fueron de 5.91 y 6.28 t ha-1 en Opogodó y Pacurita, respectivamente. El análisis a escala de la región tropical mostró una relación inversa entre el contenido del P disponible y la BRF de los bosques.
Palabras clave: balance del carbono, Chocó biogeográfico, fósforo, nutrición vegetal, Opogodó, Pacurita, suelos tropicales.
ABSTRACT
Fine roots play an important role in the ecology and dynamics of tropical forests. Fine root biomass (FRB) is mainly determined by soil characteristics (availability of water and nutrients) and is higher in soils with lower fertility. To test this hypothesis five permanent 1-hectare plots were established in the towns of Opogodó and Pacurita, where FRB was measured at 0-10 and 10-20 cm depth. In addition, soil fertility parameters (pH, nutrients and texture) were measured and related to FRB. Both towns presented soils poor in nutrients, with higher content of sand, total N and organic matter (OM) in Opogodó. The BRF presented little relation to texture and soil nutrient content, because only the correlations of the BRF with pH and content of clay were significant. In both areas FRB decreased with depth; values found between 0-20 cm depths were 5.91 t ha-1 and 6.28 t ha-1 in Opogodó and Pacurita respectively. The tropical region-wide analysis showed an inverse relationship between the available P content and the BRF of forests.
Keywords: carbon balance, biogeographic Chocó, Opogodó, pacurita, phosphorus, plant nutrition, tropical soils.
INTRODUCCIÓN
Las raíces son componentes fundamentales en el balance del carbono de los bosques húmedos tropicales pues representan entre 30 y 40% de la productividad primaria neta y pueden llegar a contener hasta un 22% de la biomasa total del bosque (Saugier et al., 2001; Chapin III et al., 2002). Se suelen dividir en raíces finas (RF) (<5 mm de diámetro) y gruesas (>5 mm de diámetro); las primeras son responsables principalmente de la absorción de agua y nutrientes del suelo, mientras que las segundas proporcionan anclaje y sostén a las plantas. En los bosques tropicales han sido más abundantes los estudios sobre biomasa de raíces finas (BRF) que los de raíces gruesas, debido en primer lugar a las dificultades implícitas en el muestreo de estas últimas, y en segundo lugar a la importancia de las RF para el funcionamiento del ecosistema, no sólo por la adquisición de recursos para las plantas, sino también por su participación en el almacenamiento de carbono y en los ciclos biogeoquímicos del bosque (Burke & Raynal, 1994; Sanford & Cuevas, 1996; Barreto & León, 2005).
La BRF evidencia en gran medida la capacidad de las plantas para capturar recursos del suelo porque es proporcional al volumen de suelo explorado. La BRF varía ampliamente en bosques tropicales, con un rango de entre 3 y 120 t ha-1 (Vogt et al., 1985), se concentran principalmente en las capas superficiales del suelo (los primeros 30 cm de profundidad pueden almacenar hasta el 57% de la BRF total del bosque) (Jackson et al., 1997). Se ha documentado que la BRF está relacionada con condiciones favorables para la fotosíntesis, la estructura y composición florística del bosque, disponibilidad de agua y nutrientes y aireación del suelo (Silver et al., 2000; Barreto & León, 2005).
Con base en algunos estudios sobre BRF realizados en bosques húmedos tropicales, se ha propuesto la hipótesis que en suelos fértiles o ricos en nutrientes se presenta menor BRF en comparación con los suelos infértiles o limitados por algunos nutrientes (Gower, 1987; Aerts & Chapin III., 1999; Maycock & Congdon, 2000). En bosques húmedos tropicales también se han reportado relaciones inversamente proporcionales entre la BRF y la disponibilidad de nutrientes (Powers et al., 2005; Espeleta & Clark, 2007; Kochsiek et al., 2013). Estas se explican parcialmente por el aumento en las alteraciones morfológicas, por el incremento de la vida media de las raíces finas bajo condiciones de baja fertilidad edáfica (Powers et al., 2005), así como también por la mayor asignación de biomasa subterránea (Hendricks et al., 1993) que se presenta como estrategia para maximizar la captura de recursos del ecosistema bajo condiciones adversas (hipótesis de asignación diferencial).
A pesar de que se han realizado numerosos estudios que evalúan las existencias de BRF y su relación con los factores edáficos en bosques tropicales (Gower, 1987; Maycock & Congdon, 2000; Powers et al., 2005; Espeleta & Clark, 2007; Kochsiek et al., 2013), en regiones de excesiva precipitación, con suelos fuertemente lixiviados y de baja fertilidad como los que se presentan en el Pacífico colombiano donde la precipitación anual puede superar los 10 000 mm, tales estudios son prácticamente inexistentes. Sobre la base de la premisa que la BRF responde a la fertilidad del suelo surgen los siguientes interrogantes: ¿De qué magnitud serán las existencias de BRF en los suelos de baja fertilidad como los del Chocó biogeográfico? ¿Qué tanto las cantidades de BRF son explicadas por las características físicas y químicas de suelos sometidos a condiciones de alta precipitación? Responder estas preguntas en dos bosques pluviales tropicales del Pacífico colombiano fue el objetivo principal del presente estudio.
