
DOI:
https://doi.org/10.14483/22487085.11315Published:
2017-08-04Issue:
Vol 19, No 2 (2017) July-DecemberSection:
Research ArticlesStrategies of metalinguistic and recast feedback during oral interactions
Estrategias de retroalimentación metalingüística durante las interacciones orales de estudiantes
Keywords:
retroalimentación correctiva, metalingüística, reformulación, respuesta del estudiante, interacciones orales (es).Keywords:
corrective feedback, metalinguistic, oral interactions, recast, student uptake (en).Downloads
References
Asari, Y. (2012). Types of recasts and learners’ uptake. Dialogue, 10, 1-20.
Chaudron, C. (1977). A descriptive model of discourse in the corrective treatment of learners’ errors. Language Learning, 27(1), 29-46.
Dörnyei, Z., & Skehan, P. (2003). Individual differences in second language learning. The handbook of second language acquisition. In C. J. Doughty & M. H. Long (Eds.), The handbook of second language
acquisition (pp. 589-630). Oxford, UK: Blackwell.
Ellis, R. (2006). Researching the effects of form-focused instruction on L2 acquisition. AILA Review, 19, 18-41.
Ferreira, A. (2007). A Study of feedback strategies in foreign language classroom and tutorials with implications for intelligent computer-assisted language learning systems. International Journal of Artificial Intelligence in Education, 17, 389-422.
Hendrickson, J. (1978). Error correction in foreign language teaching: Recent theory, research, and practice. Modern Language Journal, 62, 387-398.
Johnson, B. (2001). Toward a new classification of nonexperimental quantitative research. Educational Researcher, 30(2), 3-13.
Katayama, A. (2007). Learners’ perceptions toward oral error correction. In K. Bradford-Watts (Ed.), JALT2006 Conference Proceedings. Tokyo: JALT
Lewis, M. (2002). Giving feedback in language classes. Singapore: SEAMEO Regional Language Center.
Lightbown, P., & Spada, N. (1990). Focus-on-form and corrective feedback in communicative language teaching. Studies in Second Language Acquisition, 12(4), 429-448.
Long, M., & Robinson, P. (1998). Focus on form: Theory, research, and practice. In C. Doughty & J. Williams (Eds.), Focus on form in classroom second language acquisition (pp. 15-41). New York: Cambridge University.
Lyster, R., & Ranta, L. (1997). Corrective feedback and learner uptake: Negotiation of form in communicative classrooms. Studies in Second Language Acquisition, 19, 37-66.
Lyster, R., Saito, K., & Sato, M. (2013). Oral corrective feedback in second language classrooms. Language Teaching, 46(1), 1–40.
Nassaji, H. (2009). Effects of recasts and elicitations in dyadic interaction and the role of feedback explicitness. Language Learning, 59, 411-452.
Nassaji, H., & Fotos, S. (2011). Teaching grammar in second language classrooms: Integrating formfocused instruction in communicative contexts. New York, NY: Routledge.
Norris, J. (2000). Effectiveness of L2 instruction: A research synthesis and quantitative meta-analysis. Language Learning, 50, 417-528.
Panova, I., & Lyster, R. (2002). Patterns of corrective feedback and uptake in an adult ESL classroom. TESOL Quarterly, 36, 573-595.
Petchprasert, A. (2012). Feedback in second language teaching and learning. US-China Foreign Language, 10(4), 1112-1120.
Schmidt, R. (1990). The role of consciousness in second language learning. Applied Linguistics, 11(2). 129- 158.
Schmidt, R. (2001). Attention. In P. Robinson (Ed.), Cognition and second language instruction (pp. 3-32) Cambridge: Cambridge University Press.
Swain, M. (1985). Communicative competence: Some roles of comprehensible input and comprehensible output in its development. In S. Gass & C. Madden (Eds.), Input in second language acquisition (pp. 235– 253) Rowley, MA: Newbury House.
Swain, M. (1995). Three functions of output in second language learning. In G. Cook & B. Seidlhofer (Eds.), Principles and practice in applied linguistics (pp. 125–144). Oxford: Oxford University Press.
Westhoff, G. J. (2004). The art of playing a pinball machine. Characteristics of effective SLA-tasks. Babylonia, 12(3), 52–57.
Yang, Y., & Lyster, R. (2010). Effects of form-focused practice and feedback on Chinese EFL learners’ acquisition of regular and irregular past tense forms. Studies in Second Language Acquisition, 32, 235-
How to Cite
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Download Citation
RESEARCH ARTICLES
Strategies of Metalinguistic and Recast Feedback during Oral Interactions1
Estrategias de retroalimentación metalingüística durante las interacciones orales de estudiantes
Monica Raquel Tamayo2 Diego Cajas3
Citation/ Para citar este Artículo: Tamayo, M. & Cajas, D. (2017). Strategies of Metalinguistic and Recast Feedback on Learner Uptake during Oral Interactions in Upper-Intermediate Level Foreign Language Students. Colomb. appl linguist.]., 19(2), pp. 165-176.
Received: 05-Dec-2016 / Accepted: 05-May-2017 DOI: http://dx.doi.org/10.14483/22487085.11315
Abstract
Corrective feedback has attracted much attention in recent years, this with a particular emphasis on meaning-focused language instruction. In order to compare the effectiveness of the strategies of metalinguistic and recast feedback on student uptake during oral interactions, an eight-week non-experimental study was conducted. This study comprised thirty participants distributed in two classes. One group of 16 students was exposed to metalinguistic feedback and the other group of 14 students to recast. The selected students were aged from 18 to 20 year-old and were attending the eighth course of the English Foreign Language Program in a public university in Ecuador for one semester. To compare the effectiveness of the two aforementioned feedback strategies, four target structures were adopted: omission of subject, auxiliary use in questions, subject-verb agreement, and reported statements. The structures emerged from a survey which was administered to a sample of EFL teachers from the Language Center of the university. The findings of the study revealed that learners who were exposed to metalinguistic feedback outperformed their counterparts who were exposed to recast feedback.
Keywords: corrective feedback, metalinguistic, oral interactions, recast, student uptake
Resumen
La retroalimentación correctiva ha atraído mucha atención en los últimos años, con un énfasis particular en la enseñanza del lenguaje centrada en el significado. Con el fin de comparar la eficacia de las estrategias de retroalimentación metalingüística y de reformulación en la respuesta del estudiante durante las interacciones orales, se condujo un estudio cuasi experimental de ocho semanas. Este estudio comprendió treinta participantes distribuidos en dos clases. Un grupo de 16 estudiantes fue expuesto a la retroalimentación metalingüística y el otro grupo de 14 estudiantes a la de reformulación. Los estudiantes seleccionados tenían entre 18 y 20 años de edad y asistían al octavo curso del Programa de Inglés como lenguaje extranjera en una universidad pública en Ecuador durante un semestre.
Para comparar la efectividad de las dos estrategias de retroalimentación mencionadas anteriormente, se adoptaron cuatro estructuras: omisión del sujeto, uso del auxiliar en preguntas, relación sujeto-verbo, reporte de oraciones. Las estructuras seleccionadas surgieron de una encuesta que se administró a una muestra de profesores de Inglés del Centro de Idiomas de la universidad. Los hallazgos del estudio revelaron que los estudiantes que estaban expuestos a la retroalimentación metalingüística superaron a sus compañeros expuestos a la retroalimentación de reformulación.
Palabras clave: retroalimentación correctiva, metalingüística, reformulación, respuesta del estudiante, interacciones orales
Introduction
A sole exposure to a foreign or second language is not enough for students to acquire the target language elements with a high level of competence (Lightbown & Spada, 1990; Long & Robinson, 1998; Norris, 2000). It is necessary for students to interact with their teachers in order to receive proper feedback. Thus, one of the biggest challenges for teachers is to provide appropriate feedback which is less intimidating and more effective during communicative interactions. In this regard, two types of feedback have been the focus of research: corrective feedback and learner uptake. Corrective feedback provides important information that can be used by students in the modification of their errors. Learner uptake provides students’ responses that immediately follow the teacher’s feedback (Lyster & Ranta, 1997). At a classroom level, based on the literature, corrective feedback during language interactions seems to have a prominent role as it allows students to confirm, discard, or modify their hypothetical rules.
In this respect, one of the aims of this study is to identify the types of strategies that can best facilitate, through proper feedback, the development of English language skills. Although there are several investigations related to the type, frequency, and effectiveness of corrective feedback strategies, there is still a debate over what type of corrective feedback is more effective (Chaudron, 1977; Ellis, 2006; Ferreira, 2007). Thus, this research seeks to shed light on a deeper understanding of corrective feedback and learner uptake in order to help educators become more familiar with the type of feedback strategies that can help students develop their English. The second aim of this study is to compare and describe the effect of metalinguistic and recast feedback on learner uptake during oral interactions and its implications in ELT.
Theoretical Framework
The theoretical claim on which the development of corrective feedback and uptake have occurred is the output hypothesis proposed by Swain (1985). In her research in French immersion classrooms, she observed that students’ production was inaccurate. Therefore, Swain’s output hypothesis stated that by providing input alone, students do not improve their language acquisition and that the production of output in response to input is important for language development. However, input needs to be of quality in order to positively affect change in the output. Swain (1985) further mentioned that in order for modified output to occur, providing useful and consistent feedback is necessary. She further suggested that modified output is the description of “the leading edge of a learner’s interlanguage” (p. 248). In the context of teacher-learner interaction, modified output can be presented in the form of learner uptake, which is the student’s reaction to the teacher’s corrective feedback. Since students’ output can be influenced by the type of teacher feedback, it can therefore be considered a pedagogical tool which may pave the way for effective English learning.
