
DOI:
https://doi.org/10.14483/2256201X.13797Publicado:
24-06-2019Número:
Vol. 22 Núm. 2 (2019): Julio-diciembreSección:
Artículos de investigación científica y tecnológicaVariación genética en familias de polinización abierta de Tectona grandis L.f. en Colombia
Genetic variation among half-sib families of Tectona grandis L.f. in Colombia
Palabras clave:
forest management, breeding, genetic parameters, selection, teak, wood volume (en).Palabras clave:
manejo forestal, mejoramiento genético, parámetros genéticos, selección, teca, volumen de madera (es).Descargas
Referencias
Angiosperm Phylogeny Group (APG) (2009). An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG III. Botanical Journal of the Linnean Society, 161(2), 105-121.
https://doi.org/10.1111/j.1095-8339.2009.00996.x
Barros, K. L. C., Macedo, R. L. G. y Venturin, N. (2017). Capacidade de brotação de teca sob regime de Talhadia composta. Floresta e Ambiente, 24, e00134315.
http://doi.org/10.1590/2179-8087.134315
Becker, W. A. (1975). Manual of quantitative genetics. Pullman, WA: Washington State University Press.
Blada, I. (1994). Performance of open-pollinated families of blue pine in Romania. Silvae Genetica, 43(4), 231-238.
Blanco-Flórez, J., Fernando-Trugilho, P., Tarcisio-Lima, J., Gherardi-Hein, P. R., y Moreira da Silva, J. R. (2014). Caracterización de la madera joven de Tectona grandis L. f. plantada en Brasil. Madera y Bosques, 20, 1, 11-20.
https://doi.org/10.21829/myb.2014.201172
Callister, A. N. y Collins, S. L. (2008). Genetic parameter estimates in a clonally replicated progeny test of teak (Tectona grandis Linn. f.). Tree Genetics and Genomes, 4, 237-245.
https://doi.org/10.1007/s11295-007-0104-2
Chaix, G., Monteuuis, O., Garcia, C., Alloysius, D., Gidiman, J., Bacilieri, R. y Goh, D. K. S. (2011). Genetic variation in major phenotypic traits among diverse genetic origins of teak (Tectona grandis L.f.) planted in Taliwas, Sabah, East Malaysia. Annals of Forest Science, 68, 1015-1026.
https://doi.org/10.1007/s13595-011-0109-8
Cornelius, J. P., Mesén, J. F. y Corea, E. A. (1991). Manual sobre mejoramiento genético forestal con referencia especial a América Central. Turrialba: Centro Agronómico Tropical de Investigación y Enseñanza (Catie).
Cornelius, J. P. (1994). Heritabilities and additive genetic coefficients of variation in forest trees. Canadian Journal of Forest Research, 24, 372-379.
https://doi.org/10.1139/x94-050
Espitia, M., Murillo, O. y Castillo, C. (2011). Ganancia genética esperada en teca (Tectona grandis L.f.) en Córdoba (Colombia). Colombia Forestal, 14(1), 81-93.
https://doi.org/10.14483/udistrital.jour.colomb.for.2011.1.a07
Fleitas-Camacho, Y., Pérez-Santana, M. H., Echeverría-Caraballo, P., González-Roque, A., Rivero-Vega, M., González-Vera, E. S., Fúster-Mancha, J. A. y Placencia-Puentes, T. (2010). Comportamiento de descendencias de Tectona grandis (L. f.) en Pinar del Río. Revista Forestal Baracoa, 29(1), 31-39.
Fonseca, W. (2003). Manual para productores de teca (Tectona grandis L.f.) en Costa Rica. Heredia: Fondo Nacional de Financiamiento Forestal.
Francis, J. K., Lowe, C. A. y Trabanino, S. (2000). Bioecología de árboles nativos y exóticos de Puerto Rico y las Indias Occidentales. Puerto Rico: Departamento de Agricultura de los Estados Unidos, Servicio Forestal, Instituto Internacional de Dasonomía Tropical.
https://doi.org/10.2307/3539166
Galeano, E., Vasconcelos, T. S., Vidal, M., Mejia-Guerra, M. K. y Carrer, H. (2015). Large-scale transcriptional profiling of lignified tissues in Tectona grandis. BMC Plant Biology, 15(1), 221.
https://doi.org/10.1186/s12870-015-0599-x
Goh, D. K. S., Japarundin, Y., Alwi, A., Lapammu, M., Flori, A. y Monteuuis, O. (2013). Growth differences and genetic parameter estimates of 15 teak (Tectona grandis L.f.) genotypes of various ages clonally propagated by microcuttings and planted under humid tropical conditions. Silvae Genetica, 62(4-5), 196-206.
https://doi.org/10.1515/sg-2013-0024
Harris, F. C. (2007). The effect of competition on stand, tree, and wood growth and structure in subtropical Eucalyptus grandis plantations (tesis de doctorado). Southern Cross University, Lismore, NSW, Australia.
Holdridge, L. R. (1947). Determination of world plant formations from simple climatic data. Science, 105(2727), 367-368.
https://doi.org/10.1126/science.105.2727.367
Instituto Geográfico Agustin Codazzi (Igac) (2004). Estudio general de suelos y zonificación de tierras departamento de Tolima. Bogotá: Instituto Geográfico Agustin Codazzi (IGAC), Subdirección de agrología, grupo interno de trabajo levantamiento de suelos.
https://doi.org/10.21500/20275846.1721
Ipinza, R. H. (1998). Mejoramiento genético forestal. Bogotá: Corporación Nacional de Investigación y Fomento Forestal (Conif), Serie técnica n° 42.
Isik, F. (2009). Analysis of half-sib progeny tests in forestry. Raleigh: North Carolina State University and Departament of Forestry and Enviromental Resources.
Kollert, W. y Cherubini, L. (2012). Teak resources and market assessment 2010 (Tectona grandis Linn. F). Roma: Food and Agriculture Organization (FAO), Planted Forests and Trees Working Paper FP/47/E.
Leiva, S. (2009). Propagación vegetativa de la teca (Tectona grandis L.f.) por el método de miniestacas. Bogotá: Corporación Nacional de Investigación y Fomento Forestal (Conif).