MATERIALES Y MÉTODOS
Área de estudio
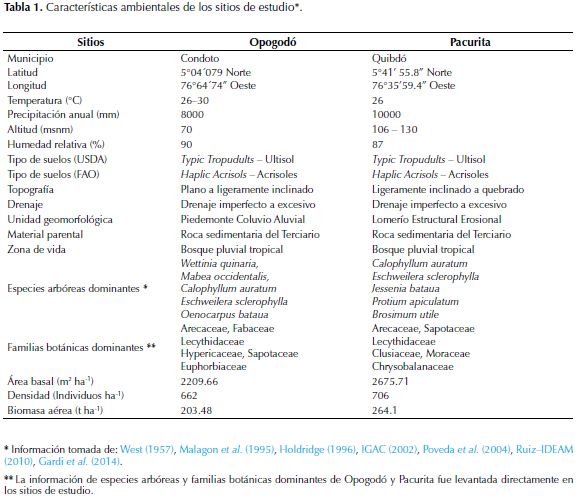
El presente estudio se realizó en bosques pluviales tropicales de la localidad de Pacuríta (municipio de Quibdó) y en la localidad de Opogodó (municipio de Condoto), departamento del Chocó, Colombia. Estas dos localidades pertenecen a la subregión ecogeográfica Central Norte del Chocó biogeográfico, que comprende las cuencas altas de los ríos Atrato y San Juan (Poveda et al., 2004). Las localidades se encuentran dentro de la unidad geomorfológica de Colinas sedimentarias del Terciario, que se caracterizan por presentar bajas altitudes, formadas por rocas sedimentarias, compuestas por arcillolitas arenosas, areniscas y calizas (West, 1957; Martínez, 1993). Las características ambientales y estructurales específicas de Opogodó y Pacurita se detallan en la tabla 1.
En la localidad de Opogodó el muestreo se realizó en tres parcelas permanentes de una ha, instaladas dos kilómetros de la margen derecha de la carretera entre Opogodó y Novita, en predios de la Universidad Tecnológica del Chocó Diego Luis Córdoba. En la localidad de Pacurita el estudio se realizó en dos parcelas permanentes establecidas en una zona de reserva forestal denominada Estación Biológica Pacurita, ubicada a 6.5 km del municipio de Quibdó, en la margen izquierda de la carretera entre Quibdó y Pacurita. En ambas localidades el muestreo se realizó en bosques primarios bien conservados.
Establecimiento de Parcelas
Las cinco parcelas permanentes se establecieron en el año 2013 y cada una consiste de un cuadrado de 100 x 100 m dividido en 25 subparcelas de 20 x 20 m (400 m2); a su vez las subparcelas se subdividieron en cuadrados de 10 x 10 m, en los cuales se tomaron las muestras de suelos y de BRF.
Biomasa de raíces finas (BRF)
Para la determinación de la BRF se colectaron muestras de suelo a dos profundidades (de 0-10 y de 10-20 cm), con un barreno de suelos Eijkelkamp® de 8 cm de diámetro y 15 cm de profundidad. Las muestras se tomaron en el centro de los cuadrados de 10 x 10 m que dividen las subparcelas. En cada subparcela se tomaron ocho muestras de suelo, para un total de 200 muestras por parcela y un gran total de 1000 muestras de raíces en las 5 parcelas.
En el campo se realizó una primera separación manual de las raíces con la ayuda de tamices (de 1 mm de amplitud) y bandejas plásticas; después de la separación el suelo restante fue depositado nuevamente en cada uno de los orificios donde se tomó la muestra. Las muestras de raíces fueron llevadas al Laboratorio de Botánica y Ecología de la Universidad Tecnológica del Chocó D.L.C., donde se les retiró la arena y arcilla remanentes con tamices de diferentes calibres (de 0.5 y 1 mm de amplitud) y agua a diferentes presiones. Las muestras de raíces finas (diámetro ≤ 5 mm) obtenidas mediante este procedimiento fueron posteriormente secadas a 70° C durante 48 horas en un horno de secado de circulación forzada Acequilab Ltda® y pesadas en una balanza analítica (0.001 g de precisión). La BRF se determinó como el peso seco de la muestra y los valores obtenidos se expresaron en toneladas por ha.
Análisis de suelos
Para evaluar la fertilidad del suelo se emplearon muestras compuestas tomadas en las cuatro esquinas y en el centro de las subparcelas y colectadas en la capa de 0- 20 cm de profundidad; para ello se retiró previamente la hojarasca y otro material orgánico no descompuesto de la superficie. En cada parcela se tomaron 25 muestras de suelo, para un total de 125 muestras en todo el estudio. Los análisis se realizaron en el laboratorio de Biogeoquímica de la Universidad Nacional de Colombia Sede Medellín, mediante las siguientes técnicas: textura con el método de Bouyoucos, pH con Potenciómetro de suelo: agua 1:2; materia orgánica (MO) con el método de Walkley y Black y Volumetría; Nitrógeno con el método de Micro- Kjeldahl; Fósforo con Acido L ascórbico y espectrofotómetro UV - VIS; y Ca, Mg, K con el método de Acetato de amonio 1N, neutro y absorción atómica (Osorio, 2014).