Lyster and Ranta (1997) define uptake as “a reaction to the teacher’s intention to draw attention to some aspect of the student’s initial utterance” (p. 49). This refers to a student’s immediate response after receiving corrective feedback by the teacher. Uptake is considered successful when a student has understood the linguistic form and has corrected the error. On the other hand, unsuccessful uptake occurs when a student has not been able to reformulate and correct the error. Lyster and Ranta (1997) suggest that uptake that results in repair of the error can be considered evidence of learning. However, this may not be considered as a proof of language learning, yet may show that noticing has occurred which is an essential starting point for the production of students’ utterances. (Nassaji, & Fotos, 2011). Through noticing, learners are encouraged to produce output which is vital to developing accuracy (Swain, 1995). Thus, the role of the teacher in facilitating students’ language awareness is of paramount importance.
Learner uptake is worth considering in relation to the noticing hypothesis (Schmidt, 1990). Uptake may be pointed out as one form of manifestation of attention. Since no explicit research has demonstrated that uptake is an oral manifestation of noticing, it may be suggested that there are some linguistic features that are noticed by students when uptake is produced. This awareness has a positive effect on students’ language acquisition since by noticing and reflecting on their mistakes, errors are minimized and error fossilization is prevented.
Lyster, Saito, and Sato (2013) consider corrective feedback as “responses to learner utterance containing an error” (p. 1). Teachers’ corrective feedback is a positive way to improve students’ language learning by drawing their attention to linguistic forms produced incorrectly. Corrective feedback provides information that students can actively use in modifying the errors of their production. The feedback strategies allow students to confirm, discard, and possibly modify their own grammar rules with the purpose of increasing learning. Feedback can be considered as a pedagogical tool that offers modified input to students who in turn would produce modified output. Lewis (2002) goes further by highlighting four aspects regarding oral feedback: (1) it supports students in acquiring language input as they learn vocabulary and structures in context; (2) it provides information for both teachers and students as it describes the way students learn, and serves as a kind of assessment; (3) it is a way of motivating students to improve their performance, and (4) it is one step forward towards students’ self- reliance, as they start identifying and correcting their own mistakes.
When the corrective feedback occurs in response to errors that arise naturally in foreign language classes, it gives students a sense of accomplishment and motivates them to learn more (Katayama, 2007; Petchprasert, 2012). To provide adequate feedback, the student knows that when exposed to the possibility of making mistakes, he or she is giving the opportunity to improve and overcome his or her insecurity. It is a way of motivating students to improve their performance as they start identifying and correcting their own errors. Westhoff (2004) states that if a language learner is aware of language learning strategies, the learning process will improve much faster. Feedback can be given to each student, a group of them, or the whole class.
Lyster and Ranta (1997) have provided the most complete taxonomy of corrective feedback. They classified corrective feedback into six categories which include: explicit correction, recast, metalinguistic feedback, elicitation, repetition, and clarification request. To these ends, they suggest that “recast is one type of feedback that refers to the teacher’s reformulation of all or part of a student’s utterance, minus the error” (p. 46). This reformulation can also be considered as “paraphrase.” This paraphrasing indicates that the student’s statement was incorrect avoiding referring to that specific error in particular. However, “some recasts are more noticeable than others since they may focus on one word only, whereas other recasts include modification in grammatical or lexical forms. Recasts also involve translations in response to a student’s use of the L1” (Lyster & Ranta, 1997, p. 47). Thus, the teacher may respond to an error by using more than one type of recast.
Another type of feedback is called metalinguistic feedback. Lyster and Ranta (1997) referred to this type of feedback as “comments, information, or questions related to the well-formedness of the student’s utterance, without explicitly providing the correct form” (p. 47). Metalinguistic information can provide the learners with a variety of hints related to possible reformulations of the target forms that contain errors. (e.g., “It’s plural”) or a word definition in the case of lexical errors (Lyster & Ranta, 1997).
Some studies that have examined the effectiveness of metalinguistic and recast feedback show that first feedback produced better results because it assisted learners in the process of language learning. For instance, the study carried out in a Chinese EFL setting by Yang & Lyster (2010) reported that by providing metalinguistic feedback, learners increased their accuracy in the use of regular past tense. Additional supportive evidence is that of Lyster and Ranta (1997). The results of their study of students of “French immersion” courses demonstrated that metalinguistic feedback led to learner uptake in 90%, whereas 31% was obtained after the provision of recast feedback. These results also seem to suggest that recasts are not very effective in eliciting learners’ uptake.
Methodology
This study employs a quantitative nonexperimental longitudinal explanatory research methodology. Johnson (2001) highlights one of the advantages of explanatory research when researchers attempt “to explain how the phenomenon operates by identifying the causal factors that produce change in it” (p. 9). It is also longitudinal because the data was collected at different moments during a specific period of time and non-experimental since there was not a random selection of participant students.
The focus of this study is to identify whether there is a relationship between the kind of teacher feedback and student uptake. In order to identify the difference between the kind of teacher feedback and student uptake, two independent variables were identified, for instance: recast and metalinguistic feedback. This was complemented with four dependent variables: omission of subject, auxiliary use in questions, subject-verb agreement, and reported statements. Thirty students attending their last mandatory EFL course in the language center of a public university were chosen as participants during the semester from October 2015 to February 2016. Since these students comprised two different EFL classes, it was not necessary to divide them into two groups. Instead, these classes as a whole constituted the two groups that comprised this study. Students in Group 1 (n = 16) were exposed to metalinguistic feedback strategy and Group 2 (n = 14) to recast feedback. The type of feedback to be used in each group was randomly done. For the random selection of the type of feedback, two students (one per each group) were asked to choose one closed envelop from a group of two. Each envelop contained a card indicating the type of feedback, for instance one envelop contained recast and the other metalinguistic feedback. Hence, the type of feedback selected by the two students was used throughout the study. Students in these two groups were aged 18 to 20 years old. The participants had been learning English for one and a half years. They had an intermediate level of English which allowed them to participate in the task-based interaction used as part of the treatment in this study. All participants had a B1 level of English according to the Common European Framework of Reference. This was identified through the final EFL exam which all participants completed to graduate from seventh to eighth course of English. Participants’ written consent was obtained previous the beginning of this research. Finally, the class teacher agreed that the researcher would be the one who conducted all the activities. A written consent was also provided to the researcher in order to carry out this study with the two groups.
Regarding the grammar structures used in the feedback activities, these emerged as the result of the administration of a survey completed by 15 teachers of the eighth EFL course in the language center of the selected university. All teachers were asked one open-ended question. This question was: “based on your experience, what grammar structures do you think are the most difficult for students to acquire?” In order to proceed with the subsequent selection of the grammar structures, the top four most recurrent teachers’ answers were selected. The results were as follows: 100% of teachers mentioned that auxiliary use in questions was the most difficult grammar structure, followed by the omission of subject and reported statements both representing the 70% of teachers’ answers. Finally, subject-verb agreement scored fourth with 68%. Furthermore, students’ knowledge of these structures was diagnosed via a multiple choice test. This test was designed by the researchers and piloted in a different class to check its reliability. Once it was piloted, this test was conducted to the two groups. The scores obtained by the students in this diagnostic test were collected in a record sheet which confirmed that the students did have difficulty in the structures mentioned by their teachers.
For purposes of this study and application of treatments, 18 sessions (nine sessions per group) were planned. Each session lasted 120 minutes during a period of eight weeks. The first group was exposed to recast feedback for nine sessions, and the second group to metalinguistic feedback for nine sessions.
In order to measure correct language repairs, students were first engaged in the following activities: (1) comments to five videos of approximately five minutes each, and (2) comments on four short readings. These activities were formulated with the objective of facilitating English interactions among students and to provide them with enough English language input through videos and reading activities. The data for analysis included the interactions that students did with the teacher.
The procedure in each activity was as follows: First, the theme was introduced in the same way it is usually done in a normal class. Second, students were introduced to focused tasks which targeted the use of the four grammatical structures. Third, instructions were given to students regarding the tasks they had to perform. Useful words were reviewed but no new structures were taught. This stage was for preparation prior to the task cycle, which consisted of three elements:
(A) Task: This was done by students individually and in pairs. The researcher supervised the students, but did not intervene to correct errors.
(B) Preparation: Students had to watch the video of approximately 5-6 minutes or read the chosen text, individually prepare a one-minute oral summary to present it, have four questions ready to ask his or her partner about the video or text, and be prepared to report four statements said by his or her partner.
(C) Report: Each student had to interact with a partner selected by the researcher at least four times in each task. The total number of interventions per student was 16. During the intervention of the students, the researcher provided either recast or metalinguistic feedback that corresponded to each group. The feedback was provided to each pair without the presence of the other students.
A record sheet was used to collect data in order to register the type of student response (uptake) to the corresponding feedback. Based on the on the existing categories of uptake sequences proposed by Lyster and Ranta, (1997), uptake was then coded as either “repair” or “needs repair.” For this study, the following verbal responses were considered “uptake” as presented in the following table:
The frequency and percentage of students’ error repairs were calculated with the values obtained. The values were expressed numerically and graphically. This helped determine the feedback treatment that led students to have the most successful repairs. A
Note. Adapted from Asari (2012).
final survey was given to students who participated in the study. This helped analyze the degree of acceptance of the feedback received.