Llanos, L. F. (2017). Selección de progenies superiores de Tectona grandis L.f. en el norte del Tolima (te¬sis de pregrado, Ingeniería Forestal). Universidad del Tolima, Ibagué, Colombia.
https://doi.org/10.14482/pege.43.10583
Lokmal, N. (1994). Genetic parameters of Gmelina arborea: height and diameter growth. Journal of Tropical Forest Science, 7(2), 323-331.
López, A. M., Barrios, A., Trincado, G. y Nieto, V. M. (2011). Monitoreo y modelamiento del crecimiento para el manejo de plantaciones forestales comerciales. Bogotá: Corporación Nacional de Investigación y Fomento Forestal (Conif), Ministerio de Agricultura y Desarrollo Rural (Madr).
Lynch, M. y Walsh, B. (1998). Genetics and analysis of quantitative traits. Massachusetts: Sinauer Associates, Inc.
Matínez, C. (2012). Estadistica y muestreo. Bogotá: Ecoe ediciones.
Meza, A., Rodríguez, J., Gatti, K. C. y Espinoza, E. (2015). Propagación de arboles de teca Tectona grandis L. f. por miniestacas. Revista Temas Agrarios, 20(2), 43-48.
https://doi.org/10.21897/rta.v20i2.757
Mora, F. y Zamudio, F. (2006). Variabilidad genética de crecimiento en progenies selectas de Pinus radiata. Ciência Florestal, 16(4), 399-405.
http://doi.org/10.5902/198050981921
Narayanan, C., Chawhaan, P. H. y Mandal A. K. (2009). Inheritance pattern of growth and wood traits in teak (Tectona grandis L.f.). Silvae Genetica, 58(3), 97-101.
https://doi.org/10.1515/sg-2009-0013
Ordóñez, J. L. (2013). Plantaciones de teca: Negocio en alza. Revista el Mueble y la Madera, 80, 20-25.
Pastrana-Vargas, I. J., Espítia-Camacho, M. y Murillo-Gamboa, O. (2012). Evaluación del potencial de mejoramiento genético en el crecimiento en altura de Acacia mangium Willd. Acta Agronómica, 61(2), 143-150.
Pavlotzky, B. y Murillo, O. (2013). Ganancia genética esperada en Acacia mangium en los Chiles, zona norte de Costa Rica. Agronomía Mesoamericana, 23(1), 93-106.
https://doi.org/10.18845/rfmk.v10i24.1320
Pereira, G. (1994). Crecimiento y patrones de heredabilidad de la progenie de polinización abierta de Crupressus lusitanica en diez ensayos a la edad de diez años. Bogotá.: Instituto Nacional de los Recursos Naturales Renovables y del Ambiente (Inderena).
https://doi.org/10.7705/biomedica.v23i4.1230
Raj, A., Sehgal, R. N., Sharma, K. R. y Sharma, P. K. (2010). Genetic variation in wood specific gravity among half-sib families of chir pine (Pinus roxburghii sargent). New Forests, 40, 213-227.
https://doi.org/10.1007/s11056-010-9195-y
Rivero, J. y Moya, R. (2006). Propiedades físico-mecánicas de la madera de Tectona grandis Linn. F. (teca), proveniente de una plantación de ocho años de edad en Cochabamba, Bolivia. Kurú: Revista Forestal, 3(9), 1-14.
https://doi.org/10.18845/rfmk.v13i30.2460
Rodríguez, J., Urueña, E. J. y Borralho, N. (2011). Mejoramiento de semillas para incrementar las plantaciones de teca (Tectona grandis L.F.) y melina (Gmelina arborea Roxb.) en el departamento del Tolima. Bogotá: Corporación Nacional de Investigación y Fomento Forestal (Conif), Ministerio de Agrícultura y Desarrollo Rural (Madr).
Ruotsalainen, S. (2014) Increased forest production through forest tree breeding. Scandinavian Journal of Forest Research, 29(4), 333-344.
https://doi.org/10.1080/02827581.2014.926100
Sharma, R., Swain, D. y Mandal, A. K. (2000). Estimates of genetic parameters from an open pollinated genetic test of teak (Tectona grandis). Journal of Tropical Forest Science, 12(1), 44-48.
Sreekanth, P. M., Balasundaran, M. y Nazeem, P. A. (2014). Genetic and morphological variation in natural teak (Tectona grandis) populations of the Western Ghats in Southern India. Journal of Forestry Research, 25(4), 805-812.
https://doi.org/10.1007/s11676-014-0528-0
Stastical Analysis System Institute Inc. (SAS) (2009). SAS/STAT® 9.2 User’s Guide, Second Edition. Cary, EE. UU.: SAS® Publishing.
Unidad de Planificación Rural Agropecuaria (Upra) (2015). Zonificación para plantaciones forestales con fines comerciales-Colombia, escala 1:100.000. Memoria técnica. Bogotá: Unidad de Planificación Rural Agropecuaria (Upra).
Tambarussi, E. V., Rogalski, M., Galeano, E., Brondani, G. E., Martin, V. F., Silva, L. A. y Carrer, H. (2017). Efficient and new method for Tectona grandis in vitro regeneration. Crop Breeding and Applied Biotechnology, 17, 124-132.
http://doi.org/10.1590/1984-70332017v17n2a19
Vallejo, A. y Avendaño, J. (2013). Modelos de crecimiento y rendimiento. En R. de Camino y J.P. Morales (eds), Las plantaciones de teca en América Latina: mitos y realidades (pp. 114-132). Turrialba: Centro Agronómico Tropical de Investigación y Enseñanza (Catie).
Vásquez, A. (2001). Silvicultura de plantaciones forestales en Colombia. Ibagué: Universidad del Tolima, Facultad de Ingeniería Forestal.