Análisis estadístico de los datos
Se evaluaron los supuestos de normalidad y homogeneidad de varianzas con los estadísticos de Bartlett, Hartley y Kurtosis(Hoshmand, 1998) antes de evaluar la correlación entre la BRF y la fertilidad del suelo (Textura, MO, pH, Al, P, Ca, K, Mg, CICE). En virtud de que estos supuestos no se cumplieron, tal correlación se evaluó mediante el Coeficiente de Correlación por Rangos de Spearman (Rs). Así mismo, la variación de la BRF en función de las localidades y las profundidades del suelo se evaluó mediante la prueba no paramétrica de Mann-Whitney-Wilcoxon (W)(Hoshmand, 1998). Posteriormente, las relaciones lineales entre todas las variables incluidas en este estudio se evaluaron mediante un Análisis de Componentes Principales (ACP).
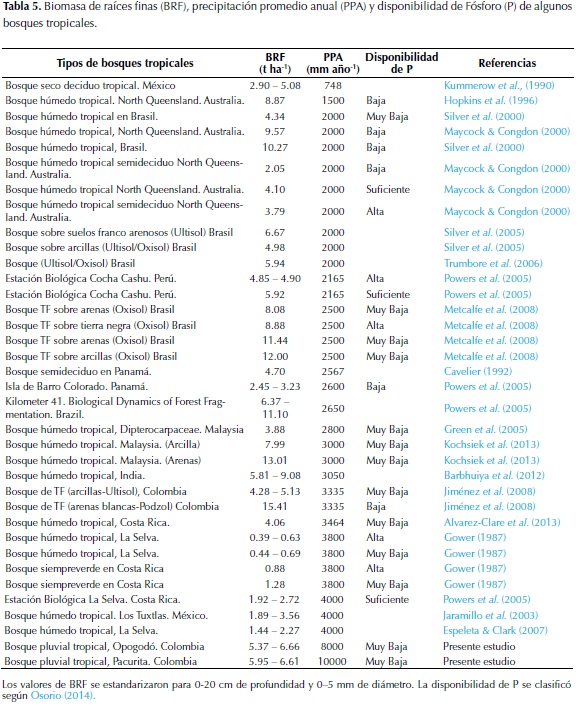
Finalmente, la relación entre la BRF y variables ambientales (precipitación y P disponible) de distintos bosques húmedos tropicales de baja altitud (incluidos Pacurita y Opogodó) se evaluó mediante Regresiones Lineales. Puesto que algunos estudios consideran que el límite de diámetro de las RF es de 2 mm, para estos análisis los datos de BRF se estandarizaron a un diámetro < 5 mm, basados en el hecho de que la BRF < 2 mm es 29% menor que la BRF < 5 mm de diámetro (Finér et al., 2011). Así mismo, se emplearon los estudios de Moreno (2004) y Espeleta & Clark (2007) para calcular los porcentajes de BRF a distintas profundidades y estandarizarlas a una profundidad de 20 cm, dado que los estudios reportan mediciones a diferente profundidad en el suelo. Los valores de nitrógeno fueron excluidos de los análisis porque se consideró que este nutriente no es limitante en bosques húmedos tropicales de baja altitud, pues sus concentraciones edáficas son por lo general altas en comparación con otros nutrientes como el P (Vitousek, 1984). Los análisis se realizaron con los programas Statgraphics Centurion XV (Statistical Graphics Corp., 2002) y The R Project for Statistical Computing (R Development Core Team. 2012).
RESULTADOS
Fertilidad básica de los suelos
Los suelos estudiados son extremadamente ácidos; con mayor acidez y porcentajes altos de saturación de Al en la localidad de Pacurita (Saturación de Al = 57.21%). Se presentaron concentraciones altas de MO y N, con los mayores registros en Opogodó (MO = 11.94%; N = 0.61%). Las cantidades edáficas de P, Mg, y Ca fueron muy bajas, mientras que los valores de K fueron intermedios; así mismo, la CICE fue baja en las dos zonas. Las concentraciones de P y Ca fueron muy similares en las dos localidades; sin embargo, el resto de los parámetros de fertilidad presentaron diferencias estadísticamente significativas (Tabla 2). Por lo tanto, basados en la menor acidez, altas concentraciones de MO y N, y en los valores intermedios de K, se considera que los suelos de Opogodó son más fértiles que los de Pacurita.