Analysis and Discussion of Results
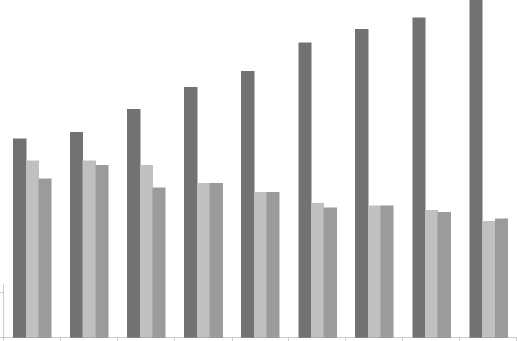
When the metalinguistic feedback strategy was used with Group 1, the number of error repairs steadily increased during the nine sessions. The number of error repairs improved from 89 in session one to 151 in session nine. On the other hand, students’ uptake that led to no repair was identified. The students modified errors but did not repair, decreasing from 79 in session one to 52 in session nine. Likewise, the students repeated the errors or left them unmodified without repair during the nine sessions (71 times in the first session, to 53 times in the last one; see Figure 1). This reduction in both modified and unmodified uptakes is related to the increase in repairs.
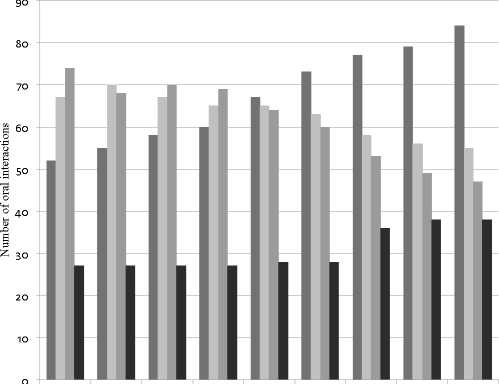
When the recast feedback strategy was applied to Group 2, the number of error repairs slightly increased during the nine sessions. The number of error repairs improved from 52 in session one to 84 in session nine. On the other hand, the number of errors that the students modified but did not repair as well as the ones that the students repeated or unmodified was higher than the number of repairs until session four. It is from session five that the number of repairs exceeds the number of no repairs. When evaluating recast feedback, acknowledgement, which is another type of uptake, was also registered. Acknowledgement was stable (27-28) from the first to the sixth session, and was increasing from the seventh session to the last one (36-38; Figure 2). This student uptake may have occurred due to the repeatability of the strategy.
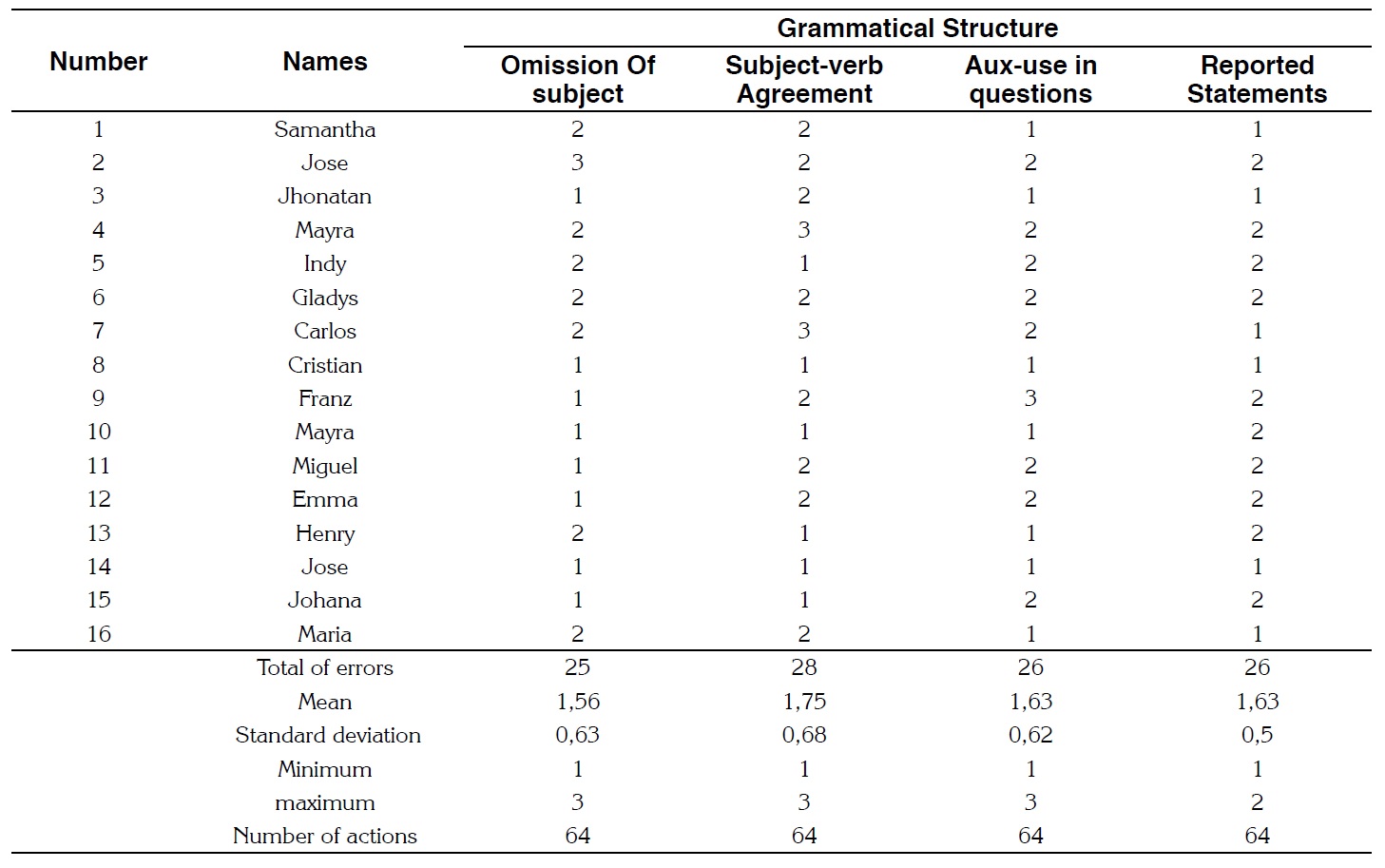
With the metalinguistic feedback strategy in Group 1, 105 errors were committed in the last session (Table 2). This implies that 41% of the errors were not repaired from the total of 256 possible interactions. To exemplify how students repaired their errors when using metalinguistic feedback, the following quote includes an interaction between a student (S) and the researcher (R) testing a subject verb agreement:
0
40
140
120
100
80
60
20
■ Repair
■ Modified
■ Unmodified
Five
Session
Eight
Three
One
Two
Four
Six
Seven
Nine
Fig 1. Effect of metalinguistic feedback strategy on student uptake (error repair, modified and unmodified error through nine intervention sessions)
■ Repair
■ Modified
■ Unmodified
■ Acknowledgement
One Two Three Four Five Six Seven Eight Nine
Session
Fig 2. Effect of recast feedback strategy on student uptake (error repair, modified,unmodified error and acknowledgement) through nine intervention sessions.
S: The last two questions was especially hard.
R: Is the word “questions” singular or plural?
S: It’s plural, ok then... were especially hard
Here, the researcher promotes self-reflection of the error produced by the student by emphasizing the grammatical number through the word ‘questions’ drawing the student’s attention to the noun. The student noticed the correct use of the verb to be (subject verb agreement). For further statistical analysis of the results of metalinguistic feedback, the mean of non-repaired errors of the four grammatical structures (omission of subject, subject-verb agreement, auxiliary use in questions, and reported statements) collected in session nine using this feedback strategy (Table 2) was lower than the mean of errors registered in the diagnostic test on the aforementioned structures. Additionally for testing the statistical significance of this study an independent sample T-test was calculated. The results of this test demonstrated statistical significance for the first session (p = .001) as well as from the second to the ninth session (p = .000). Considering that values which are lower than 0.05 are statistically significant; for this study, the null hypothesis was rejected. This means that the students who received a metalinguistic feedback repair more errors that students who received a recast feedback.
The standard deviation of non-repair errors of the four grammatical structures in session nine using metalinguistic feedback strategy (Table 2) was lower in comparison with the standard deviation of errors registered in the diagnostic test.
With the recast feedback strategy in Group 2, 140 errors were committed in the last session (Table 3). The interaction occurred between a student (S) and the researcher (R) are encapsulated in the following interaction:
S: The girl in the park watched the movie twice.
R: The girl in the park has watched the movie twice.