Vergara, C., Cardona, C., Murillo, O., Jarma, A. y Araméndiz, H. (2013). Valor de mercado de plantaciones de teca (Tectona grandis Linn.) en el departamento de córdoba. Revista Temas Agrarios, 18(1), 9-22.
https://doi.org/10.21897/rta.v18i1.705
Yasodha, R., Vasudeva, R., Balakrishnan, S., Sakthi, A. R., Abel, N., Binai, N., Rajashekar, B., Bachpai, V. K. W., Pillai, C. y Dev, S. A. (2018). Draft genome of a high value tropical timber tree, Teak (Tectona grandis L. f): insights into SSR diversity, phylogeny and conservation. DNA Research, 25(4), 409-419.
https://doi.org/10.1093/dnares/dsy013
Zobel, B. J. y Talbert, J. T. (1988). Técnicas de mejoramiento genético de árboles forestales. México D.F.: Editorial Limusa S.A.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Recibido: 29 de agosto de 2018; Aceptado: 4 de junio de 2019
Resumen
La teca es una especie comercial de alto interés, principalmente por las propiedades físico-mecánicas de su madera, tasas de crecimiento y adaptabilidad. El objetivo del estudio fue evaluar la variación genética de rasgos de crecimiento y calidad del árbol entre 40 familias de polinización abierta. Usando datos de un ensayo de progenies de cuatro años, se calcularon heredabilidades en sentido estricto, ganancias genéticas, correlaciones genéticas y se construyó un ranking de familias de acuerdo con el volumen total de madera por hectárea. Las familias mostraron una supervivencia alta entre 73-100 %. Las ganancias genéticas seleccionando 30 % de las mejores familias fueron de 2.93 % para diámetro, 4.83 % para volumen del fuste, 11.97 % para volumen por hectárea, 6.53 % para área de proyección de copa, 4.26 % para ángulo de inserción de ramas y 14.42 % para número de renuevos. Se identificaron tres familias con productividad superior y potencial para el establecimiento de plantaciones de alto rendimiento.
Palabras clave:
manejo forestal, mejoramiento genético, parámetros genéticos, selección, teca, volumen de madera..Abstract
Teak is a commercial tree species of high interest, mainly for its outstanding wood physical and mechanical properties, growth rates and adaptability. The aim of the study was to evaluate the genetic variation of growth and tree quality traits among 40 half-sib families. Using data from a 4-year-old progeny test, narrow sense heritabilities, genetic gains, and genetic correlations were calculated, as well as a ranking of families according to total wood volume per hectare. All the families showed a higher survival rate ranging from 73-100 %. The genetic gains by selecting 30 % of the best families were of 2.93 % for diameter, 4.83 % for stem volume, 11.97 % for total volume per hectare, 6.53 % for crown projection area, 4.26 % for branch insertion angle and 14.42 % for number of basal sprouts. Three families with superior productivity and potential for the establishment of high yield plantations were identified.
Keywords:
breeding, genetic parameters, selection, teak, forest management, wood volume..INTRODUCCIÓN
Tectona grandis L.f. (teca) pertenece a la familia Lamiaceae (APG, 2009), es distinguida por tener una alta demanda y un alto valor en el mercado internacional (Ordoñez, 2013), principalmente por las excelentes características de su madera, tales como su resistencia, trabajabilidad, alta calidad estética, buena duración natural y una amplia variedad de usos industriales (Rivero y Moya, 2006; Blanco-Flórez et al., 2014; Yasodha et al. 2018). La especie es originaria de hábitats húmedos hasta secos, pertenecientes al sudeste de Asia, India, Birmania, Laos y Tailandia (Francis et al., 2000; Sreekanth et al., 2014; Tambarussi et al. 2017).
En el año 2010 se estimó un área plantada con teca de 4.35 millones de hectáreas en el mundo; en Suramérica se registraron 122 300 ha (Kollert y Cherubini, 2012). Para el 2011, en Colombia existían alrededor de 25 000 ha de superficie plantada con la especie principalmente en la Costa Atlántica (Espitia et al., 2011), donde ha sido una de las principales especies empleadas en programas de reforestación comercial. El volumen de producción anual en madera rolliza es de 12 000 a 15 000 m³, y aproximadamente 6 000 a 8 000 m³ de madera aserrada (Ordoñez, 2013). A pesar de su importancia económica, aún existen vacíos de información respecto a la genética (Galeano et al., 2015) y la producción de material vegetal, el manejo de plantaciones de la especie, la industrialización y el comercio de madera (Ordoñez, 2013; Vergara et al., 2013; Meza et al., 2015).
Una gran proporción del éxito del negocio forestal basado en plantaciones radica en la calidad del material vegetal, que requiere el uso de semillas con características genéticas deseables y una adecuada producción en vivero (Ruotsalainen, 2014). Por lo tanto, para el establecimiento de plantaciones comerciales competitivas es necesaria la selección y desarrollo de fuentes de semilla o material propagativo con individuos que muestren superioridad genética en cuanto a las tasas de crecimiento, calidad del fuste y calidad de la madera (Pavlotzky y Murillo, 2013; Pereira, 1994; Vasquez, 2001). Actualmente en Colombia, existe un déficit de material vegetal de alta calidad para el fomento de plantaciones comerciales con esta especie en diferentes regiones país (Meza et al., 2015).
En el pais, se han realizado estudios encaminados al mejoramiento genético de la especie y su propagación vegetativa, liderados principalmente por empresas privadas (Leiva, 2009). Espitia et al. (2011) evaluaron árboles plus seleccionados en plantaciones de teca en el norte de Colombia (departamento de Córdoba) para determinar ganancias genéticas esperadas. Leiva (2009) y Meza et al. (2015) desarrollaron un protocolo para la propagación de árboles de teca a través del enraizamiento de miniestacas aplicando diferentes concentraciones de hormonas y antioxidantes. Sin embargo, no ha sido sustancial el avance que han tenido los programas de mejora genética en el interior del país, donde existe una superficie potencial para la reforestación con la especie de 1.76 millones de ha, principalmente en los valles interandinos en altitudes menores a 1000 m (UPRA, 2015). Solo el departamento del Tolima cuenta con un área de 221 463 ha con aptitud forestal alta (UPRA, 2015).