Biomasa de raíces finas
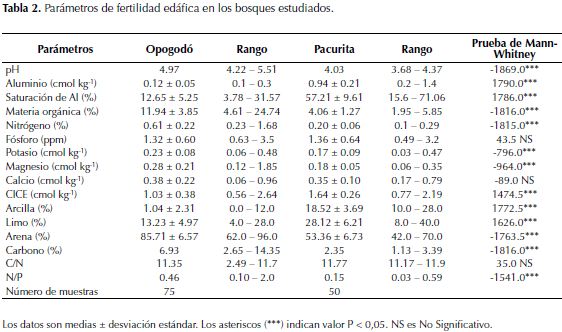
En los bosques de Opogodó se registró una BRF promedio (± D.E.) de 5.91 ± 1.84 t ha-1 entre 0 y 20 cm de profundidad, de la cual el 59% se encontró en los primeros 10 cm. En los suelos de Pacurita se registró una BRF de 6.28 ± 1.66 t ha-1, de la cual el 63% se encontró en los primeros 10 cm (Tabla 3). La BRF presentó diferencias entre profundidades; pero no entre localidades.
Relación entre la biomasa de raíces finas y la fertilidad del suelo
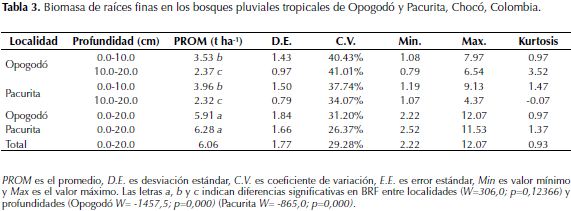
Al analizar los datos de las dos localidades en conjunto se observó que la BRF no presentó correlación significativa con los parámetros edáficos evaluados. El análisis individual en cada localidad mostró correlaciones significativas con algunas variables edáficas, pero muy débiles: en Opogodó la BRF presentó correlación negativa con la arcilla, mientras que en Pacurita la BRF presentó correlación positiva con el pH.
El ACP mostró que los dos primeros componentes fueron significativos y explicaron el 92.8 % de la varianza de los datos. Las condiciones edáficas (textura, pH, N total y MO) tuvieron alta correlación entre sí y diferenciaron claramente las parcelas de Opogodó de las de Pacurita. Sin embargo, la BRF (total y en las dos profundidades) presentó poca relación con las variables edáficas (Figura 1).
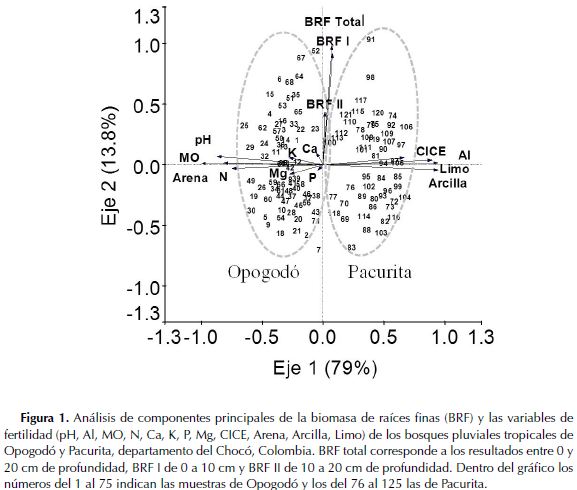
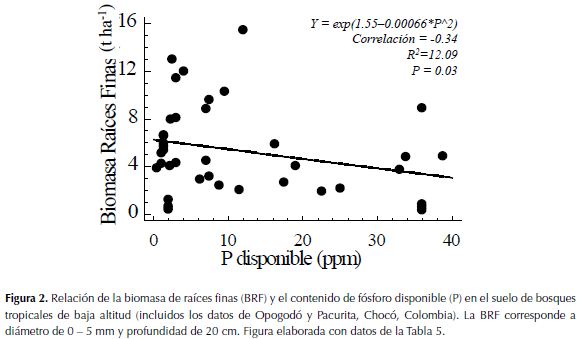
Por otra parte, al relacionar la BRF de distintos bosques húmedos tropicales (incluidos los datos de este estudio), con la precipitación anual y el contenido de P disponible del suelo (Tabla 5), se encontró que el P disponible del suelo explica parcialmente la BRF (Figura 2), pero la precipitación anual no (R2= 2.06; P= 0.34). En síntesis, se evidenció que en los trópicos la BRF presenta una relación significativa inversamente proporcional con el P disponible del suelo, pero aparentemente es independiente de los niveles de precipitación promedia anual.
DISCUSIÓN
Biomasa de raíces finas en bosques pluviales tropicales
La BRF registrada entre 0 y 20 cm de profundidad, en suelos arenosos de los bosques pluviales tropicales de Opogodó y en suelos arcillosos de Pacurita, se encuentra dentro de los rangos reportados en bosques tropicales de tierras bajas con valores de entre 3 - 120 t ha-1 (Vogt et al., 1985). Nuestros resultados fueron similares a los registrados en otros bosques tropicales de baja altitud con diferentes regímenes de precipitación en Australia (Maycock & Congdon 2000), Brasil (Silver et al., 2000; 2005), Perú (Powers et al., 2005), Colombia (Jiménez et al., 2008), India (Barbhuiya et al., 2012), Costa Rica (Alvarez-Clare et al., 2013) y Malaysia (Kochsiek et al., 2013); así como también a los registrados en bosques semideciduos de Panamá (Cavelier, 1992) y en bosques secos de México (Kummerow et al., 1990) (Tabla 5). El hecho de que estos bosques tropicales con diferentes niveles de precipitación presenten una BRF similar, muestra que otros factores como la disponibilidad de nutrientes (por ejemplo el P disponible - Figura 1) (Gower, 1987; Maycock & Congdon, 2000; Espeleta & Clark, 2007), textura (Silver et al., 2000; 2005; Jiménez et al., 2008; Kochsiek et al., 2013), material parental (Espeleta & Clark, 2007), tipo de vegetación y área basal (Finér et al., 2011), entre otros, podrían tener un mayor efecto sobre la BRF en comparación con la precipitación anual. Al respecto, Finér et al., (2011), afirman que factores como temperatura, precipitación promedia anual, latitud y altitud, explican muy poco las variaciones en la BRF de los bosques tropicales.