S. Yes, the girl has watched the movie twice.
The above excerpt shows how the researcher uses a recast feedback with the student. In this interaction, the researcher does not use extra clues to raise
Grammatical Structure
|
Number |
Names |
Omission Of subject |
Subject-verb Agreement |
Aux-use in questions |
Reported Statements |
|
1 |
Samantha |
2 |
2 |
1 |
1 |
|
2 |
Jose |
3 |
2 |
2 |
2 |
|
3 |
Jhonatan |
1 |
2 |
1 |
1 |
|
4 |
Mayra |
2 |
3 |
2 |
2 |
|
5 |
Indy |
2 |
1 |
2 |
2 |
|
6 |
Gladys |
2 |
2 |
2 |
2 |
|
7 |
Carlos |
2 |
3 |
2 |
1 |
|
8 |
Cristian |
1 |
1 |
1 |
1 |
|
9 |
Franz |
1 |
2 |
3 |
2 |
|
10 |
Mayra |
1 |
1 |
1 |
2 |
|
11 |
Miguel |
1 |
2 |
2 |
2 |
|
12 |
Emma |
1 |
2 |
2 |
2 |
|
13 |
Henry |
2 |
1 |
1 |
2 |
|
14 |
Jose |
1 |
1 |
1 |
1 |
|
15 |
Johana |
1 |
1 |
2 |
2 |
|
16 |
Maria |
2 |
2 |
1 |
1 |
|
Total of errors |
25 |
28 |
26 |
26 | |
|
Mean |
1,56 |
1,75 |
1,63 |
1,63 | |
|
Standard deviation |
0,63 |
0,68 |
0,62 |
0,5 | |
|
Minimum |
1 |
1 |
1 |
1 | |
|
maximum |
3 |
3 |
3 |
2 | |
|
Number of actions |
64 |
64 |
64 |
64 |
the student’s language awareness. The researcher limits her feedback to the repetition of the structure using the correct form of subject verb agreement and emphasis on the correct structure is done. This emphasis is expressed in bold (has). Data regarding this kind of feedback manifested that 63% of the errors were not repaired from the total of 224 possible interactions. The mean of non-repaired errors of the four grammatical structures (omission of subject, subject-verb agreement, auxiliary use in questions, and reported statements) collected in session nine using this feedback strategy (Table 3) was lower than the mean of errors registered in the diagnostic test on the aforementioned structures. The standard deviation of non-repair errors for the four grammatical structures in session nine using recast feedback (Table 3) was lower in comparison with the standard deviation of error registered in the diagnostic test.
The two feedback strategies result in a reduction of errors during oral interactions. Fifty-nine percent
of grammatical errors were repaired by using metalinguistic feedback in comparison with 38% of errors repaired by using recast feedback. Therefore, metalinguistic feedback, based on the data, proved to be more effective than recast.
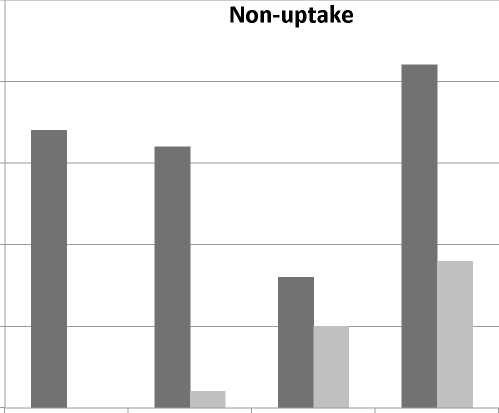
Twenty-one non-uptakes were registered during interactions using metalinguistic feedback in contrast to nine no-uptakes during interactions using recast feedback. Perhaps this occurred because the metalinguistic strategy elicited answers from students instead of providing them, so some students decided not to respond (Figure 3).
Survey Results
An acceptance survey was applied to the two groups. The results of the survey to the two types of feedback strategies of metalinguistic and recast were 90% of the students who thought that receiving
Grammatical Structure
25
20
15
10
5
0
■ Metalinguistic
■ Recast
OMISSION OF SUBJECT VERB AUX-USE IN REPORTED
SUBJECT AGREEMENT QUESTIONS STATEMENTS
Target structure
Table 3. Number of non-repairs registered in the last session with recast feedback Strategy
|
Number |
Names |
Omission Of subject |
Subject-verb Agreement |
Aux-use in questions |
Reported Statements |
|
1 |
Elizabeth |
3 |
2 |
2 |
2 |
|
2 |
Luis |
3 |
3 |
2 |
2 |
|
3 |
Alexander |
2 |
3 |
3 |
2 |
|
4 |
Alejandra |
2 |
3 |
3 |
2 |
|
5 |
Natan |
3 |
2 |
3 |
3 |
|
6 |
Veronica |
2 |
3 |
2 |
3 |
|
7 |
Eduardo |
2 |
3 |
2 |
3 |
|
8 |
Michael |
3 |
3 |
2 |
1 |
|
9 |
Francois |
3 |
2 |
2 |
3 |
|
10 |
Lucia |
3 |
3 |
2 |
3 |
|
11 |
Joel |
2 |
3 |
2 |
2 |
|
12 |
David |
3 |
2 |
2 |
3 |
|
13 |
Iveth |
2 |
4 |
3 |
2 |
|
14 |
Yolanda |
3 |
3 |
2 |
2 |
|
Total of errors |
36 |
39 |
32 |
33 | |
|
Mean |
2,57 |
2,79 |
2,29 |
2,36 | |
|
Standard deviation |
0,51 |
0,58 |
0,47 |
0,63 | |
|
Minimum |
2 |
2 |
2 |
1 | |
|
maximum |
3 |
4 |
3 |
3 | |
|
Number of actions |
56 |
56 |
56 |
56 |
Figure 3. Number of non-uptake during metalinguistic and recast feedback sessions
feedback was interesting and that contributed to improving their oral production. Seventy-nine percent of the students graded the degree of contribution of the feedback strategies to their oral production as excellent, 13% as very good, and 8% as good. This suggests that students who participated in this study are open to receiving feedback from teachers.
This study was designed to compare the effects of two different feedback strategies on learner uptake during oral interactions. The result of this study showed that students repaired more errors after being exposed to feedback strategies. This indicates that exposing students to the target structures in combination with consciousnessraising activities such as error correction can improve their production. This finding is supported by Swain (1995) who claimed that corrective feedback assists students to test their hypotheses about the target language and replace them with the correct ones. This is also in line with Schmidt’s (1990) noticing hypothesis which highlights the role of feedback in drawing students’ attention to the target structures and comparing with their own forms, so as to correct their errors.
The analysis of data showed that metalinguistic feedback led to higher uptake results than recasts (Tables 2 & 3), since the participants exposed to metalinguistic corrective feedback outperformed those receiving recasts during communicative interactions. One explanation can be that metalinguistic feedback is “output-pushing” as suggested by Yang and Lyster (2010). While producing output, learners in this study faced some difficulties in conveying their meaning, but by exposing them to metalinguistic feedback, they were pushed to analyze and modify their output (Figure 1). This is in line with Swain (1995) who suggested that output practice can facilitate acquisition because it permits the development of cognitive processes, such as noticing, hypothesis testing, and metalinguistic reflection.
The findings in the current study also lend support to Schmidt’s (2001) hypothesis which emphasizes the importance of drawing learners’ attention to formal aspects of language for accomplishing linguistic development. In this case, metalinguistic feedback stimulates learners to openly identify the gap between their interlanguage and the target forms. Similarly, Hendrickson (1978) emphasized that students’ interlanguage can be developed by motivating them to produce more comprehensible and appropriate output rather than simply telling them the correct target forms. .
One justification behind the reduced effectiveness of recast in this study can be the fact that this group of learners is accustomed to selfproduction without feedback since the emphasis at the institution’s language center has always been on meaning rather than form. Therefore, students may have ignored the teacher’s reformulation as corrective feedback assuming it was mere positive evidence for their output. Research by Panova and Lyster (2002) seems to support the claim that recasts can be ambiguous to foreign language learners as they may tend to repeat mechanically what the teacher says without any significant learning. In their study, Panova and Lyster (2002) reported that metalinguistic prompts led students to develop better formulated utterances than those from recast. In this case, recasts appeared not to be very effective because learners, contrary to metalinguistic feedback, were just exposed to the correct form of target structures as modified input, without noticing the important role of output; that is, forcing learners to correct their interlanguage and elicit their self-repair. Metalinguistic feedback seems to be particularly helpful to these participating groups during the process of learning of specific target forms. The guidance provided by the teacher through this type of feedback can be a worthwhile investment of time and effort for teachers and students.
As far as non-uptake is concerned, certain students did not provide any verbal response immediately following teacher feedback. Chaudron (1977) viewed uptake as a main immediate measurement of the effectiveness of corrective feedback. When uptake with repair is produced, it is an indication that learner’s attention to the correct forms has been drawn and that learners have taken a step toward learning, while the lack of uptake can show that learners did not notice the mismatch between their statements and the target ones after the provision of feedback. This belief is opposed to Nassaji (2009) who mentioned that the absence of uptake is not an evidence of no noticing which can be assumed that the provision of feedback is not effective. It is possible that learners in this study have processed and learned from the feedback in their inner thought, but did not want to respond to it. Contextual variables may also have been intertwined in the uptake moves. For example, attitudes, concepts about learning, and motivation can influence how learners actually approach tasks and respond to feedback. The existence of those connections is acknowledged by Dornyei and Skehan (2003) who mention that “there are individual differences between learners in noticing abilities: other things being equal, some learners may be more likely to notice relevant qualities of input than others” (p. 597).
The acceptance survey showed that learners thought feedback was interesting and did not feel uncomfortable when receiving feedback. Some learners’ comments mentioned that they were receptive to feedback and were more willing to incorporate it because they perceived the correction as a manifestation of lending a helping hand, rather than a display of mockery, and the feedback was done individually and not shared in a group setting.
Conclusions
The findings of this study highlight the importance of corrective feedback in the process of English learning. In this regard, teachers can respond to students’ errors through two kinds of feedback, both metalinguistic and recast. Concerning these two types of feedback, it emerged from the data that metalinguistic feedback is more effective to students rather than recast. Its effectiveness may be correlated to students’ selfrealization of their mistakes facilitated through “comments, information, or questions related to the well-formedness of the student’s utterance, without explicitly providing the correct form” (Lyster & Ranta, 1997, p. 47). Teachers, therefore, need to become active key players in providing proper clues to improve students’ language awareness and subsequent reduction of errors. In this study, the data manifested that more exposure to metalinguistic feedback results in better error correction.