De esta forma, los programas de mejora genética tienen como objetivo seleccionar material genéticamente superior en una población base para producir plantaciones con aumento de características deseables paulatinamente (Pastrana-Vargas et al., 2012). El propósito al evaluar ensayos genéticos es determinar parámetros de la población de mejoramiento, mediante la separación eficiente de los efectos genéticos de los ambientales, así como seleccionar de manera eficiente los mejores genotipos con base en su mérito genético (Espitia et al., 2011). Las primeras etapas de estos programas incluyen la identificación de procedencias, seguida de la identificación de árboles plus en plantaciones comerciales, para posteriormente evaluar la capacidad de heredar atributos deseables en pruebas de descendencia o progenies (Cornelius et al., 1991; Zobel y Talbert, 1988).
Dentro de los parámetros genéticos, la heredabilidad, es uno de los más importantes y más usado en genética cuantitativa (Lynch y Walsh, 1998). El valor de la heredabilidad expresa la proporción de la variación en la población que es atribuible a diferencias genéticas entre individuos (Mora y Zamudio, 2006). Por otro lado, la ganancia genética es el mejor parámetro genético para definir la cantidad de mejoramiento obtenido para la especie dentro de un programa de mejora genética (Zobel y Talbert, 1988). Para T. grandis hay poca información sobre la heredabilidad y ganancia genética de rasgos de crecimiento y calidad del árbol y no hay datos cuantitativos sobre la relación entre estos rasgos. Los escasos estudios realizados reportan mejoras genéticas modestas de un 6 % en volumen comercial y aproximadamente un 22 % en calidad del fuste para teca en el departamento del Tolima (Rodríguez et al., 2011). De manera similar, estudios realizados en la costa norte de Colombia determinaron ganancias genéticas esperadas a partir de la selección de árboles plus de 2.35 % en diámetro y 12.5 % en volumen (Espitia et al., 2011). Otros estudios han reportado ganancias genéticas de 15.7 % en volumen para genotipos clonalmente propagados en Malasia (Goh et al., 2013).
El objetivo del presente estudio fue evaluar la variación genética de rasgos de crecimiento y calidad del árbol entre familias (árboles plus y progenies) de polinización abierta de T. grandis a los cuatro años de edad. Los objetivos específicos fueron: 1) determinar la heredabilidad de rasgos de crecimiento y de calidad del árbol y la ganancia genética como resultado de la selección de familias; 2) establecer correlaciones genéticas entre los rasgos de crecimiento y de calidad del árbol de mayor interés económico; y 3) identificar familias con altas tasas de producción de madera a través del análisis de los valores genéticos aditivos para cada familia.
MATERIALES Y MÉTODOS
Material vegetal
Semillas de polinización abierta de 40 árboles plus de T. grandis fueron recogidas y viverizadas para la obtención de material propagativo. Los árboles plus fueron seleccionados en plantaciones comerciales localizadas en los municipios de Puerto Libertador, San Antero y Canalete en el departamento de Córdoba y en el municipio de Plato, Magdalena (Rodríguez et al., 2011; tabla 1). Así, una prueba de progenies de medios hermanos fue establecida con las plántulas obtenidas de los 40 árboles plus durante marzo de 2011, usando un distanciamiento de siembra de 3 × 3 metros (1111 árboles.ha-1). El terreno fue preparado a través de un subsolado a 60 cm de profundidad sobre las líneas de siembra, permitiendo una mejor expresión del genotipo por la homogenización del terreno (Rodríguez et al., 2011). Se empleó un diseño experimental en bloques completos al azar (30 bloques), donde la unidad experimental estuvo constituida por cada árbol (Single Tree Plot).
Tabla 1: Localización de los árboles plus de T. grandis evaluados y características ambientales de los sitios de selección.
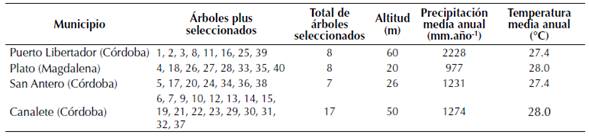
Área de estudio
La prueba de progenies está ubicada en el valle del rio Magdalena, en el municipio de Armero-Guayabal al norte del departamento del Tolima, en la coordenada 5°00’52.10’’ latitud norte-74°54’36’’ longitud oeste. El sitio se ubica a 280 m de altitud, pertenece a la zona de vida bosque seco tropical (bs-T) y está caracterizado por una temperatura media anual mayor a los 24 °C y precipitación media anual de 1700 mm (Holdridge, 1947). La geomorfología del paisaje es de piedemonte con un tipo de relieve abanico-terraza con pendientes bajas de 0-3 % y suelos de clase de textural franco arcillo arenosa (IGAC, 2004).
Datos
Las mediciones se realizaron en julio de 2015 cuando el ensayo contaba con 4.3 años de edad. Rasgos de crecimiento fueron medidos a todos los árboles, tales como diámetro a 1.3 m (d) empleando una cinta diamétrica, altura total (h) empleando un hipsómetro láser, radios de copa en las direcciones N-S-E-O empleando un distanciometro láser. El volumen del fuste sin corteza (vsc) fue calculado empleando el siguiente modelo general de volumen ajustado por López et al. (2011) a partir de una muestra de 50 árboles de T. grandis cosechados en la zona de estudio:
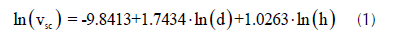
A partir de los radios de copa se calculó el área de la proyección de copa (APC) como,
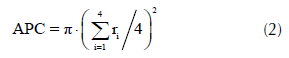
donde, ri son los radios de copa en las direcciones norte, sur, este y oeste.
Adicionalmente, a cada árbol se le registró rasgos de calidad como: número de renuevos (NR), rectitud fustal (RF), diámetro de ramas (DR) y ángulo de inserción de ramas (AIR). En el presente estudio los renuevos o chupones hace referencia a los brotes leñosos en la base del fuste del árbol. Para facilitar la valoración de los rasgos de calidad del árbol se empleó la codificación propuesta por Ipinza (1998), modificada por Llanos (2017) (tabla 2).
Tabla 2: Codificación de referencia de los rasgos de calidad del árbol. RF: Rectitud fustal, DR: Diámetro de ramas, AIR: Ángulo de inserción de ramas. d: diámetro a 1.3 m.