La mayor BRF registrada en los primeros 10 cm del suelo en este estudio (Tabla 3) es similar a lo reportado en estudios realizados en bosques tropicales de baja altitud (Kummerow et al., 1990; Cavelier 1992; Silver et al., 2000; Maycock & Congdon, 2000; Jiménez et al., 2008; Barbhuiya et al., 2012; Alvarez-Clare et al., 2013; Kochsiek et al., 2013). Tal concentración de RF en los primeros centímetros del suelo posiblemente se debe al hecho de que en la superficie se presenta mayor disponibilidad de nutrientes provenientes de la descomposición de la hojarasca, por lo cual una mayor BRF superficial facilita la captura de iones solubles directamente de la MO y evita su pérdida por lixiviación (Stark & Jordan 1978; Kummerow et al., 1990). Particularmente, en la zona de estudio, la mayor BRF superficial probablemente está muy influenciada por la alta precipitación que provoca fuerte pérdida y lixiviación de nutrientes, frente a lo cual las plantas desarrollan un entramado de RF superficiales como estrategia de conservación de nutrientes.
Relación entre la biomasa de raíces finas y la fertilidad del suelo
El análisis de la relación entre las condiciones edáficas y la BRF a escala local en los bosques estudiados mostró que la variación en BRF fue poco explicada por los cambios en las condiciones edáficas como textura, acidez y concentración de nutrientes del suelo (Figura 1; Tabla 4). Esta baja relación puede deberse a varias razones: en primer lugar, la variación de los parámetros de fertilidad de los suelos fue baja (Tabla 2), pues los suelos de todas las subparcelas fueron muy infértiles. Si bien varios estudios han reportado variación de la BRF con la fertilidad del suelo (Gower, 1987; Maycock & Congdon, 2000; Espeleta & Clark 2007; Kochsiek et al., 2013), ello es detectable solamente cuando existe un gradiente de fertilidad más marcado, lo cual no ocurrió en el presente estudio, ya que a pesar de que los sitios estudiados difirieron en la mayoría de los parámetros utilizados para evaluar la fertilidad edáfica, en ambas zonas los suelos fueron infértiles.
La segunda razón, la alta diversidad de especies arbóreas de la región del Chocó biogeográfico (≈200 especies por ha, datos no publicados), así como las variaciones marcadas en la estructura (área basal y biomasa) del bosque posibilitan la existencia de múltiples estrategias para la captura de nutrientes, puesto que las especies responden de manera distinta a la disponibilidad de nutrientes del suelo (Aerts & Chapin III., 1999; James et al., 2010; Adams et al., 2013); por ejemplo, en bosques pluviales cercanos a la zona de estudio se observó que la distribución de las especies arbóreas es explicada en un 37% por las variables espaciales y ambientales (edáficas y topográficas), incluida la disponibilidad de nutrientes (Quinto & Moreno, 2014). Lo cual, evidencia parcialmente porqué a nivel de comunidad puede resultar difícil encontrar relaciones significativas entre BRF y condiciones edáficas. Similares resultados han sido reportados por Alvarez-Clare et al., (2013), en bosques húmedos tropicales de baja altitud en Costa Rica, quienes no encontraron una respuesta de la BRF del ecosistema a la adición de nutrientes (N y P) al suelo.
En tercer lugar, como se mencionó anteriormente, por medio de la producción y descomposición de hojarasca se incorporan al suelo grandes cantidades de nutrientes (Stark & Jordan, 1978; Vitousek & Sanford, 1986) que incluso pueden ser superiores a las cantidades disponibles en el suelo. Por ejemplo, el ingreso de P vía hojarasca (1.55 ppm de P disponible) en suelos tropicales infértiles (Vitousek & Sanford, 1986) puede llegar a ser mayor que el registrado hasta 20 cm de profundidad en suelos de Opogodó y Pacurita (Tabla 2); es decir, gran parte de los nutrientes requeridos por las plantas pueden ser obtenidos directamente de la hojarasca en descomposición (Stark & Jordan 1978), lo cual genera cierto grado de independencia de la BRF sobre las condiciones del suelo mineral.