Metalinguistic feedback proves to be more effective because it encourages students more actively to self-repair, contrary to recast, when the correct form is provided to the students. This was reaffirmed by the data that the highest rate of uptake and successful repairs resulted from metalinguistic feedback, whereas recast feedback led to the lowest rate of uptake and successful repairs.
Complementary to the results, a survey of students who participated in the study was applied in order to determine the level of acceptance of the feedback received. The survey results manifested a positive opinion towards metalinguistic feedback and added that this feedback has contributed to the improvement of their oral production.
Comparing the effects of these two feedback strategies provided some light on theoretical issues such as the important contributions of corrective feedback in the EFL classroom and the cognitive role of noticing during the process of feedback. The results of this study also provide teachers with pedagogical alternatives to maximize the effect of feedback in their classes.
References
Asari, Y (2012). Types of recasts and learners’ uptake. Dialogue, 10, 1-20.
Chaudron, C. (1977). A descriptive model of discourse in the corrective treatment of learners’ errors. Language Learning, 27(1), 29-46.
Dornyei, Z., & Skehan, P (2003). Individual differences in second language learning. The handbook of second language acquisition. In C. J. Doughty & M. H. Long (Eds.), The handbook of second language acquisition (pp. 589-630). Oxford, UK: Blackwell.
Ellis, R. (2006). Researching the effects of form-focused instruction on L2 acquisition. A1LA Review, 19, 1841.
Ferreira, A. (2007). A Study of feedback strategies in foreign language classroom and tutorials with implications for intelligent computer-assisted language learning systems. International Journal of Artificial Intelligence in Education, 17, 389-422.
Hendrickson, J. (1978). Error correction in foreign language teaching: Recent theory, research, and practice. Modern Language Journal, 62, 387-398.
Johnson, B. (2001). Toward a new classification of nonexperimental quantitative research. Educational Researcher, 30(2), 3-13.
Katayama, A. (2007). Learners’ perceptions toward oral error correction. In K. Bradford-Watts (Ed.), JALT2006 Conference Proceedings. Tokyo: JALT
Lewis, M. (2002). Giving feedback in language classes. Singapore: SEAMEO Regional Language Center.
Lightbown, P, & Spada, N. (1990). Focus-on-form and corrective feedback in communicative language teaching. Studies in Second Language Acquisition, 12(4), 429-448.
Long, M., & Robinson, P (1998). Focus on form: Theory, research, and practice. In C. Doughty & J. Williams (Eds.), Focus on form in classroom second language acquisition (pp. 15-41). New York: Cambridge University.
Lyster, R., & Ranta, L. (1997). Corrective feedback and learner uptake: Negotiation of form in communicative classrooms. Studies in Second Language Acquisition, 19, 37-66.
Lyster, R., Saito, K., & Sato, M. (2013). Oral corrective feedback in second language classrooms. Language Teaching, 46(1), 1-40.
Nassaji, H. (2009). Effects of recasts and elicitations in dyadic interaction and the role of feedback explicitness. Language Learning, 59, 411-452.
Nassaji, H., & Fotos, S. (2011). Teaching grammar in second language classrooms: Integrating form-focused instruction in communicative contexts. New York, NY: Routledge.
Norris, J. (2000). Effectiveness of L2 instruction: A research synthesis and quantitative meta-analysis. Language Learning, 50, 417-528.
Panova, I., & Lyster, R. (2002). Patterns of corrective feedback and uptake in an adult ESL classroom. TESOL Quarterly, 36, 573-595.
Petchprasert, A. (2012). Feedback in second language teaching and learning. US-China Foreign Language, 10(4), 1112-1120.
Schmidt, R. (1990). The role of consciousness in second language learning. Applied Linguistics, 11 (2). 129158.
Schmidt, R. (2001). Attention. In P Robinson (Ed.), Cognition and second language instruction (pp. 3-32) Cambridge: Cambridge University Press.
Swain, M. (1985). Communicative competence: Some roles of comprehensible input and comprehensible output in its development. In S. Gass & C. Madden (Eds.), Input in second language acquisition (pp. 235- 253) Rowley, MA: Newbury House.
Swain, M. (1995). Three functions of output in second language learning. In G. Cook & B. Seidlhofer (Eds.), Principles and practice in applied linguistics (pp. 125-144). Oxford: Oxford University Press.
Westhoff, G. J. (2004). The art of playing a pinball machine. Characteristics of effective SLA-tasks. Babylonia, 12(3), 52-57.
Yang, Y, & Lyster, R. (2010). Effects of form-focused practice and feedback on Chinese EFL learners’ acquisition of regular and irregular past tense forms. Studies in Second Language Acquisition, 32, 235263.
This research project was a part of the activities we have to do as teachers/researchers of the Human Sciences Department from Universidad de las Fuerzas Armadas ESPE and Universidad Nacional de Educación, Ecuador. The department provided all the facilities to carry out this project.
Universidad de las Fuerzas Armadas ESPE, Sangolquí, Ecuador. mrtamayo@espe.edu.ec
Universidad Nacional de Educación, Azogues, Ecuador. diego.cajas@unae.edu.ec
Recibido: 5 de diciembre de 2016; Aceptado: 5 de mayo de 2017
Resumen
La retroalimentación correctiva ha atraído mucha atención en los últimos años, con un énfasis particular en la enseñanza del lenguaje centrada en el significado. Con el fin de comparar la eficacia de las estrategias de retroalimentación metalingüística y de reformulación en la respuesta del estudiante durante las interacciones orales, se condujo un estudio cuasi experimental de ocho semanas. Este estudio comprendió treinta participantes distribuidos en dos clases. Un grupo de 16 estudiantes fue expuesto a la retroalimentación metalingüística y el otro grupo de 14 estudiantes a la de reformulación. Los estudiantes seleccionados tenían entre 18 y 20 años de edad y asistían al octavo curso del Programa de Inglés como lenguaje extranjera en una universidad pública en Ecuador durante un semestre. Para comparar la efectividad de las dos estrategias de retroalimentación mencionadas anteriormente, se adoptaron cuatro estructuras: omisión del sujeto, uso del auxiliar en preguntas, relación sujeto-verbo, reporte de oraciones. Las estructuras seleccionadas surgieron de una encuesta que se administró a una muestra de profesores de Inglés del Centro de Idiomas de la universidad. Los hallazgos del estudio revelaron que los estudiantes que estaban expuestos a la retroalimentación metalingüística superaron a sus compañeros expuestos a la retroalimentación de reformulación.
Palabras clave
retroalimentación correctiva, metalingüística, reformulación, respuesta del estudiante, interacciones orales.Abstract
Corrective feedback has attracted much attention in recent years, this with a particular emphasis on meaning-focused language instruction. In order to compare the effectiveness of the strategies of metalinguistic and recast feedback on student uptake during oral interactions, an eight-week non-experimental study was conducted. This study comprised thirty participants distributed in two classes. One group of 16 students was exposed to metalinguistic feedback and the other group of 14 students to recast. The selected students were aged from 18 to 20 year-old and were attending the eighth course of the English Foreign Language Program in a public university in Ecuador for one semester. To compare the effectiveness of the two aforementioned feedback strategies, four target structures were adopted: omission of subject, auxiliary use in questions, subject-verb agreement, and reported statements. The structures emerged from a survey which was administered to a sample of EFL teachers from the Language Center of the university. The findings of the study revealed that learners who were exposed to metalinguistic feedback outperformed their counterparts who were exposed to recast feedback.
Keywords
corrective feedback, metalinguistic, oral interactions, recast, student uptake.Introduction
A sole exposure to a foreign or second language is not enough for students to acquire the target language elements with a high level of competence (Lightbown & Spada, 1990; Long & Robinson, 1998; Norris, 2000). It is necessary for students to interact with their teachers in order to receive proper feedback. Thus, one of the biggest challenges for teachers is to provide appropriate feedback which is less intimidating and more effective during communicative interactions. In this regard, two types of feedback have been the focus of research: corrective feedback and learner uptake. Corrective feedback provides important information that can be used by students in the modification of their errors. Learner uptake provides students’ responses that immediately follow the teacher’s feedback (Lyster & Ranta, 1997). At a classroom level, based on the literature, corrective feedback during language interactions seems to have a prominent role as it allows students to confirm, discard, or modify their hypothetical rules.
In this respect, one of the aims of this study is to identify the types of strategies that can best facilitate, through proper feedback, the development of English language skills. Although there are several investigations related to the type, frequency, and effectiveness of corrective feedback strategies, there is still a debate over what type of corrective feedback is more effective (Chaudron, 1977; Ellis, 2006; Ferreira, 2007). Thus, this research seeks to shed light on a deeper understanding of corrective feedback and learner uptake in order to help educators become more familiar with the type of feedback strategies that can help students develop their English. The second aim of this study is to compare and describe the effect of metalinguistic and recast feedback on learner uptake during oral interactions and its implications in ELT.