A partir del conteo del número de árboles, se determinó la sobrevivencia de cada familia, la cual se obtuvo como el cociente entre el número de individuos vivos sobre el total de individuos plantados o número de repeticiones, permitiendo observar las familias con mayor adaptabilidad a las condiciones ecológicas del área de estudio (Vásquez, 2001),
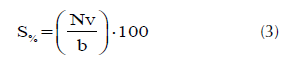
donde S%, Nv y ‘b’ es la sobrevivencia en porcentaje, número de árboles vivos y número de bloques o repeticiones, respectivamente.
Para cada familia se estimó el volumen por hectárea (Vha), expresado en m3.ha-1, considerando una densidad de plantación de 1111 árb.ha-1, y teniendo en cuenta la sobrevivencia de cada familia al momento de la medición del ensayo, así:
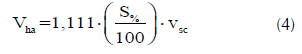
donde, S% es la sobrevivencia de la familia y vsc es el volumen del fuste.
Adicionalmente, al dividir el volumen por hectárea (Vha) en la edad (4.3 años) se obtuvo una estimación del incremento medio anual (IMA4 años) para cada familia.
Análisis de datos
Modelo estadístico y análisis de varianza
Todos los análisis estadísticos fueron realizados usando Statistical Analysis System (SAS Institute Inc., 2009). Debido a que el ensayo correspondió a un diseño de un árbol por parcela (STP) el siguiente modelo de análisis de varianza fue empleado para el análisis estadístico (Isik, 2009),
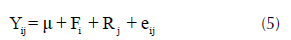
donde, Yij es una observación individual en la i-ésima familia en el j-ésimo bloque, μ es la media general, Fi es el efecto aleatorio de la i-ésima familia (i = 1, 2,…, 40) [NID (0,s2 f)], Rj es el efecto fijo del j-ésimo bloque (j = 1, 2, …, 30), y eij es el error aleatorio [NID (0,s2 e)]. Los componentes de varianza fueron estimados usando máxima verosimilitud restringida (REML), utilizando el procedimiento MIXED (SAS Institute Inc., 2009).
El modelo anterior evalúa qué parte de la variación fenotípica se debe a la genética y cuánto se debe al ambiente. Debido a que las familias representan una muestra aleatoria de la población (seleccionada al azar), la familia (F) y el error (e) dentro de la familia fueron considerados efectos aleatorios. Por el contrario, los bloques se consideraron como efectos fijos y fueron utilizados para controlar la variación ambiental al interior del ensayo; de esta manera, no se cuantificó la variación total explicada por los mismos.
Estimación de la heredabilidad
Las estimaciones de heredabilidad en sentido estricto fueron obtenidas siguiendo la formulación sugerida por Blada (1994),
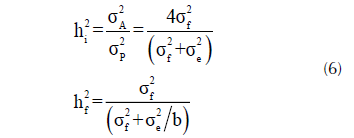
donde, h2 i y h2 f es la heredabilidad con respecto a la media individual y familiar, respectivamente, σ2 A y σ2 P son las varianzas aditiva y fenotípica, respectivamente, σ2 f y σ2 e son las varianzas debido a los efectos familiares y residuales y ‘b’ es el número de bloques o repeticiones. El error estándar de las heredabilidades estimadas fue calculado a través del método delta el cual toma en cuenta la relación entre varianza-covarianza (Lynch y Walsh, 1998, p. 561).
Ganancia genética
La ganancia genética esperada resultante de la selección familiar fue estimada usando la fórmula (Lokmal, 1994),

donde, h2 f es la heredabilidad familiar y S es el diferencial de selección. En el estudio se determinó la ganancia genética a tres diferentes niveles de intensidad de selección (50 %, 40 % y 30 %) de familias conservadas después del raleo genético (Becker, 1975; Raj et al., 2010).
Matriz de correlación genética
Para calcular la correlación genética entre los rasgos analizados, se utilizó un análisis de varianza multivariado (Manova) (Raj et al., 2010; Zobel y Talbert, 1988). La covarianza familiar entre los rasgos ‘x’ e ‘y’ (COVf(xy)) fue calculada como,
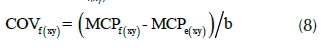
donde, MCPf(xy) y MCPe(xy) son las medias de los productos cruzados de los efectos familiares y residuales entre los rasgos ‘x’ e ‘y’, respectivamente, y ‘b’ es el número de repeticiones. Una vez calculada la covarianza, la correlación genética se calculó como:
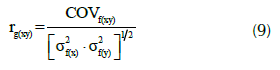
donde, rg(xy) es el coeficiente de correlación genético, COVf(xy) es la covarianza entre los rasgos ‘x’ e ‘y’, σ2 f(x) y σ2 f(y) son las varianzas debido a los efectos familiares para los rasgos ‘x’ e ‘y’. Una vez halladas las correlaciones genéticas se procedió a determinar la significancia de estos valores empleando una prueba estadística de t-student calculada a través de la siguiente fórmula (Martínez, 2012; p. 414):
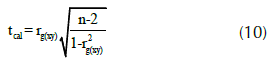
donde ‘n’ es el número de pares de datos observados (n = 1070), y tcal es el valor de t-student calculado.
Construcción de ranking genético para volumen total por hectárea
La habilidad combinatoria general de cada familia fue obtenida extrayendo los BLUP (best linear unbiased predictors) generados en el análisis de varianza respecto a la media global ajustada para las 40 familias evaluadas (Isik, 2009). Se construyó un ranking genético de las familias evaluadas de acuerdo con los valores genéticos aditivos obtenidos para el volumen total por hectárea. El cálculo del volumen total por hectárea integró los caracteres de crecimiento (diámetro y altura) y la sobrevivencia de cada familia.