Contrario a lo anterior, al analizar la relación entre la BRF y el contenido edáfico de P disponible a escala regional (bosques tropicales de baja altitud) se denotó una relación significativa e inversamente proporcional entre las variables (Figura 2), lo cual corrobora la hipótesis de la disminución de la BRF en suelos con mayor contenido de nutrientes, como lo reportaron Gower (1987), Maycock & Congdon (2000), Powers et al. (2005), Espeleta & Clark (2007) y Kochsiek et al. (2013). Finalmente, basados en la hipótesis de Gower (1987) que considera que la BRF es controlada por nutriente(s) mineral(es) limitante(s) en el ecosistema forestal, el presente estudio aporta evidencias para considerar al P disponible como uno de los nutrientes más limitantes de los bosques tropicales a escala regional.
Finamente, a pesar de que en el presente estudio se registró un gradiente de textura (de arenosa a arcillosa) entre los suelos de Opogodó y Pacurita, respectivamente (Figura 1), dicho gradiente no afectó la variabilidad de la BRF. Sin embargo, diversos estudios realizados en bosques húmedos tropicales han encontrado una influencia fuerte de la variación de la textura sobre la BRF (Silver et al., 2000; Jiménez et al., 2008; Kochsiek et al., 2013). No obstante, en dichos estudios las variaciones de textura estuvieron acompañadas de cambios en la disponibilidad de nutrientes, especialmente P disponible (Silver et al., 2000; Jiménez et al., 2008), situación que no se presentó en los suelos del presente estudio, donde no hubo una variación significativa del P disponible (Tabla 2); esta situación probablemente explica el poco efecto del gradiente de textura sobre la BRF observada en este estudio.
CONCLUSIONES
El presente estudio permitió evidenciar la influencia del contenido edáfico del P disponible sobre la BRF en bosques tropicales a escala regional. A escala local no se evidenció tal influencia, debido a la baja variabilidad de la fertilidad de los suelos estudiados, y probablemente a la influencia de los ingresos de nutrientes al suelo a través de la hojarasca y de otras fuentes aéreas y por la alta diversidad de especies de estos bosques. Para evaluar tal influencia a escala local probablemente sea necesario adicionar nutrientes al suelo, como lo han hecho Ostertag (2001), Wright et al. (2011) y Sayer et al. (2012); con tal adición se generaría un verdadero gradiente de fertilidad que posiblemente conlleve a un cambio en la asignación de biomasa subterránea y a una redistribución del carbono del bosque.
AGRADECIMIENTOS
Este proyecto fue financiado por el Convenio 0010-2013 firmado entre la Universidad Tecnológica del Chocó Diego Luis Córdoba, la Universidad Nacional de Colombia Sede Medellín y el Departamento Administrativo de Ciencia, Tecnología e Innovación, COLCIENCIAS, en el marco del proyecto titulado: “Evaluación del efecto de la fertilización del suelo sobre la productividad primaria neta de bosques pluviales tropicales del departamento del Chocó (CÓDIGO: 1128-569-35113)”. Agradecemos la hospitalidad y colaboración de los habitantes de Opogodó y Pacurita.
REFERENCIAS BIBLIOGRÁFICAS
Adams, T., McCormack M. L. & Eissenstat, D. M. (2013). Foraging strategies in trees of different root morphology: the role of root lifespan. Tree Physiology, 33, 940 - 948.
Aerts, R. & Chapin III, F. S. (1999). The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns. Advances in Ecological Research, 30, 1 - 67.
Alvarez-Clare, S., Mack, M. C. & Brooks, M. (2013). A direct test of nitrogen and phosphorus limitation to net primary productivity in a lowland tropical wet forest. Ecology, 94(7), 1540 - 1551.
Barbhuiya, A. R., Arunachalam A., Pandey H. N., Khan M. L.& Arunachalam, K. (2012). Fine root dynamics in undisturbed and disturbed stands of a tropical wet evergreen forest in northeast India. Tropical Ecology, 53(1), 69 - 79.
Barreto, L. H. & León J. D. (2005). Masa total y contenido de nutrientes en raíces finas de ecosistemas forestales (Pinus patula Schltdl y Cham, Cupressus lusitanica Mill y Quercus humboldtii Bonpl.) de Piedras Blancas, Antioquia-Colombia. Revista Facultad Nacional de Agronomía Medellín, 58 (2), 2907 - 2929.
Burke, M.K. & Raynal, D.J. (1994). Fine root growth phenology, production, and turnover in a northern hardwood forest ecosystem. Plant and Soil, 162, 135 - 146.
Cavelier, J. (1992). Fine-root biomass and soil properties in a semideciduous and a lower montane rain forest in Panama. Plant and Soil, 142, 187 - 201.
Chapin III, F.S., Matson, P.A., & Mooney, H.A. (2002). Principles of Terrestrial Ecosystem Ecology. Springer-Verlag New York, Inc. United States of America. 436 p.
Espeleta, J.F. & Clark, D. A. (2007). Multi-Scale Variation in Fine-Root Biomass in a Tropical Rain Forest: A Seven-Year Study. Ecological Monographs, 77(3), 377 - 404.
Finér, L., Ohashi, M., Noguchi, K., & Hirano, Y. (2011). Factors causing variation in fine root biomass in forest ecosystems. Forest Ecology and Management, 261, 265 - 277.