Theoretical Framework
The theoretical claim on which the development of corrective feedback and uptake have occurred is the output hypothesis proposed by Swain (1985). In her research in French immersion classrooms, she observed that students’ production was inaccurate. Therefore, Swain’s output hypothesis stated that by providing input alone, students do not improve their language acquisition and that the production of output in response to input is important for language development. However, input needs to be of quality in order to positively affect change in the output. Swain (1985) further mentioned that in order for modified output to occur, providing useful and consistent feedback is necessary. She further suggested that modified output is the description of “the leading edge of a learner’s interlanguage” (p. 248). In the context of teacher-learner interaction, modified output can be presented in the form of learner uptake, which is the student’s reaction to the teacher’s corrective feedback. Since students’ output can be influenced by the type of teacher feedback, it can therefore be considered a pedagogical tool which may pave the way for effective English learning.
Lyster and Ranta (1997) define uptake as “a reaction to the teacher’s intention to draw attention to some aspect of the student’s initial utterance” (p. 49). This refers to a student’s immediate response after receiving corrective feedback by the teacher. Uptake is considered successful when a student has understood the linguistic form and has corrected the error. On the other hand, unsuccessful uptake occurs when a student has not been able to reformulate and correct the error. Lyster and Ranta (1997) suggest that uptake that results in repair of the error can be considered evidence of learning. However, this may not be considered as a proof of language learning, yet may show that noticing has occurred which is an essential starting point for the production of students’ utterances. (Nassaji, & Fotos, 2011). Through noticing, learners are encouraged to produce output which is vital to developing accuracy (Swain, 1995). Thus, the role of the teacher in facilitating students’ language awareness is of paramount importance.
Learner uptake is worth considering in relation to the noticing hypothesis (Schmidt, 1990). Uptake may be pointed out as one form of manifestation of attention. Since no explicit research has demonstrated that uptake is an oral manifestation of noticing, it may be suggested that there are some linguistic features that are noticed by students when uptake is produced. This awareness has a positive effect on students’ language acquisition since by noticing and reflecting on their mistakes, errors are minimized and error fossilization is prevented.
Lyster, Saito, and Sato (2013) consider corrective feedback as “responses to learner utterance containing an error” (p. 1). Teachers’ corrective feedback is a positive way to improve students’ language learning by drawing their attention to linguistic forms produced incorrectly. Corrective feedback provides information that students can actively use in modifying the errors of their production. The feedback strategies allow students to confirm, discard, and possibly modify their own grammar rules with the purpose of increasing learning. Feedback can be considered as a pedagogical tool that offers modified input to students who in turn would produce modified output. Lewis (2002) goes further by highlighting four aspects regarding oral feedback: (1) it supports students in acquiring language input as they learn vocabulary and structures in context; (2) it provides information for both teachers and students as it describes the way students learn, and serves as a kind of assessment; (3) it is a way of motivating students to improve their performance, and (4) it is one step forward towards students’ self- reliance, as they start identifying and correcting their own mistakes.
When the corrective feedback occurs in response to errors that arise naturally in foreign language classes, it gives students a sense of accomplishment and motivates them to learn more (Katayama, 2007; Petchprasert, 2012). To provide adequate feedback, the student knows that when exposed to the possibility of making mistakes, he or she is giving the opportunity to improve and overcome his or her insecurity. It is a way of motivating students to improve their performance as they start identifying and correcting their own errors. Westhoff (2004) states that if a language learner is aware of language learning strategies, the learning process will improve much faster. Feedback can be given to each student, a group of them, or the whole class.
Lyster and Ranta (1997) have provided the most complete taxonomy of corrective feedback. They classified corrective feedback into six categories which include: explicit correction, recast, metalinguistic feedback, elicitation, repetition, and clarification request. To these ends, they suggest that “recast is one type of feedback that refers to the teacher’s reformulation of all or part of a student’s utterance, minus the error” (p. 46). This reformulation can also be considered as “paraphrase.” This paraphrasing indicates that the student’s statement was incorrect avoiding referring to that specific error in particular. However, “some recasts are more noticeable than others since they may focus on one word only, whereas other recasts include modification in grammatical or lexical forms. Recasts also involve translations in response to a student’s use of the L1” (Lyster & Ranta, 1997, p. 47). Thus, the teacher may respond to an error by using more than one type of recast.
Another type of feedback is called metalinguistic feedback. Lyster and Ranta (1997) referred to this type of feedback as “comments, information, or questions related to the well-formedness of the student’s utterance, without explicitly providing the correct form” (p. 47). Metalinguistic information can provide the learners with a variety of hints related to possible reformulations of the target forms that contain errors. (e.g., “It’s plural”) or a word definition in the case of lexical errors (Lyster & Ranta, 1997).
Some studies that have examined the effectiveness of metalinguistic and recast feedback show that first feedback produced better results because it assisted learners in the process of language learning. For instance, the study carried out in a Chinese EFL setting by Yang & Lyster (2010) reported that by providing metalinguistic feedback, learners increased their accuracy in the use of regular past tense. Additional supportive evidence is that of Lyster and Ranta (1997). The results of their study of students of “French immersion” courses demonstrated that metalinguistic feedback led to learner uptake in 90%, whereas 31% was obtained after the provision of recast feedback. These results also seem to suggest that recasts are not very effective in eliciting learners’ uptake.
Methodology
This study employs a quantitative nonexperimental longitudinal explanatory research methodology. Johnson (2001) highlights one of the advantages of explanatory research when researchers attempt “to explain how the phenomenon operates by identifying the causal factors that produce change in it” (p. 9). It is also longitudinal because the data was collected at different moments during a specific period of time and non-experimental since there was not a random selection of participant students.
The focus of this study is to identify whether there is a relationship between the kind of teacher feedback and student uptake. In order to identify the difference between the kind of teacher feedback and student uptake, two independent variables were identified, for instance: recast and metalinguistic feedback. This was complemented with four dependent variables: omission of subject, auxiliary use in questions, subject-verb agreement, and reported statements. Thirty students attending their last mandatory EFL course in the language center of a public university were chosen as participants during the semester from October 2015 to February 2016. Since these students comprised two different EFL classes, it was not necessary to divide them into two groups. Instead, these classes as a whole constituted the two groups that comprised this study. Students in Group 1 (. = 16) were exposed to metalinguistic feedback strategy and Group 2 (. = 14) to recast feedback. The type of feedback to be used in each group was randomly done. For the random selection of the type of feedback, two students (one per each group) were asked to choose one closed envelop from a group of two. Each envelop contained a card indicating the type of feedback, for instance one envelop contained recast and the other metalinguistic feedback. Hence, the type of feedback selected by the two students was used throughout the study. Students in these two groups were aged 18 to 20 years old. The participants had been learning English for one and a half years. They had an intermediate level of English which allowed them to participate in the task-based interaction used as part of the treatment in this study. All participants had a B1 level of English according to the Common European Framework of Reference. This was identified through the final EFL exam which all participants completed to graduate from seventh to eighth course of English. Participants’ written consent was obtained previous the beginning of this research. Finally, the class teacher agreed that the researcher would be the one who conducted all the activities. A written consent was also provided to the researcher in order to carry out this study with the two groups.
Regarding the grammar structures used in the feedback activities, these emerged as the result of the administration of a survey completed by 15 teachers of the eighth EFL course in the language center of the selected university. All teachers were asked one open-ended question. This question was: “based on your experience, what grammar structures do you think are the most difficult for students to acquire?” In order to proceed with the subsequent selection of the grammar structures, the top four most recurrent teachers’ answers were selected. The results were as follows: 100% of teachers mentioned that auxiliary use in questions was the most difficult grammar structure, followed by the omission of subject and reported statements both representing the 70% of teachers’ answers. Finally, subject-verb agreement scored fourth with 68%. Furthermore, students’ knowledge of these structures was diagnosed via a multiple choice test. This test was designed by the researchers and piloted in a different class to check its reliability. Once it was piloted, this test was conducted to the two groups. The scores obtained by the students in this diagnostic test were collected in a record sheet which confirmed that the students did have difficulty in the structures mentioned by their teachers.
For purposes of this study and application of treatments, 18 sessions (nine sessions per group) were planned. Each session lasted 120 minutes during a period of eight weeks. The first group was exposed to recast feedback for nine sessions, and the second group to metalinguistic feedback for nine sessions.
In order to measure correct language repairs, students were first engaged in the following activities: (1) comments to five videos of approximately five minutes each, and (2) comments on four short readings. These activities were formulated with the objective of facilitating English interactions among students and to provide them with enough English language input through videos and reading activities. The data for analysis included the interactions that students did with the teacher.
The procedure in each activity was as follows: First, the theme was introduced in the same way it is usually done in a normal class. Second, students were introduced to focused tasks which targeted the use of the four grammatical structures. Third, instructions were given to students regarding the tasks they had to perform. Useful words were reviewed but no new structures were taught. This stage was for preparation prior to the task cycle, which consisted of three elements:
(A) Task: This was done by students individually and in pairs. The researcher supervised the students, but did not intervene to correct errors.
(B) Preparation: Students had to watch the video of approximately 5-6 minutes or read the chosen text, individually prepare a one-minute oral summary to present it, have four questions ready to ask his or her partner about the video or text, and be prepared to report four statements said by his or her partner.
(C) Report: Each student had to interact with a partner selected by the researcher at least four times in each task. The total number of interventions per student was 16. During the intervention of the students, the researcher provided either recast or metalinguistic feedback that corresponded to each group. The feedback was provided to each pair without the presence of the other students.