RESULTADOS
Las 40 familias mostraron una buena adaptabilidad al sitio, con una sobrevivencia entre 73 y 100 % (figura 1), con un promedio de 89.17 % y un coeficiente de variación de 6.62 % (tabla 3). Del total de familias, solo 16 presentaron una sobrevivencia menor a la media, indicando que entre el grupo de familias evaluadas, hay un número importante de familias que se adaptan apropiadamente a las condiciones de sitio del ensayo. La familia 37 y 21 alcanzaron respectivamente la mayor (100 %) y menor (73.33 %) sobrevivencia, ambas familias son originarias de Canalete, Córdoba (tabla 3).
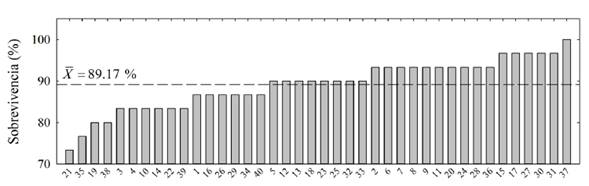
Figura 1: Sobrevivencia en porcentaje para 40 familias de polinización abierta de T. grandis a los 4.3 años. La línea horizontal discontinua representa la sobrevivencia media del ensayo.
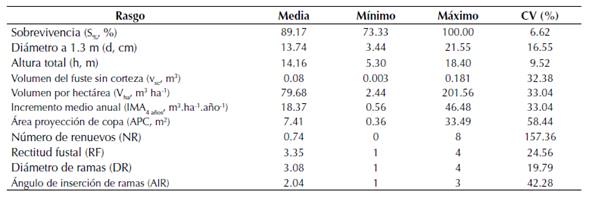
Los árboles medidos a los 4.3 años, presentaron un diámetro medio entre 3.4 a 21.6 cm (media de 13.73 cm) y altura total entre 5.3 a 18.4 m (media de 14.2 m) (tabla 3). El incremento medio anual (IMA4 años) presentó un promedio de 18.4 m3.ha-1.año-1, variando entre 0.56 y 46.5 m3.ha-1.año-1 (tabla 3C. Los rasgos con menor variación fueron la altura total (h) y el diámetro (d) con coeficientes de variación de 9.5 y 16.6 %, respectivamente (tabla 3), mientras que los de mayor variabilidad fueron NR (157.4 %), APC (58.4 %) y AIR (42.3 %).
Tabla 3: Media, valor mínimo y máximo, coeficiente de variación (CV%) para sobrevivencia y rasgos de crecimiento y de calidad del árbol para 40 familias de polinización abierta de T. grandis, a los 4.3 años, en el municipio de Armero-Guayabal, Tolima (n = 1070).
Componentes de varianza y heredabilidad estimada
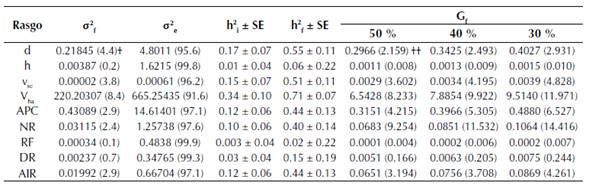
La varianza residual o varianza dentro de las familias (σ2 e) fue la mayor fuente de variación en el ensayo, oscilando entre 91.6 a 99.9 % de la varianza total de los rasgos evaluados (tabla 5). De manera recíproca, los componentes de varianza familiar (σ2 f) fueron bajos, variando entre 0.1 a 8.4 %. El volumen por hectárea (Vha), diámetro (d), volumen del fuste sin corteza (vsc) y el área de proyección de copa (APC) presentaron los componentes de varianza familiar más altos, con 8.4, 4.4, 3.8, 2.9 % del total de la variación fenotípica, respectivamente. Estos rasgos mostraron las mayores heredabilidades individuales con valores de 0.34, 0.174, 0.15 y 0.12, para Vha, d, vsc y APC, respectivamente (tabla 4). Entre los rasgos de calidad del árbol se destacan las variables ángulo de inserción de ramas (AIR) y número de renuevos (NR), los cuales presentaron heredabilidades de 0.10 y 0.12, respectivamente. Los rasgos restantes presentaron heredabilidades bajas a nulas. La heredabilidad basada en la media familiar siguió el mismo comportamiento que la heredabilidad individual, sin embargo, presentó valores mayores (tabla 4).
SE: Error estándar. † Valores en paréntesis son el porcentaje del total de la varianza. †† Valores en paréntesis son el porcentaje de ganancia obtenida sobre la media del respectivo rasgo analizado.
Tabla 4: Componentes de varianza, heredabilidad y ganancia genética estimada con 3 diferentes intensidades de selección para los rasgos de crecimiento y de calidad del árbol para familias de polinización abierta de T. grandis, a los 4.3 años, en el municipio de Armero-Guayabal, Tolima. d: Diámetro a 1.3 m, h: Altura total, vsc: Volumen del fuste sin corteza, Vha: Volumen por hectárea, APC: Área de proyección de copa, NR: Número de renuevos, RF: Rectitud fustal, DR: Diámetro de ramas, AIR: Ángulo de inserción de ramas.
Ganancia genética
Las ganancias genéticas obtenidas para cada rasgo son presentadas en la tabla 4. El diámetro (d) presentó ganancias genéticas modestas menores a 3 % cuando se seleccionan las mejores 12 familias (que corresponde al 30 % de intensidad de selección). La ganancia genética en volumen del fuste sin corteza (vsc) varió entre 3.60 y 4.83 % cuando se seleccionan las 20 familias (50 % intensidad de selección) y 12 familias (30 % de intensidad de selección) superiores, respectivamente. Similares resultados se obtuvieron para el Vha, mostrando valores entre 8.23 y 11.97 % cuando se seleccionan 50 o 30 % de las mejores familias, respectivamente. El APC alcanzó valores de 6.53 % y 4.22 % cuando se seleccionan las mejores 12 familias o 20 familias. La ganancia genética para NR y AIR presentaron valores de 9.25 % y 14.42 % y de 3.19 % y 4.26 % cuando se seleccionan 50 y 30 % de las familias, respectivamente. Por otro lado, la altura total (h) y la RF muestran ganancias genéticas muy bajas cercanas a cero.