Gardi, C., Angelini, M., Barceló, S., Comerma, J., Cruz Gaistardo, C., Encina Rojas, A., Jones, A., Krasilnikov, P., Mendonça Santos Brefin, M.L., Montanarella, L., Muñiz Ugarte, O., Schad, P., Vara Rodríguez, M.I., & Vargas, R. (eds), (2014). Atlas de suelos de América Latina y el Caribe, Comisión Europea - Oficina de Publicaciones de la Unión Europea, L-2995 Luxembourg, 176 p.
Gower , S.T. (1987). Relations between mineral nutrient availability and fine root biomass in two Costa Rican tropical wet forests: a hypothesis. Biotropica, 19(2), 171- 175.
Green, J.J., Dawson L.A., Proctor, J., Duff, E.I. & Elston, D.A. (2005). Fine root dynamics in a tropical rain forest is influenced by rainfall. Plant and Soil, 276, 23 - 32.
Hendricks, J.J., Nadelhoffer, K.J. & Aber, J.D. (1993). Assessing the role of fine roots in carbon and nutrient cycling. Trends in Ecology and Evolution, 8,174 - 178.
Holdridge, L. P. (1996). Ecología Basada en las Zonas de Vida. San José, Costa Rica: Instituto Interamericano para la Agricultura. 216 p.
Hopkins, M.S., Reddell, P., Hewett, R.K. & Graham, A.W. (1996). Comparison of Root and Mycorrhizal Characteristics in Primary and Secondary Rainforest on a Metamorphic Soil in North Queensland, Australia. Journal of Tropical Ecology, 12 (6), 871 - 885.
Hoshmand, A. R. (1998). Statistical Methods for Environmental & Agricultural Sciences. Second edition. New York: CRC Press LLC. 439 p.
IGAC. Instituto Geográfico Agustín Codazzi. (2002). Mapa de Suelos de Colombia. Recuperado de: http://mapascolombia.igac.gov.co/wps/portal/mapasdecolombia/.
Jackson, R. B., Mooney, H. A. & Schulze. E.D. (1997). A global budget for fine root biomass, surface area, and nutrient contents. Proceedings of the National Academy of Sciences of the United States of America, 94, 7362 - 7366.
James, J.J., Ziegenhagen, L. & Aanderud, Z. T. (2010). Exploitation of Nutrient-Rich Soil Patches by Invasive Annual and Native Perennial Grasses. Invasive Plant Science and Management, 3, 169 - 177.
Jaramillo, V.J., Ahedo-Hernández, R. & Kauffman, J. B. (2003). Root biomass and carbon in a tropical evergreen forest of Mexico: changes with secondary succession and forest conversion to pasture. Journal of Tropical Ecology, 19, 457 - 464.
Jiménez, E.L., Moreno-Hurtado, F., Peñuela, M.C., Lloyd, J., Patiño, S., & Álvarez. E. (2008). Estimación de la masa de raíces finas en dos bosques de tierra firme sobre suelos contrastantes en la Amazonia colombiana. Pp. 165 - 196. En: Buitrago, A.I., & Jiménez, E.M. (Eds.). Gente, Tierra y Agua: Imani Mundo III. Universidad Nacional de Colombia Sede Amazonia. Instituto Amazónico de Investigaciones IMANI. Bogotá: Editorial Guadalupe Ltda. 240 p.
Kochsiek, A., Tan, S. & Russo, S.E. (2013). Fine root dynamics in relation to nutrients in oligotrophic Bornean rain forest soils. Plant Ecology, 214(5), 1385 - 0237.
Kummerow, J., Castillanos, J. Maas, M. & Larigauderie, A. (1990). Production of fine roots and the seasonality of their growth in a Mexican deciduous dry forest. Vegetatio, 90, 73 - 80.
Malagon D., Pulido, C., Llinas, R. D., Chamorro, C. & Fernández, J. (1995). Suelos de Colombia. Origen, Evolución, Clasificación, Distribución y uso. Bogotá: Instituto Geográfico Agustín Codazzi. Subdirección de Agrología. 632 p.
Martínez, J. O. (1993). Geomorfología. En: Leyva, P. (Ed.). Colombia Pacífico, Tomo I. Fondo para la Protección del Medio Ambiente “José Celestino Mutis” FEN Colombia.
Maycock, C.R. & Congdon, R.A. (2000). Fine Root Biomass and Soil N and P in North Queensland Rain Forests. Biotropica, 32(1), 185 - 190.
Metcalfe, D. B., Meir, P., Aragao, L. E., Da Costa, A. C. L., Braga, A. P., Gonḉalves, P. H. L., De Athaydes Silva Jr., J., De Almeida, S. S., Dawson, L. A., Malhi, Y., & Williams, M. (2008). The effects of water availability on root growth and morphology in an Amazon rainforest. Plant Soil, 311, 189 - 199.
Moreno-Hurtado, F. (2004). Soil Carbon Dynamics in Primary and Secondary Tropical Forests in Colombia. Thesis of Doctor of Philosophy in Biology. Miami: Florida International University. 208 p.
Osorio, N. W. (2014). Manejo de nutrientes en suelos del Trópico. Segunda edición. Medellín: Editorial L. Vieco S.A.S. 416 p.