A record sheet was used to collect data in order to register the type of student response (uptake) to the corresponding feedback. Based on the on the existing categories of uptake sequences proposed by Lyster and Ranta, (1997), uptake was then coded as either “repair” or “needs repair.” For this study, the following verbal responses were considered “uptake” as presented in the following table:
Table 1: Types of Recast and Learner’s Uptake

The frequency and percentage of students’ error repairs were calculated with the values obtained. The values were expressed numerically and graphically. This helped determine the feedback treatment that led students to have the most successful repairs. A final survey was given to students who participated in the study. This helped analyze the degree of acceptance of the feedback received.
Analysis and Discussion of Results
When the metalinguistic feedback strategy was used with Group 1, the number of error repairs steadily increased during the nine sessions. The number of error repairs improved from 89 in session one to 151 in session nine. On the other hand, students’ uptake that led to no repair was identified. The students modified errors but did not repair, decreasing from 79 in session one to 52 in session nine. Likewise, the students repeated the errors or left them unmodified without repair during the nine sessions (71 times in the first session, to 53 times in the last one; see Figure 1). This reduction in both modified and unmodified uptakes is related to the increase in repairs.
When the recast feedback strategy was applied to Group 2, the number of error repairs slightly increased during the nine sessions. The number of error repairs improved from 52 in session one to 84 in session nine. On the other hand, the number of errors that the students modified but did not repair as well as the ones that the students repeated or unmodified was higher than the number of repairs until session four. It is from session five that the number of repairs exceeds the number of no repairs. When evaluating recast feedback, acknowledgement, which is another type of uptake, was also registered. Acknowledgement was stable (27-28) from the first to the sixth session, and was increasing from the seventh session to the last one (36-38; Figure 2). This student uptake may have occurred due to the repeatability of the strategy.
With the metalinguistic feedback strategy in Group 1, 105 errors were committed in the last session (Table 2). This implies that 41% of the errors were not repaired from the total of 256 possible interactions. To exemplify how students repaired their errors when using metalinguistic feedback, the following quote includes an interaction between a student (S) and the researcher (R) testing a subject verb agreement:
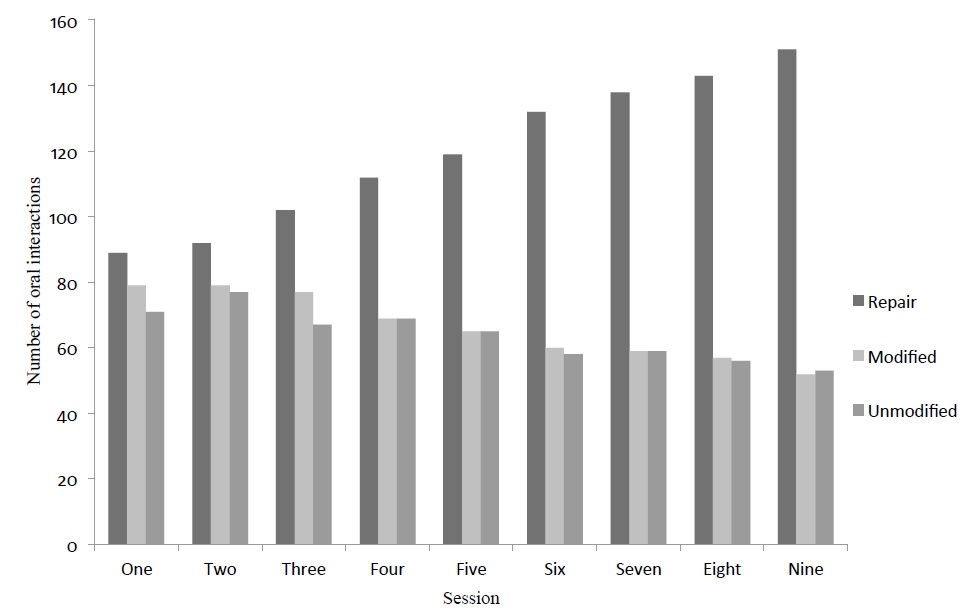
Figure 1: Effect of metalinguistic feedback strategy on student uptake (error repair, modified and unmodified error through nine intervention sessions)
S: The last two questions was especially hard.
R: Is the word “questions” singular or plural?
S: It’s plural, ok then... were especially hard
Here, the researcher promotes self-reflection of the error produced by the student by emphasizing the grammatical number through the word ‘questions’ drawing the student’s attention to the noun. The student noticed the correct use of the verb to be (subject verb agreement). For further statistical analysis of the results of metalinguistic feedback, the mean of non-repaired errors of the four grammatical structures (omission of subject, subject-verb agreement, auxiliary use in questions, and reported statements) collected in session nine using this feedback strategy (Table 2) was lower than the mean of errors registered in the diagnostic test on the aforementioned structures. Additionally for testing the statistical significance of this study an independent sample T-test was calculated. The results of this test demonstrated statistical significance for the first session (. = .001) as well as from the second to the ninth session (p = .000).
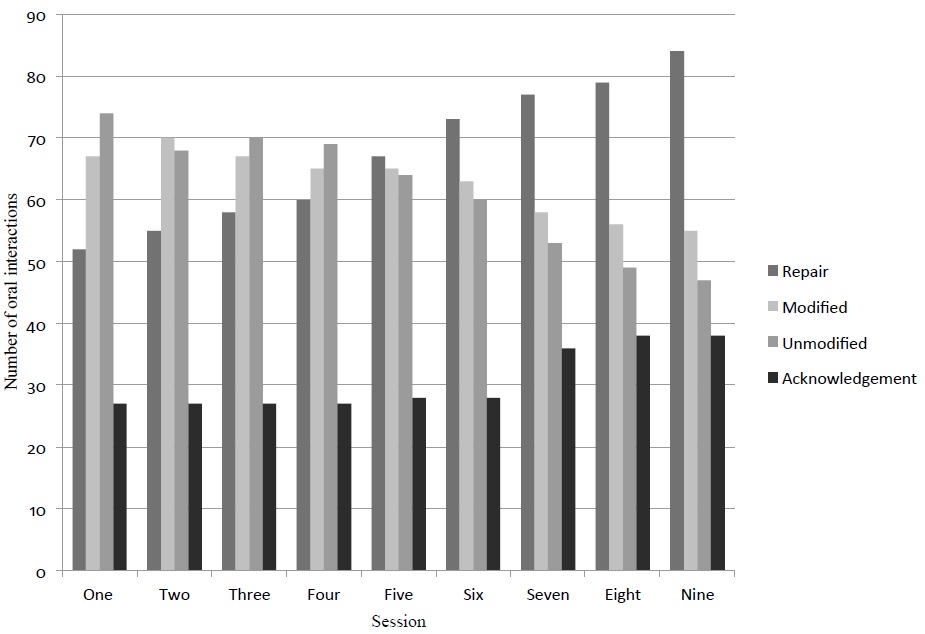
Figure 2: Effect of recast feedback strategy on student uptake (error repair, modified, unmodified error and acknowledgement) through nine intervention sessions.
Considering that values which are lower than 0.05 are statistically significant; for this study, the null hypothesis was rejected. This means that the students who received a metalinguistic feedback repair more errors that students who received a recast feedback.
The standard deviation of non-repair errors of the four grammatical structures in session nine using metalinguistic feedback strategy (Table 2) was lower in comparison with the standard deviation of errors registered in the diagnostic test.
With the recast feedback strategy in Group 2, 140 errors were committed in the last session (Table 3). The interaction occurred between a student (S) and the researcher (R) are encapsulated in the following interaction:
S: The girl in the park watched the movie twice.
R: The girl in the park has watched the movie twice.
S. Yes, the girl has watched the movie twice.
Table 2: Number of non-repairs registered in the last session with metalinguistic feedback strategy
The above excerpt shows how the researcher uses a recast feedback with the student. In this interaction, the researcher does not use extra clues to raise the student’s language awareness. The researcher limits her feedback to the repetition of the structure using the correct form of subject verb agreement and emphasis on the correct structure is done. This emphasis is expressed in bold (has). Data regarding this kind of feedback manifested that 63% of the errors were not repaired from the total of 224 possible interactions. The mean of non-repaired errors of the four grammatical structures (omission of subject, subject-verb agreement, auxiliary use in questions, and reported statements) collected in session nine using this feedback strategy (Table 3) was lower than the mean of errors registered in the diagnostic test on the aforementioned structures. The standard deviation of non-repair errors for the four grammatical structures in session nine using recast feedback (Table 3) was lower in comparison with the standard deviation of error registered in the diagnostic test.
The two feedback strategies result in a reduction of errors during oral interactions. Fifty-nine percent of grammatical errors were repaired by using metalinguistic feedback in comparison with 38% of errors repaired by using recast feedback. Therefore, metalinguistic feedback, based on the data, proved to be more effective than recast.
Twenty-one non-uptakes were registered during interactions using metalinguistic feedback in contrast to nine no-uptakes during interactions using recast feedback. Perhaps this occurred because the metalinguistic strategy elicited answers from students instead of providing them, so some students decided not to respond (Figure 3).
Survey Results
An acceptance survey was applied to the two groups. The results of the survey to the two types of feedback strategies of metalinguistic and recast were 90% of the students who thought that receiving feedback was interesting and that contributed to improving their oral production. Seventy-nine percent of the students graded the degree of contribution of the feedback strategies to their oral production as excellent, 13% as very good, and 8% as good. This suggests that students who participated in this study are open to receiving feedback from teachers.