Correlaciones genéticas
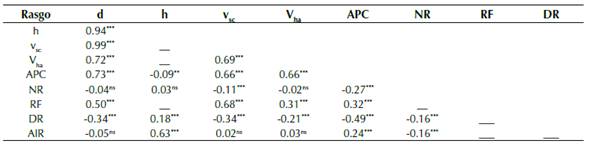
El volumen del fuste sin corteza (vsc) y volumen total por hectárea (Vha) correlacionaron positivamente con el d, APC, y RF y negativamente con DR (tabla 5). Árboles de mayor diámetro, rectos, con copa grande y menor diámetro de ramas producirán mayor volumen fustal y total (a nivel de hectárea). El vsc mostró una correlación negativa baja pero significativa con NR (r = -0.11, p < 0.001), contrariamente Vha no correlacionó con el NR. Adicionalmente, vsc y Vha no correlacionaron con AIR (p > 0.05). Entretanto, no fue posible calcular la correlación entre algunos rasgos, principalmente aquellos que presentaron una baja a nula heredabilidad, como el caso de h, RF y DR (tabla 5).
ns: no significativo; *: p < 0.05; **: p < 0.01; ***: p < 0.001.
Tabla 5: Coeficientes de correlación genética entre rasgos de crecimiento y de calidad del árbol (n = 1070). d: Diámetro a 1.3 m, h: Altura total, vsc: Volumen del fuste sin corteza, Vha: Volumen por hectárea, APC: Área de proyección de copa, NR: Número de renuevos, RF: Rectitud fustal, DR: Diámetro de ramas, AIR: Ángulo de inserción de ramas. La significancia de las correlaciones fue evaluada empleando la prueba de t-student.
Ranking de familias
En la figura 2 se presenta un ranking de las 40 familias considerando la producción de volumen total por hectárea. Entre la familia con mejor (37) y peor (35) desempeño, hay una diferencia de 59.98 m3.ha-1 (121 %). El promedio de las mejores 12 familias (92.88 m3.ha-1) supera en 13.41 m3.ha-1 al promedio general de las 40 familias. De las mejores 12 familias del ranking, un 67 % de ellas son procedentes del municipio de Canalete, Córdoba. Entre las familias evaluadas sobresalen las familias 37, 31 y 20, provenientes de los municipios de Canalete y San Antero, Córdoba, las cuales superaron la media del volumen por hectárea de todas las 40 familias (figura 2).
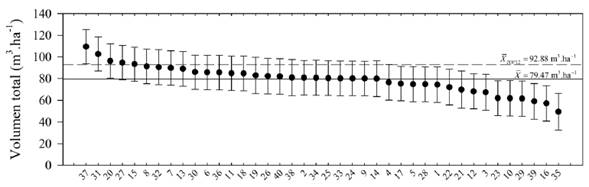
Figura 2: Valores genéticos (puntos) y límites de confianza (barras) para el volumen total por hectárea de 40 familias de polinización abierta de T. grandis a los 4.3 años. Dentro del gráfico se indica el promedio general de las 40 familias (línea horizontal continua) y de las mejores 12 familias (línea horizontal discontinua).
DISCUSIÓN
El estudio presenta una evaluación de la variación genética de rasgos de crecimiento y calidad del árbol entre familias de polinización abierta de T. grandis en un ensayo de progenies de 4.3 años de edad en el norte del Tolima, Colombia. La especie presenta una adecuada adaptación a las condiciones ambientales de los valles interandinos del alto Magdalena, mostrando una alta sobrevivencia (intervalo de 73-100 %) (figura 1). Es sabido que esta zona suple en cierta medida los requerimientos ambientales de la especie, principalmente aquellos relacionados con clima, suelo y altitud (Fonseca, 2003; Callister y Collins, 2008; Chaix et al., 2011; Goh et al., 2013).
La heredabilidad individual (h2 i) fue moderada para la mayoría de los rasgos evaluados (tabla 4). El Vha, d, vsc y APC mostraron las mayores heredabilidades individuales con valores de 0.34, 0.17, 0.15 y 0.12, respectivamente (tabla 4). Similares resultados fueron reportados por Callister y Collins (2008) para un ensayo de progenies de teca de 3.5 años en Tailandia, en el cual se hallaron heredabilidades en sentido estricto de 0.22 para diámetro y 0.18 para volumen. De manera similar, Goh et al. (2013) reportaron heredabilidades en sentido amplio de 0.120, 0.257, y 0.125 para diámetro, altura y volumen, respectivamente, para clones de teca de 7 años en Sabah (Malasia). Es así como ambos estudios son consistentes con los resultados presentados aquí.
Sin embargo, Sharma et al. (2000) reportaron heredabilidades en sentido estricto mayores o iguales a 0.64, 0.77 y 0.69 para diámetro, altura total y área basal para familias de polinización abierta de teca de 7 años en Orissa (India). Estudios previos por Cornelius (1994), Callister y Collins (2008) y Narayanan et al. (2009) han reportado valores de heradabilidad entre 0.18 a 0.34 para la altura total; no obstante, el presente estudio encontró valores de heradabilidad nulos (h2 i = 0.01) para este rasgo. Este resultado se puede asociar al nivel de competencia existente al interior del rodal, fenómeno que ocurre mayormente por la radiación solar, lo que impulsa a los árboles a ocupar los estratos altos del dosel y aumentar su uniformidad (Harris, 2007). Entre los rasgos de calidad del árbol evaluados sobresalen el AIR y NR los cuales presentaron heredabilidades de 0.10 y 0.12, respectivamente. De manera contraria, RF y DR presentaron heredabilidades bajas a nulas (tabla 4). Callister y Collins (2008) reportaron heredabilidades bajas para rectitud fustal (h2 i = 0.07) y brotes epicórmicos (h2 i = 0.03). Semejante a los estudios de Narayanan et al. (2009) y Goh et al. (2013), en el presente estudio las heredabilidades familiares fueron mayores que las heredabilidades individuales, tal y como se espera, ya que toma en cuenta el aporte en la variación genética del colectivo de individuos que componen la familia.