Ostertag, R. (2001). Effects of nitrogen and phosphorus availability on fine-root dynamics in Hawaiian montane forests. Ecology, 82(2), 485-499.
Poveda, I.C., Rojas, C., Rudas, A., & Rangel, O. (2004). El Chocó biogeográfico: Ambiente Físico. En Rangel, O. (2004). Colombia Diversidad Biótica IV. El Chocó biogeográfico/ Costa Pacífica. Instituto de Ciencias Naturales. Bogotá: Universidad Nacional de Colombia. 1024 p.
Powers, J. S., Treseder, K. K. & Lerdau, M. T. (2005). Fine roots, arbuscular mycorrhizal hyphae and soil nutrients in four neotropical rain forests: patterns across large geographic distances. New Phytologist, 165, 913 - 921.
Quinto, H., & Moreno, F. H. (2014). Diversidad florística arbórea y su relación con el suelo en un bosque pluvial tropical del Chocó Biogeográfico. Revista árvore, 38 (6), 1123 -1132.
R Development Core Team. (2012). R: A language and environment for statistical computing. Vienna, Austria. R Foundation for Statistical Computing. ISBN: 3-900051-07-0. Recuperado de http://www.r-project.org/.
Ruiz - Murcia, J. F. (2010). Cambio Climático en Temperatura, Precipitación y Humedad Relativa para Colombia Usando Modelos Meteorológicos de Alta Resolución (Panorama 2011-2100). Instituto de Hidrología, Meteorología y Estudios Ambientales - IDEAM. Subdirección de Meteorología. Nota Técnica IDEAM-METEO/005-2010. Bogotá, D. C., Mayo (2010). 91 p.
Sanford, R. L. & Cuevas, E. (1996). Root growth and rhizosphere interactions in tropical forests. En: Mulkey, S. S.; Chazdon, R. L and Smith, A. P. (Eds). Tropical forest plant ecophysiology. New York: Chapman and Hall. p. 268 - 300.
Saugier, B., Roy, J. & Mooney, H.A. (2001). Estimations of global terrestrial productivity: Converging toward a single number? in Roy, J., Saugier, B. & Mooney, H.A. (Eds). Terrestrial Global Productivity. San Diego, CA: Academic Press. 543-557.
Sayer, E. J., Wright, S. J., Tanner, E. V. J., Yavitt, J. B., Harms, K. E., Powers, J. S., Kaspari, M., Garcia, M. N., & Turner, B. L. (2012). Variable Responses of Lowland Tropical Forest Nutrient Status to Fertilization and Litter Manipulation. Ecosystems, 15, 387 - 400.
Silver, W. L., Thompson, A. W., Mcgroddy, M. E., Varner, R. K., Dias, J. D., Silva, H., Crill, P. M., & Keller, M. (2005). Fine root dynamics and trace gas fluxes in two lowland tropical forest soils. Global Change Biology, 11, 290 - 306.
Silver, W.L., Neff, J., McGroddy, M., Veldkamp, E., Keller, M. & Cosme, R. (2000). Effects of Soil Texture on Belowground Carbon and Nutrient Storage in a Lowland Amazonian Forest Ecosystem. Ecosystems, 3, 193 - 209.
Stark, N.M. & Jordan, C.F. (1978). Nutrient retention by the root mat of an Amazonian Rain Forest. Ecology, 59(3), 434 - 437.
Statistical Graphics Corp. (2002).StatgraphicsPlusCenturium Version 5.1. Recuperado de http://www.Statgraphics.com.
Trumbore, S., Da Costa, E. S., Nepstad, D. C., De Camargo, P. B., Martinelli, L., Ray, D., Restom, T., & Silver, W. (2006). Dynamics of fine root carbon in Amazonian tropical ecosystems and the contribution of roots to soil respiration, Global Change Biology, 12, 217 - 229.
Vitousek, P.M. (1984). Litterfall, Nutrient Cycling, and Nutrient Limitation in Tropical Forests. Ecology, 65 (1), 285 - 298.
Vitousek, P.M. & Sanford. R. L. (1986). Nutrient cycling in moist tropical forest. Annual Review of Ecology and Systematics, 17, 137 - 167.
Vogt, K.A., Grier, C.C. & Vogt, D.J. (1985). Production, turnover, and nutrient dynamics of above- and below-ground detritus of world forests. Advances in Ecological Research, 15, 303- 377.
West, R. (1957). Las tierras bajas del Pacífico colombiano. Instituto Colombiano de Antropología. Bogotá: Imprenta Nacional de Colombia. .300 pp.
Wright, S.J., Yavitt, J.B., Wurzburger, N., Turner, B. L., Tanner, E.V.J., Sayer, E.J., Santiago, L.S., Kaspari, M., Hedin, L.O., Harms, K.E., Garcia, M.N. & Corre, M.D. (2011). Potassium, phosphorus, or nitrogen limit root allocation, tree growth, or litter production in a lowland tropical forest. Ecology, 92(8), 1616 - 1625.




Licencia
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