Table 3: Number of nonrepairs registered in the last session with recast feedback Strategy
Number
Names
Grammatical
Structure
Omission
Of subject
Subject-verb
Agreement
Aux-use in questions
Reported
Statements
1
Elizabeth
3
2
2
2
2
Luis
3
3
2
2
3
Alexander
2
3
3
2
4
Alejandra
2
3
3
2
5
Natan
3
2
3
3
6
Veronica
2
3
2
3
7
Eduardo
2
3
2
3
8
Michael
3
3
2
1
9
Francois
3
2
2
3
10
Lucia
3
3
2
3
11
Joel
2
3
2
2
12
David
3
2
2
3
13
Iveth
2
4
3
2
14
Yolanda
3
3
2
2
Total of errors
36
39
32
33
Mean
2,57
2,79
2,29
2,36
Standard deviation
0,51
0,58
0,47
0,63
Minimum
2
2
2
1
maximum
3
4
3
3
Number of actions
56
56
56
56
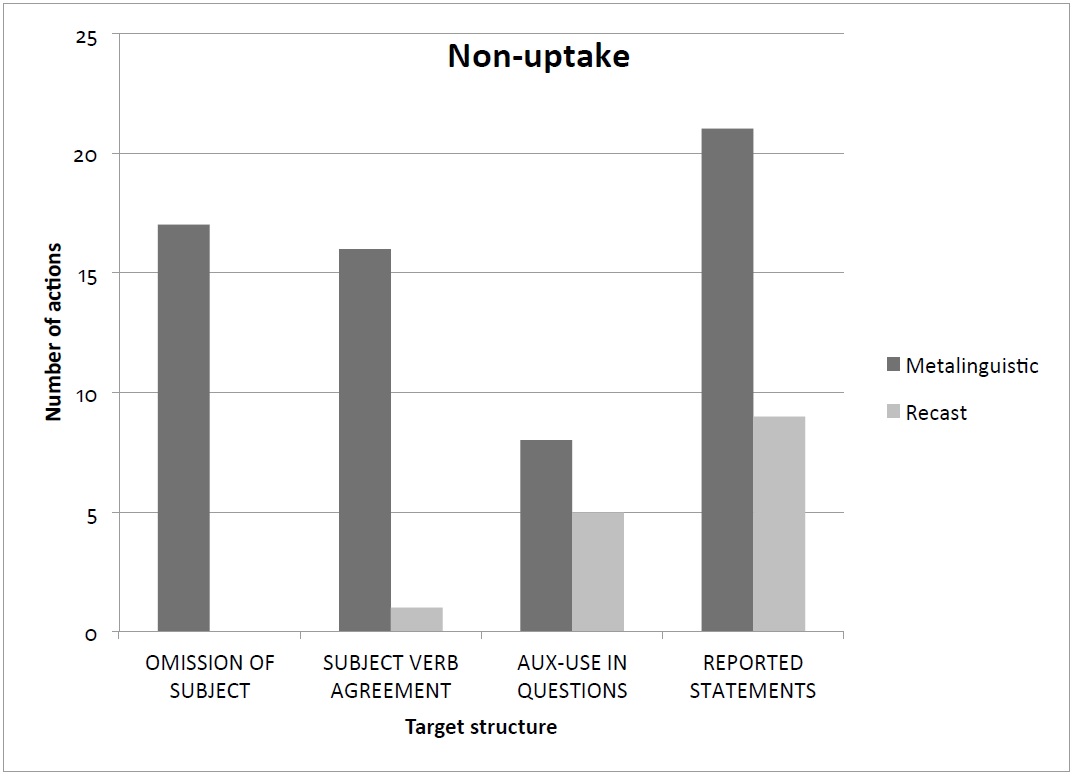
Figure 3: Number of non-uptake during metalinguistic and recast feedback sessions
This study was designed to compare the effects of two different feedback strategies on learner uptake during oral interactions. The result of this study showed that students repaired more errors after being exposed to feedback strategies. This indicates that exposing students to the target structures in combination with consciousnessraising activities such as error correction can improve their production. This finding is supported by Swain (1995) who claimed that corrective feedback assists students to test their hypotheses about the target language and replace them with the correct ones. This is also in line with Schmidt’s (1990) noticing hypothesis which highlights the role of feedback in drawing students’ attention to the target structures and comparing with their own forms, so as to correct their errors.
The analysis of data showed that metalinguistic feedback led to higher uptake results than recasts (Tables 2 & 3), since the participants exposed to metalinguistic corrective feedback outperformed those receiving recasts during communicative interactions. One explanation can be that metalinguistic feedback is “output-pushing” as suggested by Yang and Lyster (2010). While producing output, learners in this study faced some difficulties in conveying their meaning, but by exposing them to metalinguistic feedback, they were pushed to analyze and modify their output (Figure 1). This is in line with Swain (1995) who suggested that output practice can facilitate acquisition because it permits the development of cognitive processes, such as noticing, hypothesis testing, and metalinguistic reflection.
The findings in the current study also lend support to Schmidt’s (2001) hypothesis which emphasizes the importance of drawing learners’ attention to formal aspects of language for accomplishing linguistic development. In this case, metalinguistic feedback stimulates learners to openly identify the gap between their interlanguage and the target forms. Similarly, Hendrickson (1978) emphasized that students’ interlanguage can be developed by motivating them to produce more comprehensible and appropriate output rather than simply telling them the correct target forms.
One justification behind the reduced effectiveness of recast in this study can be the fact that this group of learners is accustomed to selfproduction without feedback since the emphasis at the institution’s language center has always been on meaning rather than form. Therefore, students may have ignored the teacher’s reformulation as corrective feedback assuming it was mere positive evidence for their output. Research by Panova and Lyster (2002) seems to support the claim that recasts can be ambiguous to foreign language learners as they may tend to repeat mechanically what the teacher says without any significant learning. In their study, Panova and Lyster (2002) reported that metalinguistic prompts led students to develop better formulated utterances than those from recast. In this case, recasts appeared not to be very effective because learners, contrary to metalinguistic feedback, were just exposed to the correct form of target structures as modified input, without noticing the important role of output; that is, forcing learners to correct their interlanguage and elicit their self-repair. Metalinguistic feedback seems to be particularly helpful to these participating groups during the process of learning of specific target forms. The guidance provided by the teacher through this type of feedback can be a worthwhile investment of time and effort for teachers and students.
As far as non-uptake is concerned, certain students did not provide any verbal response immediately following teacher feedback. Chaudron (1977) viewed uptake as a main immediate measurement of the effectiveness of corrective feedback. When uptake with repair is produced, it is an indication that learner’s attention to the correct forms has been drawn and that learners have taken a step toward learning, while the lack of uptake can show that learners did not notice the mismatch between their statements and the target ones after the provision of feedback. This belief is opposed to Nassaji (2009) who mentioned that the absence of uptake is not an evidence of no noticing which can be assumed that the provision of feedback is not effective. It is possible that learners in this study have processed and learned from the feedback in their inner thought, but did not want to respond to it. Contextual variables may also have been intertwined in the uptake moves. For example, attitudes, concepts about learning, and motivation can influence how learners actually approach tasks and respond to feedback. The existence of those connections is acknowledged by Dörnyei and Skehan (2003) who mention that “there are individual differences between learners in noticing abilities: other things being equal, some learners may be more likely to notice relevant qualities of input than others” (p. 597).
The acceptance survey showed that learners thought feedback was interesting and did not feel uncomfortable when receiving feedback. Some learners’ comments mentioned that they were receptive to feedback and were more willing to incorporate it because they perceived the correction as a manifestation of lending a helping hand, rather than a display of mockery, and the feedback was done individually and not shared in a group setting.
Conclusions
The findings of this study highlight the importance of corrective feedback in the process of English learning. In this regard, teachers can respond to students’ errors through two kinds of feedback, both metalinguistic and recast. Concerning these two types of feedback, it emerged from the data that metalinguistic feedback is more effective to students rather than recast. Its effectiveness may be correlated to students’ selfrealization of their mistakes facilitated through “comments, information, or questions related to the well-formedness of the student’s utterance, without explicitly providing the correct form” (Lyster & Ranta, 1997, p. 47). Teachers, therefore, need to become active key players in providing proper clues to improve students’ language awareness and subsequent reduction of errors. In this study, the data manifested that more exposure to metalinguistic feedback results in better error correction.
Metalinguistic feedback proves to be more effective because it encourages students more actively to self-repair, contrary to recast, when the correct form is provided to the students. This was reaffirmed by the data that the highest rate of uptake and successful repairs resulted from metalinguistic feedback, whereas recast feedback led to the lowest rate of uptake and successful repairs.
Complementary to the results, a survey of students who participated in the study was applied in order to determine the level of acceptance of the feedback received. The survey results manifested a positive opinion towards metalinguistic feedback and added that this feedback has contributed to the improvement of their oral production.
Comparing the effects of these two feedback strategies provided some light on theoretical issues such as the important contributions of corrective feedback in the EFL classroom and the cognitive role of noticing during the process of feedback. The results of this study also provide teachers with pedagogical alternatives to maximize the effect of feedback in their classes.
References
Notas
Metrics
License
Copyright (c) 2017 Monica Raquel Tamayo

This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License.

This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License.
Attribution — You must give appropriate credit, provide a link to the license, and indicate if changes were made. You may do so in any reasonable manner, but not in any way that suggests the licensor endorses you or your use.
NonCommercial — You may not use the material for commercial purposes.
NoDerivatives — If you remix, transform, or build upon the material, you may not distribute the modified material.
The journal allow the author(s) to hold the copyright without restrictions. Also, The Colombian Apllied Linguistics Journal will allow the author(s) to retain publishing rights without restrictions.