La ganancia genética esperada para el volumen del fuste sin corteza y volumen por hectárea, considerando una intensidad de selección de 30 %, fue de 4.83 % y 11.97 % respectivamente (tabla 4), menores a las reportadas por Espitia et al. (2011), con valores de 12.5 % en la selección de árboles plus de teca en el departamento de Córdoba (Colombia). Estas discrepancias podrían explicarse debido a las diferencias en los valores de heredabilidad empleados. En el estudio mencionado, se emplearon valores estimados a partir de estudios previos. Fleitas-Camacho et al. (2010) reportaron ganancias genéticas entre 17-19 % para altura total y de 25 % para el diámetro, seleccionando el 30 % de las mejores familias de teca de 3.5 años de edad en Cuba. Contrariamente, en el presente estudio estos rasgos obtuvieron ganancias bajas menores a 2.93 % para diámetro y 0.010 % para altura. El NR presentó la mayor ganancia genética esperada alcanzando hasta un 14.42 % para un índice de selección de 30 % (tabla 4). El NR es un rasgo importante en el manejo de plantaciones forestales, y es considerado un elemento crítico en el proceso de conducción de rodales, elevando los costos de mantenimiento, y contribuyendo a la desuniformidad del rodal (Barros et al., 2017). Adicionalmente, la presencia de renuevos está asociada a disminuciones en las tasas de crecimiento cuando la especie es cultivada comercialmente (Fonseca, 2003). El presente estudio soporta esta hipótesis al reportar una correlación negativa entre NR y vsc (rg(xy) = -0.11, p <0.001) y entre NR y APC (rg(xy) = -0.27, p < 0.001) (tabla 5).
Como se esperaba, el análisis de correlación mostró que árboles de mayor diámetro presentan mayor volumen. Estudios previos han reportado escasa correlación entre rasgos de crecimiento y de calidad del árbol (Chaix et al., 2011). Contrariamente, en el presente estudio el análisis de correlación mostró que árboles más rectos con copa grande producirán mayor volumen (tabla 5). Callister y Collins (2008) también reportaron una correlación positiva entre la rectitud fustal y el volumen (rg(xy) = 0.40), para progenies de teca de 3.5 años de edad. Por otro lado, el vsc y Vha mostraron correlaciones significativas negativas moderadas con DR (tabla 5).
El ranking genético construido a partir de los BLUP respecto a la media de todas las 40 familias permitió seleccionar a partir de la población base un conjunto de familias con características deseables en cuanto a la producción de volumen por hectárea. El cálculo del volumen total por hectárea integró los caracteres de crecimiento de cada progenie (diámetro y altura total) y la sobrevivencia de cada familia, por lo tanto, es un buen indicador del valor económico real de cada familia (Pavlotzky y Murillo, 2013). La selección de las mejores 12 familias permitiría obtener una ganancia esperada de 13.41 m3.ha-1, lo que equivale a 11.97 % de avance (figura 2). El incremento medio anual (IMA4 años) presentó un promedio de 18.37 m3.ha-1.año-1 para las 40 familias. Este valor esta dentro del rango presentado por Vallejo y Avendaño (2013) para plantaciones de teca bajo condiciones favorables de crecimiento inicial con IMAs entre 10 a 20 m3.ha-1.año-1. Sin embargo, en el presente estudio, 9 familias superaron el intervalo presentado por Vallejo y Avendaño (2013) con un IMA4 años máximo de 25.2 m3.ha-1.año-1 alcanzado por la familia 37 procedente de Canalete (Córdoba). Finalmente, los resultados muestran la existencia de familias de teca con características superiores en términos de sus tasas de crecimiento, calidad y adaptación a las condiciones ambientales del alto Magdalena que permitirán ampliar la oferta de material propagativo y así contribuir a la expansión de la base de plantaciones en el interior del país.
CONCLUSIONES
Las familias de polinización abierta de T. grandis evaluadas presentaron una adecuada adaptación al sitio, reflejándose en una sobrevivencia promedio de 89 % a los 4.3 años. La heredabilidad, en sentido estricto con respecto a la media individual, presentó valores moderados para Vha (0.34), d (0.17), vsc (0.15), APC (0.12), AIR (0.10) y NR (0.12). Por otro lado, los valores de heredabilidad media familiar fueron de 0.71 para Vha, 0.55 para d, 0.51 para vsc, 0.44 para APC, 0.44 para AIR y 0.40 para NR. Estos valores muestran que estos rasgos son moderadamente transferibles de los progenitores a sus progenies.
Asimismo, se registró una correlación positiva entre el volumen del fuste sin corteza y el volumen por hectárea con la rectitud del fuste, mientras que la correlación fue negativa con el diámetro de ramas.
Se estimó una ganancia genética de 4.83 % para el volumen del fuste sin corteza, 11.97 % para el volumen por hectárea, 14.42 % para el número de renuevos y 4.26 % para el ángulo de inserción de ramas al seleccionar las mejores 12 familias del ranquin genético (30 % de intensidad de selección). Estos resultados representan avances significativos en la mejora genética de la especie.
Entre las familias de polinización abierta evaluadas sobresalen las familias 37, 31 y 20, provenientes de los municipios de Canalete y San Antero, Córdoba, las cuales superaron la media del volumen por hectárea de todas las 40 familias. Estos genotipos (y sus progenies) deberán ser considerados para el establecimiento de nuevas plantaciones comerciales y en estudios complementarios para la mejora genética de la especie. Finalmente, se recomienda evaluar otros caracteres de interés comercial, como las propiedades físicas y mecánicas de la madera y su relación con las tasas de crecimiento, que permitan identificar familias e individuos con mayor valor comercial. Así mismo, se recomienda incluir en futuros análisis la localización espacial de cada individuo dentro del ensayo para modelar el efecto de la autocorrelación espacial y aislar los posibles efectos del micrositio.
Acknowledgements
AGRADECIMIENTOS
Los autores agradecen a la Oficina de Investigaciones y Desarrollo Científico de la Universidad del Tolima por el apoyo financiero otorgado para desarrollar la presente investigación a través de los proyectos n.º 340130516, 180130516 y 620120516 ejecutados por el Grupo de Investigación Madera y Bosques de la Universidad del Tolima.
REFERENCIAS




Licencia
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).






